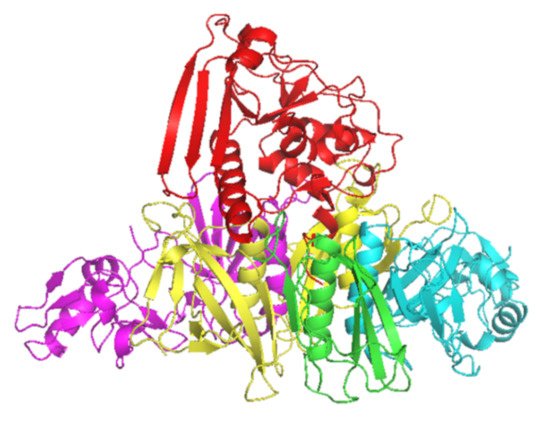
Pertussis toxin (PT) is considered the main virulence factor causing whooping cough or pertussis. The protein is widely studied and its composition was revealed and sequenced already during the 1980s. The human immune system creates a good response against PT when measured in quantity. However, the serum anti-PT antibodies wane rapidly, and only a small amount of these antibodies are found a few years after vaccination/infection. Therefore, multiple approaches to study the functionality (quality) of these antibodies, e.g., avidity, neutralizing capacity, and epitope specificity, have been investigated. In addition, the long-term B cell memory (Bmem) to PT is crucial for good protection throughout life.
- antibodies
- avidity
- B cells
- CHO cell
- epitopes
- pertussis toxin
1. Introduction
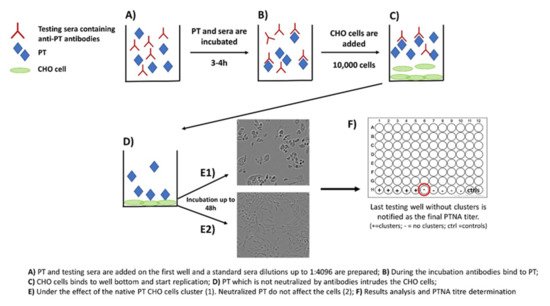
2. The Role of Neutralizing Antibodies to Pertussis Toxin (PTNAs)
This entry is adapted from the peer-reviewed paper 10.3390/toxins13080508
References
- Pittman, M. Pertussis toxin: The cause of the harmful effects and prolonged immunity of whooping cough. A hypothesis. Rev. Infect. Dis. 1979, 1, 401–412.
- Locht, C. Molecular aspects of Bordetella pertussis pathogenesis. Int. Microbiol. 1999, 2, 137–144.
- Carbonetti, N.H. Pertussis toxin and adenylate cyclase toxin: Key virulence factors of Bordetella pertussis and cell biology tools. Future Microbiol. 2010, 5, 455–469.
- Nicosia, A.; Perugini, M.; Franzini, C.; Casagli, M.C.; Borri, M.G.; Antoni, G.; Neri, P.; Ratti, G.; Rappuoli, R. Cloning and sequencing of the pertussis toxin genes: Operon structure and gene duplication. Proc. Natl. Acad. Sci. USA 1986, 83, 4631–4635.
- Sato, H.; Ito, A.; Chiba, J.; Sato, Y. Monoclonal antibody against pertussis toxin: Effect on toxin activity and pertussis infections. Infect. Immun. 1984, 46, 422–428.
- Locht, C.; Keith, J.M. Pertussis toxin gene: Nucleotide sequence and genetic organization. Science 1986, 232, 1258–1264.
- Podda, A.; Nencioni, L.; De Magistris, M.T.; Di Tommaso, A.; Bossù, P.; Nuti, S.; Pileri, P.; Peppoloni, S.; Bugnoli, M.; Ruggiero, P.; et al. Metabolic, humoral, and cellular responses in adult volunteers immunized with the genetically inactivated pertussis toxin mutant PT-9K/129G. J. Exp. Med. 1990, 172, 861–868.
