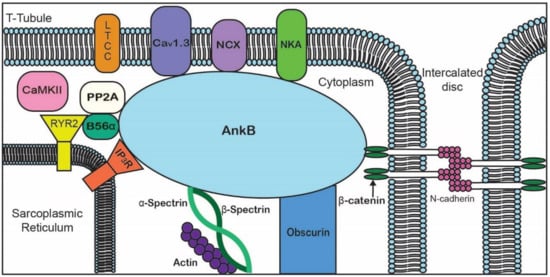
Ankyrin-B (encoded by ANK2), originally identified as a key cytoskeletal-associated protein in the brain, is highly expressed in the heart and plays critical roles in cardiac physiology and cell biology. In the heart, ankyrin-B directs the targeting and localization of key ion channels and transporters, structural proteins, and signaling molecules. The role of ankyrin-B in normal cardiac function is illustrated in animal models lacking ankyrin-B expression, which display significant electrical and structural phenotypes,1 and life-threatening arrhythmias. Further, ankyrin-B dysfunction has been associated with cardiac phenotypes in humans (now referred to as “ankyrin-B syndrome”) including sinus node dysfunction, heart rate variability, atrial fibrillation, conduction block, arrhythmogenic cardiomyopathy, structural remodeling, and sudden cardiac death.
- ankyrin-B
- ANK2
- ion channels
- cardiovascular disease
1. Introduction: Ankyrin Proteins
| Ankyrin-R | Ankyrin-B | Ankyrin-G | |
|---|---|---|---|
| Tissue Expression | erythrocytes [1], myelinated axons [10], striated muscle [11] | ubiquitously expressed, cardiomyocytes (T-tubules, SR, plasma membrane) [12], neurons [8] | ubiquitously expressed, neurons (AIS, and nodes of Ranvier) [13], cardiomyocytes (intercalated disc) [14] |
| Examples of Binding Partners | CD44 [15], NKA [16], Rh type A glycoprotein [17], obscurin [11] | PP2A [12][13], NCX [12], NKA [18], Kir6.2 [12][13], CaV1.3 [19], βII-spectrin [20] | NaV1.6, βIV-spectrin, L1CAMs [1][21][22], plakophilin-2 [23] NaV1.5 [14] |
| Isoforms | sAnk1.5, 1.6, 1.7, and 1.9 [11] | AnkB-188 and AnkB-212 [24]. Giant AnkB (440-kD) | Giant AnkG (480-kD) [25] |
| Disease associated with variants | hereditary spherocytosis [26] | Ankyrin B syndrome: SCD, SND, AF, LQTS, VT, bradycardia, syncope [12], ARVC [27] | Brugada syndrome [12], dilated cardiomyopathy [28], cognitive disabilities [29] |
2. Ankyrin-B Structure and Binding Partners
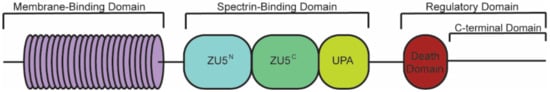
| Membrane-Binding Domain | Spectrin-Binding Domain | Regulatory Domain | |
|---|---|---|---|
| Ion channels | Transporters/Pumps | β-spectrin | HSP40 |
| IP3R | Anion Exchanger | PP2A | Obscurin |
| Cav1.3 | Na/Ca Exchanger | Ankyrin MBD | |
| Kir6.2 | Na/K ATPase | ||
| Structural | Cell adhesion | ||
| Tubulin β-catenin | L1CAMs | ||
| β-dystroglycan | |||
| Dystrophin | |||
3. Ankyrin-B Variants in Cardiovascular Disease
This entry is adapted from the peer-reviewed paper 10.3390/biom10020211
References
- Bennett, V.; Stenbuck, P.J. Identification and partial purification of ankyrin, the high affinity membrane attachment site for human erythrocyte spectrin. J. Biol. Chem. 1979, 254, 2533–2541.
- Bennett, V.; Davis, J.; Fowler, W.E. Immunoreactive forms of erythrocyte spectrin and ankyrin in brain. Philos. Trans. R. Soc. Lond. Ser. Biol. Sci. 1982, 299, 301–312.
- Bennett, V.; Davis, J. Spectrin and ankyrin in brain. Cell Motil. 1983, 3, 623–633.
- Davis, J.Q.; Bennett, V. Brain ankyrin. Purification of a 72,000 Mr spectrin-binding domain. J. Biol. Chem. 1984, 259, 1874–1881.
- Moon, R.T.; Ngai, J.; Wold, B.J.; Lazarides, E. Tissue-specific expression of distinct spectrin and ankyrin transcripts in erythroid and nonerythroid cells. J. Cell Biol. 1985, 100, 152–160.
