Pseudophosphatases are important regulators, and their proper regulation is important for maintaining homeostasis and preventing diseases.
- pseudoenzymes
- pseudophosphatases
- protein tyrosine phosphatases (PTPs)
- dual specificity phosphatases (DUSPs)
- myotubularin phosphatases (MTMs)
- STYX (phosphoserine/threonine/tyrosine-interacting protein)
- MK-STYX (MAPK (mitogen-activated protein kinase) phosp
1. Introduction
Over the past decade, the relevance of pseudoenzymes has been elevated and solidified as important signaling regulators. These modes of action and their roles in various cellular processes serve as an idea link for the roles of pseudoenzymes in pathologies and diseases. Misregulation of pseudoenzymes has been implicated in the etiology of various diseases such as cancer, obesity, and neurological disorders [4,5,7,8,9]. This review highlights the signaling pseudophosphatases and their implications in diseases, while demonstrating some of their essential roles in preventing or causing such diseases.
Pseudophosphatases of PTPs are widely accepted as having mutations within their signature active site motif (HCX5R) They maintain the three-dimensional fold and the ability to bind phosphorylated proteins [4,11,16]. Recent reports have advanced the field, complicating the mutated sequence definition [11,17] and the “grab and hold on” perspective, where pseudophosphatases stably interact with phosphorylated residues, thereby competitively inhibiting active phosphatases or kinases [11,16]. For example, the histidine domain containing protein tyrosine phosphatase (HD-PTP) is reported as a pseudophosphatase [4,5,18,19,20,21,22] and an active phosphatase [4,5,14,23,24].
This dual and contradictory functionality of HD-PTP highlights the complexities of investigating pseudophosphatases. Wishart and Dixon defined pseudophosphatases as STYX domains, and demonstrated a loss of function of dephosphorylating phosphorylated substrates [12]; thus, they defined the prototypical STYX domain beyond pure bioinformatics [12]. The analysis of pseudophosphatases through a combination of sequencing and functional experiments—dephosphorylation assays with appropriate substrates and contextual situations (localization, substrates/interactors, or abundance of interactor or pseudphosphatase—will provide more of an insight into the full spectrum [4,5,7]. Nevertheless, the roles of pseudophosphatases as regulators in many cellular processes such as spermatogenesis, stress response, neuronal differentiation, cell fate, migration, ubiquitylation, demyelination, oocyte-to-zygote transition, transcription, and apoptosis [4,6,7,16,20,22,25,26] have led to interest into their apparent roles in diseases.
The increased interest in pseudophosphatases resulted in the discovery of their relevance in many diseases. provided a useful framework for pseudophosphatases’ linkage to diseases [7]. Mutations of these molecules or their misregulation leads to diseases such as leukemia, breast cancer, colorectal cancer, hepatocarcinoma, glioblastoma, other cancers, Charcot-Marie-Tooth (CMT) disease (abnormal nerve myelination), obesity, diabetes, chronic obstructive pulmonary disease (COPD), and nephrotic syndrome [5,7,27,28,29,30,31,32,33]. A comprehensive list of the known functions of pseudophosphatases and their implications in diseases is provided in Table 1.
| Pseudophosphatase Name | Alternative Names | Normal Function | Role in Disease |
|---|---|---|---|
| MTMR5 (Myotubularin-related protein 5) |
SBF1 |
|
|
| MTMR9 (Myotubularin-related protein 9) |
|||
| MTMR10 (Myotubularin-related protein 10) |
- |
|
|
| MTMR11 (Myotubularin-related protein 11) |
- | ||
| MTMR12 (Myotubularin-related protein 12) |
3-PAP | - | |
| MTMR13 (Myotubularin-related protein 13) |
SBF2 |
|
|
| STYX (Serine/threonine/tyrosine -interacting protein) |
|||
| MK-STYX (Mitogen-activated protein kinase phosphoserine/ threonine/tyrosine-binding protein) |
STYXL1, DUSP-24 |
|
|
| STYXL2 (Serine/threonine/tyrosine-interacting-like protein 2) |
DUSP-27 (duplicated) |
|
|
| TAB1 (TGF-beta-activated kinase 1 and MAP3K7-binding protein 1) |
MAP3K7IP1 | ||
| Tensin 1 | TNS1 |
The proteins here all contain alterations in their catalytic active site motifs that are associated with loss of phosphatase function.