- Podda, A.; Carapella De Luca, E.; Titone, L.; Casadei, A.M.; Cascio, A.; Bartalini, M.; Volpini, G.; Peppoloni, S.; Marsili, I.; Nencioni, L.; et al. Immunogenicity of an acellular pertussis vaccine composed of genetically inactivated pertussis toxin combined with filamentous hemagglutinin and pertactin in infants and children. J. Pediatr. 1993, 123, 81–84.
- Rappuoli, R. The vaccine containing recombinant pertussis toxin induces early and long-lasting protection. Biologicals 1999, 27, 99–102.
- Cherry, J.D.; Gornbein, J.; Heininger, U.; Stehr, K. A search for serologic correlates of immunity to Bordetella pertussis cough illnesses. Vaccine 1998, 16, 1901–1906.
- Storsaeter, J.; Hallander, H.O.; Gustafsson, L.; Olin, P. Levels of anti-pertussis antibodies related to protection after household exposure to Bordetella pertussis. Vaccine 1998, 16, 1907–1916.
- Storsaeter, J.; Hallander, H.O.; Gustafsson, L.; Olin, P. Low levels of antipertussis antibodies plus lack of history of pertussis correlate with susceptibility after household exposure to Bordetella pertussis. Vaccine 2003, 21, 3542–3549.
- Schure, R.M.; Hendrikx, L.H.; de Rond, L.G.; Oztürk, K.; Sanders, E.A.; Berbers, G.A.; Buisman, A.M. Differential T- and B-cell responses to pertussis in acellular vaccine-primed versus whole-cell vaccine-primed children 2 years after preschool acellular booster vaccination. Clin. Vaccine Immunol. 2013, 20, 1388–1395.
- Carollo, M.; Pandolfi, E.; Tozzi, A.E.; Buisman, A.M.; Mascart, F.; Ausiello, C.M. Humoral and B-cell memory responses in children five years after pertussis acellular vaccine priming. Vaccine 2014, 3, 2093–2099.
- Plotkin, S.A. Complex correlates of protection after vaccination. Clin. Infect. Dis. 2013, 56, 1458–1465.
- Edelman, K.; He, Q.; Makinen, J.; Sahlberg, A.; Haanpera, M.; Schuerman, L.; Wolter, J.; Mertsola, J. Immunity to pertussis 5 years after booster immunization during adolescence. Clin. Infect. Dis. 2007, 44, 1271–1277.
- Dalby, T.; Petersen, J.W.; Harboe, Z.B.; Krogfelt, K.A. Antibody responses to pertussis toxin display different kinetics after clinical Bordetella pertussis infection than after vaccination with an acellular pertussis vaccine. J. Med. Microbiol. 2010, 59, 1029–1036.
- Hallander, H.O.; Ljungman, M.; Storsaeter, J.; Gustafsson, L. Kinetics and sensitivity of ELISA IgG pertussis antitoxin after infection and vaccination with Bordetella pertussis in young children. APMIS 2009, 117, 797–807.
- Dalby, T.; Andersen, P.H.; Hoffmann, S. Epidemiology of pertussis in Denmark, 1995 to 2013. Eurosurveillance 2016, 21.
- Bhavsar, A.; Mertsola, J.; Poulsen, A.; Silfverdal, S.A. Pertussis in infants in Nordic countries. Acta Paediatr. 2021.
- Barkoff, A.M.; Grondahl-Yli-Hannuksela, K.; He, Q. Seroprevalence studies of pertussis: What have we learned from different immunized populations. Pathog. Dis. 2015, 73.
- Minh, N.N.; He, Q.; Ramalho, A.; Kaufhold, A.; Viljanen, M.K.; Arvilommi, H.; Mertsola, J. Acellular vaccines containing reduced quantities of pertussis antigens as a booster in adolescents. Pediatrics 1999, 104, e70.