- Nelson, W.J.; Lazarides, E. Posttranslational control of membrane-skeleton (ankyrin and alpha beta-spectrin) assembly in early myogenesis. J. Cell Biol. 1985, 100, 1726–1735.
- Otto, E.; Kunimoto, M.; McLaughlin, T.; Bennett, V. Isolation and characterization of cDNAs encoding human brain ankyrins reveal a family of alternatively spliced genes. J. Cell Biol. 1991, 114, 241–253.
- Davis, J.Q.; Bennett, V. Brain ankyrin. A membrane-associated protein with binding sites for spectrin, tubulin, and the cytoplasmic domain of the erythrocyte anion channel. J. Biol. Chem. 1984, 259, 13550–13559.
- Lambert, S.; Yu, H.; Prchal, J.T.; Lawler, J.; Ruff, P.; Speicher, D.; Cheung, M.C.; Kan, Y.W.; Palek, J. cDNA sequence for human erythrocyte ankyrin. Proc. Natl. Acad. Sci. USA 1990, 87, 1730–1734.
- Ho, T.S.-Y.; Zollinger, D.R.; Chang, K.-J.; Xu, M.; Cooper, E.C.; Stankewich, M.C.; Bennett, V.; Rasband, M.N. A hierarchy of ankyrin-spectrin complexes clusters sodium channels at nodes of Ranvier. Nat. Neurosci. 2014, 17, 1664–1672.
- Pierantozzi, E.; Szentesi, P.; Al-Gaadi, D.; Oláh, T.; Dienes, B.; Sztretye, M.; Rossi, D.; Sorrentino, V.; Csernoch, L. Calcium Homeostasis Is Modified in Skeletal Muscle Fibers of Small Ankyrin1 Knockout Mice. Int. J. Mol. Sci. 2019, 20, 3361.
- Curran, J.; Mohler, P.J. Coordinating electrical activity of the heart: Ankyrin polypeptides in human cardiac disease. Expert Opin. Targets 2011, 15, 789–801.
- Cunha, S.R.; Mohler, P.J. Ankyrin-based cellular pathways for cardiac ion channel and transporter targeting and regulation. Semin. Cell Dev. Biol. 2011, 22, 166–170.
- Makara, M.A.; Curran, J.; Little, S.C.; Musa, H.; Polina, I.; Smith, S.A.; Wright, P.J.; Unudurthi, S.D.; Snyder, J.; Bennett, V.; et al. Ankyrin-G coordinates intercalated disc signaling platform to regulate cardiac excitability in vivo. Circ. Res. 2014, 115, 929–938.
- Kalomiris, E.L.; Bourguignon, L.Y. Mouse T lymphoma cells contain a transmembrane glycoprotein (GP85) that binds ankyrin. J. Cell Biol. 1988, 106, 319–327.
- Nelson, W.J.; Veshnock, P.J. Ankyrin binding to (Na+ + K+)ATPase and implications for the organization of membrane domains in polarized cells. Nature 1987, 328, 533–536.
- Nicolas, V.; Le Van Kim, C.; Gane, P.; Birkenmeier, C.; Cartron, J.-P.; Colin, Y.; Mouro-Chanteloup, I. Rh-RhAG/Ankyrin-R, a New Interaction Site between the Membrane Bilayer and the Red Cell Skeleton, Is Impaired by Rhnull-associated Mutation. J. Biol. Chem. 2003, 278, 25526–25533.
- Skogestad, J.; Aronsen, J.M.; Tovsrud, N.; Wanichawan, P.; Hougen, K.; Stokke, M.K.; Carlson, C.R.; Sjaastad, I.; Sejersted, O.M.; Swift, F. Coupling of the Na+/K+-ATPase to Ankyrin B controls Na+/Ca2+ exchanger activity in cardiomyocytes. Cardiovasc. Res. 2019.
- Wolf, R.M.; Glynn, P.; Hashemi, S.; Zarei, K.; Mitchell, C.C.; Anderson, M.E.; Mohler, P.J.; Hund, T.J. Atrial fibrillation and sinus node dysfunction in human ankyrin-B syndrome: A computational analysis. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1253–H1266.
- Smith, S.A.; Sturm, A.C.; Curran, J.; Kline, C.F.; Little, S.C.; Bonilla, I.M.; Long, V.P.; Makara, M.; Polina, I.; Hughes, L.D.; et al. Dysfunction in the βII spectrin-dependent cytoskeleton underlies human arrhythmia. Circulation 2015, 131, 695–708.
- Kordeli, E.; Lambert, S.; Bennett, V. Ankyrin: A new ankyrin gene with neural-specific isoforms localized at the axonal initial segment and node of ranvier. J. Biol. Chem. 1995, 270, 2352–2359.