Furthermore, those that have not been linked to diseases have functions that may allude to a possible role in diseases. The diversity of these diseases, such as various types of cancer, COPD, CMT, obesity, diabetes, etc. Furthermore, their diverse range, while validating the intricate roles of pseudophosphatases in signaling cascades, also makes understanding their many functions in diseases more complicated. Considering this special edition focused on the roles of protein tyrosine phosphatases in signaling, with an emphasis on therapeutic strategies, the pseudophosphatases’ signaling mechanisms linked to diseases, myotubularins, tensins, and STYX pseudophosphatases, was discussed in more detail.
2. Myotubularin in Diseases
Six of these genes produce pseudophosphatases, which form complexes with their active homologs [27,71,72] (Figure 1A). This dimerization is context dependent, resulting in complex stability, enhancement of catalytic function (increased phosphatase activity), and regulation of the subcellular localization of the active phosphatase [73,74]. The coupling of active and inactive MTMs to form heterodimers is common; however, self-association to form homodimers also occurs among both active and inactive MTMs (Figure 1A) It is of interest that some MTMs have not been reported to interact directly as dimers [72].
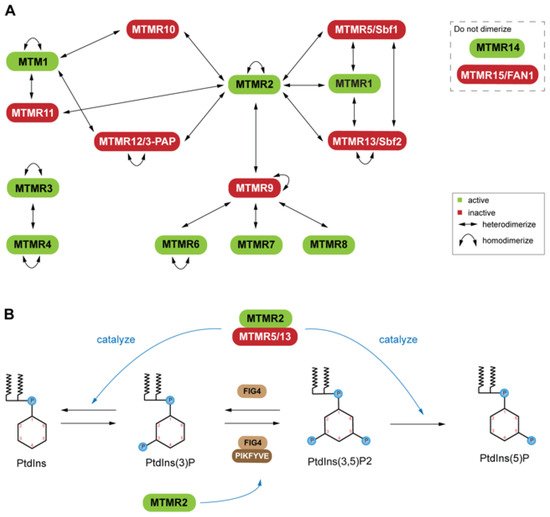
For example, mutations of MTMR2 and MTMR13 leads to misregulation of the AKT signaling in Type 4B Charcot-Marie-Tooth (CMT4B) disease [76]. Overexpression of MTMR2 prevents sustained activation of epidermal growth factor, which leads to sustained AKT activation in transgenic mice [76]. PtdIns(3)P and PtdIns(3,5)P2are important molecules for endosomal-lysosomal membrane trafficking [70], which is tightly regulated by PtdIns kinase and phosphatase activity. MTMs have also been linked to X-linked centronuclear myopathy (XLCNM)
(phosphatase) forms heterodimers with MTMR13 (pseudophosphatase) or MTMR5 (pseudophosphatase), and heterodimers with MTMR13 (pseudophosphatase) or MTMR5 (pseudophosphatase), and Genetic studies in mice revealed that MTMR12 interacts with another phosphatase, polyphosphoinositde phosphatase (FIG4) MTM pseudophosphatase: phosphatase heterodimer signaling complexes are required for cellular processes such as differentiation, membrane trafficking, endocytosis, and survival [10,27,69,70].
3. STYX Pseudophosphatases in Disease
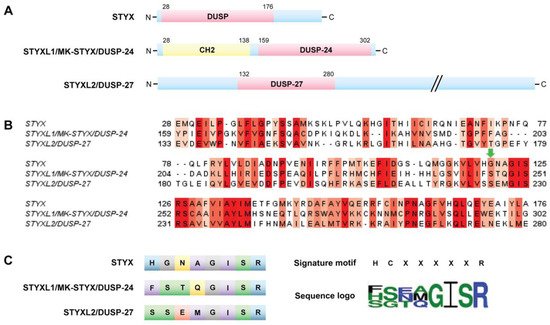
The term “STYX” (phosphoserine/threonine/tyrosine-interacting protein) was coined to designate the phosphotyrosine binding domain that has no catalytic activity, or “dead” phosphatases or pseudophosphatase [12]. These “dead” or STYX domain phosphatases allude to the Greek mythological STYX river of the dead [11,16]. Usage of a point mutation to “restore” catalytic activity in the STYX domain has proved to be a helpful tool to initiate the process of investigating molecules that contain STYX domains. STYX domain phosphatases are pseudophosphatases; therefore, the authors have referred to them as STYX pseudophosphatases, as previously reported [11].