- Blanchard Rohner, G.; Chatzis, O.; Chinwangso, P.; Rohr, M.; Grillet, S.; Salomon, C.; Lemaître, B.; Boonrak, P.; Lawpoolsri, S.; Clutterbuck, E.; et al. Boosting Teenagers with Acellular Pertussis Vaccines Containing Recombinant or Chemically Inactivated Pertussis Toxin: A Randomized Clinical Trial. Clin. Infect. Dis. 2019, 68, 1213–1222.
- Locht, C.; Mielcarek, N. New pertussis vaccination approaches: En route to protect newborns? FEMS Immunol. Med. Microbiol. 2012, 66, 121–133.
- Van der Lee, S.; van Rooijen, D.M.; de Zeeuw-Brouwer, M.L.; Bogaard, M.J.M.; van Gageldonk, P.G.M.; Marinovic, A.B.; Sanders, E.A.M.; Berbers, G.A.M.; Buisman, A.-M. Robust Humoral and Cellular Immune Responses to Pertussis in Adults after a First Acellular Booster Vaccination. Front. Immunol. 2018, 9, 681–690.
- Hovingh, E.S.; Kuipers, B.; Bonacic Marinovic, A.A.; Jan Hamstra, H.; Hijdra, D.; Mughini Gras, L.; Van Twillert, I.; Jongerius, I.; Van Els, C.A.C.M.; Pinelli, E. Detection of opsonizing antibodies directed against a recently circulating Bordetella pertussis strain in paired plasma samples from symptomatic and recovered pertussis patients. Sci. Rep. 2018, 8, 12039.
- Hewlett, E.L.; Sauer, K.T.; Myers, G.A.; Cowell, J.L.; Guerrant, R.L. Induction of a novel morphological response in Chinese hamster ovary cells by pertussis toxin. Infect. Immun. 1983, 40, 1198–1203.
- Gillenius, P.; Jäätmaa, E.; Askelöf, P.; Granström, M.; Tiru, M. The standardization of an assay for pertussis toxin and antitoxin in microplate culture of Chinese hamster ovary cells. J. Biol Stand. 1985, 13, 61–66.
- Zhang, Y.; Li, Y.; Chen, Z.; Liu, X.; Peng, X.; He, Q. Determination of serum neutralizing antibodies reveals important difference in quality of antibodies against pertussis toxin in children after infection. Vaccine 2021, 39, 1826–1830.
- Barkoff, A.M.; Grondahl-Yli-Hannuksela, K.; Vuononvirta, J.; Mertsola, J.; Kallonen, T.; He, Q. Differences in avidity of IgG antibodies to pertussis toxin after acellular pertussis booster vaccination and natural infection. Vaccine 2012, 30, 6897–6902.
- Knuutila, A.; Dalby, T.; Barkoff, A.M.; Jorgensen, C.S.; Fuursted, K.; Mertsola, J.; Markey, K.; He, Q. Differences in epitope-specific antibodies to pertussis toxin after infection and acellular vaccinations. Clin. Transl. Immunol. 2020, 9, e1161.
- Van Twillert, I.; Han, W.G.; van Els, C.A. Waning and aging of cellular immunity to Bordetella pertussis. Pathog. Dis. 2015, 73, ftv071.
- Siegrist, C.A.; Aspinall, R. B-cell responses to vaccination at the extremes of age. Nat. Rev. Immunol. 2009, 9, 185–194.
- Edara, V.V.; Hudson, W.H.; Xie, X.; Ahmed, R.; Suthar, M.S. Neutralizing Antibodies Against SARS-CoV-2 Variants After Infection and Vaccination. JAMA 2021, 325, 1896–1898.
- To, K.K.; Zhang, A.J.; Hung, I.F.; Xu, T.; Ip, W.C.; Wong, R.T.; Ng, J.C.K.; Chan, J.F.W.; Chan, K.-H.; Yuen, K.-Y. High titer and avidity of nonneutralizing antibodies against influenza vaccine antigen are associated with severe influenza. Clin. Vaccine Immunol. 2012, 19, 1012–1018.