- Jenkins, S.M.; Bennett, V. Ankyrin-G coordinates assembly of the spectrin-based membrane skeleton, voltage-gated sodium channels, and L1 CAMs at Purkinje neuron initial segments. J. Cell Biol. 2001, 155, 739–746.
- Sato, P.Y.; Coombs, W.; Lin, X.; Nekrasova, O.; Green, K.J.; Isom, L.L.; Taffet, S.M.; Delmar, M. Interactions between ankyrin-G, Plakophilin-2, and Connexin43 at the cardiac intercalated disc. Circ. Res. 2011, 109, 193–201.
- Wu, H.C.; Yamankurt, G.; Luo, J.; Subramaniam, J.; Hashmi, S.S.; Hu, H.; Cunha, S.R. Identification and characterization of two ankyrin-B isoforms in mammalian heart. Cardiovasc. Res. 2015, 107, 466–477.
- Kunimoto, M.; Otto, E.; Bennett, V. A new 440-kD isoform is the major ankyrin in neonatal rat brain. J. Cell Biol. 1991, 115, 1319–1331.
- Perrotta, S.; Gallagher, P.G.; Mohandas, N. Hereditary spherocytosis. Lancet (Lond. Engl.) 2008, 372, 1411–1426.
- Roberts, J.D.; Murphy, N.P.; Hamilton, R.M.; Lubbers, E.R.; James, C.A.; Kline, C.F.; Gollob, M.H.; Krahn, A.D.; Sturm, A.C.; Musa, H.; et al. Ankyrin-B dysfunction predisposes to arrhythmogenic cardiomyopathy and is amenable to therapy. J. Clin. Investig. 2019, 129, 3171–3184.
- Makara, M.A.; Curran, J.; Lubbers, E.R.; Murphy, N.P.; Little, S.C.; Musa, H.; Smith, S.A.; Unudurthi, S.D.; Rajaram, M.V.S.; Janssen, P.M.L.; et al. Novel Mechanistic Roles for Ankyrin-G in Cardiac Remodeling and Heart Failure. Jacc Basic Transl. Sci. 2018, 3, 675–689.
- Iqbal, Z.; Vandeweyer, G.; van der Voet, M.; Waryah, A.M.; Zahoor, M.Y.; Besseling, J.A.; Roca, L.T.; Vulto-van Silfhout, A.T.; Nijhof, B.; Kramer, J.M.; et al. Homozygous and heterozygous disruptions of ANK3: At the crossroads of neurodevelopmental and psychiatric disorders. Hum. Mol. Genet. 2013, 22, 1960–1970.
- Bennett, V.; Stenbuck, P.J. The membrane attachment protein for spectrin is associated with band 3 in human erythrocyte membranes. Nature 1979, 280, 468–473.
- Narla, J.; Mohandas, N. Red cell membrane disorders. Int. J. Lab. Hematol. 2017, 39, 47–52.
- Satchwell, T.J.; Bell, A.J.; Hawley, B.R.; Pellegrin, S.; Mordue, K.E.; van Deursen, C.T.B.M.; Braak, N.H.-T.; Huls, G.; Leers, M.P.G.; Overwater, E.; et al. Severe Ankyrin-R deficiency results in impaired surface retention and lysosomal degradation of RhAG in human erythroblasts. Haematologica 2016, 101, 1018–1027.
- Bennett, V.; Baines, A.J. Spectrin and ankyrin-based pathways: Metazoan inventions for integrating cells into tissues. Physiol. Rev. 2001, 81, 1353–1392.
- Curran, J.; Mohler, P.J. Alternative paradigms for ion channelopathies: Disorders of ion channel membrane trafficking and posttranslational modification. Annu. Rev. Physiol. 2015, 77, 505–524.
- Jenkins, P.M.; Kim, N.; Jones, S.L.; Tseng, W.C.; Svitkina, T.M.; Yin, H.H.; Bennett, V. Giant ankyrin-G: A critical innovation in vertebrate evolution of fast and integrated neuronal signaling. Proc. Natl. Acad. Sci. USA 2015, 112, 957–964.
- Camors, E.; Mohler, P.J.; Bers, D.M.; Despa, S. Ankyrin-B reduction enhances Ca spark-mediated SR Ca release promoting cardiac myocyte arrhythmic activity. J. Mol. Cell Cardiol. 2012, 52, 1240–1248.
- Kashef, F.; Li, J.; Wright, P.; Snyder, J.; Suliman, F.; Kilic, A.; Higgins, R.S.D.; Anderson, M.E.; Binkley, P.F.; Hund, T.J.; et al. Ankyrin-B protein in heart failure: Identification of a new component of metazoan cardioprotection. J. Biol. Chem. 2012, 287, 30268–30281.