They are all members of the dual specificity protein tyrosine phosphatase (DUSP) family; however, they belong to different subfamilies [5]. These domains vary both from the active site signature motif and each other (Figure 5B,C). Of note, all these STYX domain proteins have glycine, isoleucine, serine, and arginine in the active site motif (Figure 5B,C). Furthermore, early reports classified glycine as a conserved residue of the active site motif, HCxxGxxR [10,24].
STYX and MK-STYX have been shown to be important signaling regulators in cellular processes such as cell cycle, spermatogenesis, cell-fate decisions, cell migration, ubiquitylation and protein degradation, apoptosis, and neuronal differentiation [25,26,51,52,54,100,101], alluding to their roles in the etiology of various diseases. Furthermore, these STYX pseudophosphatases have roles in cancer such as colorectal, Ewing sarcoma, and hepatocarcinoma [8,28,33]. In addition, STYX promotes oncogenesis in colorectal cancer by inhibiting FBXW7, blocking the degradation of cyclin E and c-Jun Therefore, free (unbound) cyclin E and c-Jun are able to promote proliferation in colorectal cancer.
MK-STYX lacks the critical cysteine in the active site signature motif (HCX5R) An increase in MK-STYX expression also promotes hepatocellular carcinoma (Figure 6(Bii)) mRNA expression results in upregulation of PI3K (phosphatidyl 3-kinase)/AKT pathway proteins and an enhancement of proliferation, while reducing apoptosis in hepatocellular carcinoma STYXL2 is a downstream target of the Janus activated kinase/Signal transducers and activators of transcription signaling pathway [99], suggesting that STYXL2 has important roles in signaling cascades.
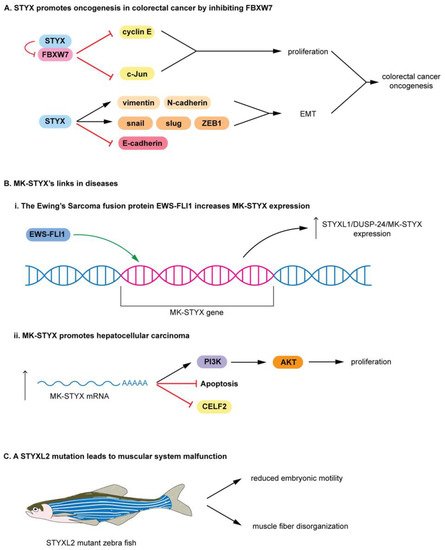
In addition, a transgene integration intoSTYXL2reduces STYXL2 expression in zebrafish, resulting in muscular system malfunction (Figure 6C) [5,99]. STYXL2 mutants have reduced embryonic motility (low paralysis), displaying low spontaneous coiling movements and severely reduced touch response (Figure 6C) [99]. Furthermore, STYXL2 mutants have a major disruptions in the contractile apparatus of their muscle fibers–disorganized muscle fiber formation (Figure 6C) [99].
4. Atypical Pseudophosphatases
This review focused on the less contradictory pseudophosphatases’ signaling in diseases, with DUSP27/STYXL2 as the exception; Reiterer et al. encompasses such contradictory pseudophosphatases [5]. In addition, the D2 domains, which are catalytically inactive and recognize substrate [10], of the PTP receptor molecules were excluded. Recently, the protein tyrosine phosphatase receptor U (PTPRU) was classified as pseudophosphatase [111], both its DUSP domains, D1 and D2, were considered as pseudophosphatase domains [111]. In addition, tensin 2 is thought to have possible activity by dephosphorylating insulin receptor substrate-1 [116].
Some pseudophosphatases were reported as having catalytic activity [4,5,7]. Phosphatase of regenerating liver-3 (PRL3) phosphatase is an example of the concept of a pseudo-pseudophosphatase, which recognizes the catalytic activity and the importance of the noncatalysis in signaling cascades [5,117,118]. In studies with PRL3 mutants specifically defective in either binding the substrate of PRL3 substrate, CBS domain divalent metal cations transport mediators (CNNM) or phosphatase activity [118] demonstrated that the phosphatase activity is dispensable [118]. Moreover, phosphatase activity prevents PRL3-CNNM interaction, which is necessary and sufficient for tumor metastasis [118].
This entry is adapted from the peer-reviewed paper 10.3390/ijms22136924