- Jalkanen, P.; Pasternack, A.; Maljanen, S.; Melén, K.; Kolehmainen, P.; Huttunen, M.; Lundberg, R.; Tripathi, L.; Kna, H.; Ritvos, M.A.; et al. A combination of N and S antigens with IgA and IgG measurement strengthens the accuracy of SARS-CoV-2 serodiagnostics. J. Infect. Dis. 2021.
- Palmer, S.R.; Balfour, A.H.; Jephcott, A.E. Immunisation of adults during an outbreak of diphtheria. Br. Med. J. Clin. Res. Ed. 1983, 286, 624–626.
- Lagergard, T.; Trollfors, B.; Claesson, B.A.; Karlberg, J.; Taranger, J. Determination of neutralizing antibodies and specific immunoglobulin isotype levels in infants after vaccination against diphtheria. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 341–345.
- Storsaeter, J.; Olin, P. Relative efficacy of two acellular pertussis vaccines during three years of passive surveillance. Vaccine 1992, 10, 142–144.
- Granström, M.; Granström, G. Serological correlates in whooping cough. Vaccine 1993, 11, 445–448.
- Markey, K.; Asokanathan, C.; Feavers, I. Assays for Determining Pertussis Toxin Activity in Acellular Pertussis Vaccines. Toxins 2019, 11, 417.
- Isbrucker, R.; Daas, A.; Wagner, L.; Costanzo, A. Transferability study of CHO cell clustering assays for monitoring of pertussis toxin activity in acellular pertussis vaccines. Pharmeur. Bio Sci. Notes 2016, 2015, 97–114.
- Markey, K.; Douglas-Bardsley, A.; Hockley, J.; Le Tallec, D.; Costanzo, A. Calibration of pertussis toxin BRP batch 1 in a standardised CHO cell-based clustering assay. Pharmeur. Bio Sci. Notes 2018, 2018, 112–123.
- Sato, H.; Sato, Y.; Ito, A.; Ohishi, I. Effect of monoclonal antibody to pertussis toxin on toxin activity. Infect. Immun. 1987, 55, 909–915.
- Granstrom, M.; Granstrom, G.; Gillenius, P.; Askelof, P. Neutralizing antibodies to pertussis toxin in whooping cough. J. Infect. Dis. 1985, 151, 646–649.
- Trollfors, B.; Krantz, I.; Sigurs, N.; Taranger, J.; Zackrisson, G.; Roberson, R. Toxin-neutralizing antibodies in patients with pertussis, as determined by an assay using Chinese hamster ovary cells. J. Infect. Dis. 1988, 158, 991–995.
- Carlsson, R.M.; Claesson, B.A.; Selstam, U.; Fagerlund, E.; Granstrom, M.; Blondeau, C.; Hoffenbach, A. Safety and immunogenicity of a combined diphtheria-tetanus-acellular pertussis-inactivated polio vaccine-Haemophilus influenzae type b vaccine administered at 2–4-6–13 or 3–5-12 months of age. Pediatr. Infect. Dis. J. 1998, 17, 1026–1033.
- Carlsson, R.M.; Claesson, B.A.; Fagerlund, E.; Knutsson, N.; Lundin, C. Antibody persistence in five-year-old children who received a pentavalent combination vaccine in infancy. Pediatr. Infect. Dis. J. 2002, 21, 535–541.
- Sirivichayakul, C.; Chanthavanich, P.; Limkittikul, K.; Siegrist, C.A.; Wijagkanalan, W.; Chinwangso, P.; Petre, J.; Thai, P.H.; Chauhan, M.; Viviani, S. Safety and immunogenicity of a combined Tetanus, Diphtheria, recombinant acellular Pertussis vaccine (TdaP) in healthy Thai adults. Hum. Vaccin. Immunother. 2017, 13, 136–143.
- Pichichero, M.E.; Francis, A.B.; Marsocci, S.M.; Green, J.L.; Disney, F.A.; Meschievitz, C. Safety and immunogenicity of an acellular pertussis vaccine booster in 15- to 20-month-old children previously immunized with acellular or whole-cell pertussis vaccine as infants. Pediatrics 1993, 91, 756–760.