- He, M.; Tseng, W.-C.; Bennett, V. A single divergent exon inhibits ankyrin-B association with the plasma membrane. J. Biol. Chem. 2013, 288, 14769–14779.
- Michaely, P.; Tomchick, D.R.; Machius, M.; Anderson, R.G.W. Crystal structure of a 12 ANK repeat stack from human ankyrinR. Embo. J. 2002, 21, 6387–6396.
- Wang, C.; Yu, C.; Ye, F.; Wei, Z.; Zhang, M. Structure of the ZU5-ZU5-UPA-DD tandem of ankyrin-B reveals interaction surfaces necessary for ankyrin function. Proc. Natl. Acad. Sci. USA 2012, 109, 4822–4827.
- Zhu, W.; Wang, C.; Hu, J.; Wan, R.; Yu, J.; Xie, J.; Ma, J.; Guo, L.; Ge, J.; Qiu, Y.; et al. Ankyrin-B Q1283H Variant Linked to Arrhythmias Via Loss of Local Protein Phosphatase 2A Activity Causes Ryanodine Receptor Hyperphosphorylation. Circulation 2018, 138, 2682–2697.
- Qu, F.; Lorenzo, D.N.; King, S.J.; Brooks, R.; Bear, J.E.; Bennett, V. Ankyrin-B is a PI3P effector that promotes polarized α5β1-integrin recycling via recruiting RabGAP1L to early endosomes. Elife 2016, 5, e20417.
- Kontrogianni-Konstantopoulos, A.; Bloch, R.J. Obscurin: A multitasking muscle giant. J. Muscle Res. Cell Motil. 2005, 26, 419–426.
- Hall, T.G.; Bennett, V. Regulatory domains of erythrocyte ankyrin. J. Biol. Chem. 1987, 262, 10537–10545.
- Davis, L.H.; Davis, J.Q.; Bennett, V. Ankyrin regulation: An alternatively spliced segment of the regulatory domain functions as an intramolecular modulator. J. Biol. Chem. 1992, 267, 18966–18972.
- Lopes, L.R.; Syrris, P.; Guttmann, O.P.; O’Mahony, C.; Tang, H.C.; Dalageorgou, C.; Jenkins, S.; Hubank, M.; Monserrat, L.; McKenna, W.J.; et al. Novel genotype-phenotype associations demonstrated by high-throughput sequencing in patients with hypertrophic cardiomyopathy. Heart 2015, 101, 294–301.
- Ichikawa, M.; Aiba, T.; Ohno, S.; Shigemizu, D.; Ozawa, J.; Sonoda, K.; Fukuyama, M.; Itoh, H.; Miyamoto, Y.; Tsunoda, T.; et al. Phenotypic Variability of ANK2 Mutations in Patients With Inherited Primary Arrhythmia Syndromes. Circ. J. 2016, 80, 2435–2442.
- Robaei, D.; Ford, T.; Ooi, S.-Y. Ankyrin-B Syndrome: A Case of Sinus Node Dysfunction, Atrial Fibrillation and Prolonged QT in a Young Adult. Heartlung Circ. 2015, 24, e31–e34.
- Schott, J.J.; Charpentier, F.; Peltier, S.; Foley, P.; Drouin, E.; Bouhour, J.B.; Donnelly, P.; Vergnaud, G.; Bachner, L.; Moisan, J.P. Mapping of a gene for long QT syndrome to chromosome 4q25-27. Am. J. Hum. Genet. 1995, 57, 1114–1122.
- Swayne, L.A.; Murphy, N.P.; Asuri, S.; Chen, L.; Xu, X.; McIntosh, S.; Wang, C.; Lancione, P.J.; Roberts, J.D.; Kerr, C.; et al. Novel Variant in the ANK2 Membrane-Binding Domain Is Associated With Ankyrin-B Syndrome and Structural Heart Disease in a First Nations Population With a High Rate of Long QT Syndrome. Circ. Cardiovasc. Genet. 2017, 10, e001537.
- Huq, A.J.; Pertile, M.D.; Davis, A.M.; Landon, H.; James, P.A.; Kline, C.F.; Vohra, J.; Mohler, P.J.; Delatycki, M.B. A Novel Mechanism for Human Cardiac Ankyrin-B Syndrome due to Reciprocal Chromosomal Translocation. Heart Lung Circ. 2017, 26, 612–618.
- Corrado, D.; Link, M.S.; Calkins, H. Arrhythmogenic Right Ventricular Cardiomyopathy. N. Engl. J. Med. 2017, 376, 61–72.