- Pichichero, M.E.; Deloria, M.A.; Rennels, M.B.; Anderson, E.L.; Edwards, K.M.; Decker, M.D.; Englund, J.A.; Steinhoff, M.C.; Deforest, A.; Meade, B.D. A safety and immunogenicity comparison of 12 acellular pertussis vaccines and one whole-cell pertussis vaccine given as a fourth dose in 15- to 20-month-old children. Pediatrics 1997, 100, 772–788.
- Giuliano, M.; Mastrantonio, P.; Giammanco, A.; Piscitelli, A.; Salmaso, S.; Wassilak, S.G. Antibody responses and persistence in the two years after immunization with two acellular vaccines and one whole-cell vaccine against pertussis. J. Pediatr. 1998, 132, 983–988.
- Rutter, D.A.; Ashworth, L.A.; Day, A.; Funnell, S.; Lovell, F.; Robinson, A. Trial of a new acellular pertussis vaccine in healthy adult volunteers. Vaccine 1988, 6, 29–32.
- Greco, D.; Salmaso, S.; Mastrantonio, P.; Giuliano, M.; Tozzi, A.E.; Anemona, A.; Ciofi degli Atti, M.L.; Giammanco, A.; Panei, P.; Blackwelder, W.C.; et al. A controlled trial of two acellular vaccines and one whole-cell vaccine against pertussis. Progetto Pertosse Working Group. N. Engl. J. Med. 1996, 334, 341–348.
- Meade, B.D.; Lynn, F.; Reed, G.F.; Mink, C.M.; Romani, T.A.; Deforest, A.; Deloria, M.A. Relationships between functional assays and enzyme immunoassays as measurements of responses to acellular and whole-cell pertussis vaccines. Pediatrics 1995, 96, 595–600.
- Millen, S.H.; Bernstein, D.I.; Connelly, B.; Ward, J.I.; Chang, S.J.; Weiss, A.A. Antibody-mediated neutralization of pertussis toxin-induced mitogenicity of human peripheral blood mononuclear cells. Infect. Immun. 2004, 72, 615–620.
- Isacson, J.; Trollfors, B.; Lagergrd, T.; Taranger, J. Comparison of a toxin neutralization assay and ELISA for determination of pertussis toxin antibodies. Serodiagn. Immunother. Infect. Dis. 1997, 8, 163–167.
- Sato, Y.; Sato, H.; Kodama, H.; Uchimura, M.; Miwa, N.; Kobayashi, T.; Yamamoto, E.; Fujita, I.; Kumamoto, T. An improved ELISA system for the measurement of IgG antibodies against pertussis toxin (PT) and filamentous hemagglutinin (FHA) in human sera. Dev. Biol. Stand. 1991, 73, 167–174.
- Dalby, T.; Sørensen, C.; Petersen, J.W.; Krogfelt, K.A. Pertussis serology: Assessment of IgG anti-PT ELISA for replacement of the CHO cell assay. APMIS 2010, 118, 968–972.
- Moriuchi, T.; Otsuka, N.; Hiramatsu, Y.; Shibayama, K.; Kamachi, K. A high seroprevalence of antibodies to pertussis toxin among Japanese adults: Qualitative and quantitative analyses. PLoS ONE 2017, 12, e0181181.
- Weiss, A.A.; Mobberley, P.S.; Fernandez, R.C.; Mink, C.M. Characterization of human bactericidal antibodies to Bordetella pertussis. Infect. Immun. 1999, 67, 1424–1431.
- Aase, A.; Herstad, T.K.; Merino, S.; Brandsdal, K.T.; Berdal, B.P.; Aleksandersen, E.M.; Aaberge, I.S. Opsonophagocytic activity and other serological indications of Bordetella pertussis infection in military recruits in Norway. Clin. Vaccine Immunol. 2007, 14, 855–862.