Posterior capsule opacification (PCO) is the most common complication arising from the corrective surgery used to treat cataract patients. PCO arises when lens epithelial cells (LEC) residing in the capsular bag post-surgery undergo hyper-proliferation and transdifferentiation into myofibroblasts, migrating from the posterior capsule over the visual axis of the newly implanted intraocular lens (IOL). The developmental pathways underlying PCO are yet to be fully understood and the current literature is contradictory regarding the impact of the recognised risk factors of PCO. The aim of this review is firstly to collate the known biochemical pathways that lead to PCO development, providing an up-to-date chronological overview from surgery to established PCO formation. Secondly, the risk factors of PCO are evaluated, focussing on the impact of IOLs’ properties. Finally, the latest experimental model designs used in PCO research are discussed to demonstrate the ongoing development of clinical PCO models, the efficacy of newly developed IOL technology, and potential therapeutic interventions. This review will contribute to current PCO literature by presenting an updated overview of the known developmental pathways of PCO, an evaluation of the impact of the risk factors underlying its development, and the latest experimental models used to investigate PCO. Furthermore, the review should provide developmental routes for research into the investigation of potential therapeutic interventions and improvements in IOL design in the aid of preventing PCO for new and existing patients.
1. Introduction
Despite the histopathology of posterior capsule opacification (PCO) being well characterised, the molecular mechanisms underlying the pathology are still unknown [
1,
2,
3,
4]. In addition to this, current literature contains contradictions regarding the extent to which established risk factors impact PCO development. Herein, the known biomolecular pathways in PCO development are reviewed and a chronological overview of the mechanisms underlying the pathophysiology of PCO formation are presented. Moreover, this review explores the risk factors for PCO and considers their impact on PCO development. Lastly, this review examines the latest experimental models in PCO research used to investigate the next generation of medical and technological advancements for patients with PCO.
PCO is the most common complication arising from corrective surgery to treat cataracts [
1,
5,
6,
7]. In 2018, the World Health Organisation [WHO] estimated that 90 million people worldwide still live with cataract-associated blindness [
8]. Each year, an additional 1–2 million people become blind, of which 75% are treatable [
9]. This number is only expected to rise due to an increasingly expanding and ageing global population [
9]. The Royal National Institute of Blind People (RNIB) estimated that by 2020, 695,000 people would be living with cataracts in the United Kingdom, a number that would increase by 30% between 2020 and 2030. In England alone, 330,000 cataract surgeries are performed per year [
10]. Whilst in the UK cataract surgery is a routine, outpatient surgical procedure, a number of complications can occur for a subsection of patients. Approximately 20–50% of patients develop PCO and require further corrective treatment [
11].
PCO presents as a secondary cataract, an agglomeration of cells over the visual axis causing a loss of acuity. During surgery, the surgeon will create an opening in the capsular bag, known as capsulorhexis, and use phacoemulsification to remove the diseased, opaque lens and lens epithelial cells (LEC) from the capsular bag before implanting an artificial intraocular lens (IOL) [
12,
13]. The extent of LEC removal influences the propensity towards PCO development [
14,
15]. The initial inflammation caused by the surgical trauma may incite the hyper-proliferation, transdifferentiation, and migration of residual LECs [
16]. The transformed LECs migrate along the posterior capsule towards the anterior chamber to accumulate over the visual axis, forming a secondary cataract [
1,
17].
1.1. Pathophysiology of Posterior Capsule Opacification
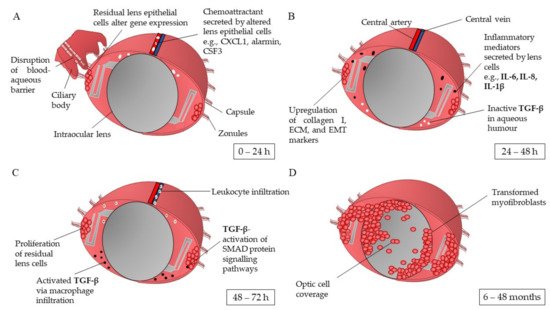
The wound healing response of LECs post-cataract surgery is believed to be the first key developmental stage of PCO. Ocular inflammation is triggered as a result of the opening incision and subsequent lens cell removal [
18,
19]. Jiang et al. [
16] showed that within the first 24 h post-surgery, the LEC transcriptome differentially expresses 19 of the 27 cataract-associated genes, some of which are markers of mesenchymal cell fate and are associated with chronic inflammatory conditions. The genes with the greatest upregulation in expression are
CXCL1, S100a9, CSF3/G-CSF, COX-2, CCL2, LCN2, and
HMOX1 [
20]. Some of these genes are involved in the production of proinflammatory chemokines such as CXCL1, alarmin S100a9, and G-CSF [
16]. This response may be initiated by the surgically induced break in the blood-aqueous barrier and subsequent leakage of plasma protein into the aqueous humour (A) [
16,
19]. At 48 h post operation, the elevated levels of inflammatory mediators interleukin (IL)-6, IL-1β, and IL-8 expressed by the LECs initiate and promote chronic inflammation pathways (B) [
5,
21,
22,
23,
24,
25]. Migrating neutrophils and macrophages attracted by the secreted chemokines cleave the inactive precursor of the latent transforming growth factor beta (TGF-β) via proteinases in the aqueous humour [
26,
27]. The activated TGF-β binds to LECs via type I and type II receptor serine-threonine kinases on the cell surface [
2]. This begins a cascade of SMAD proteins signalling, the messengers activated via TGFβ signalling [
28,
29]. These signalling pathways lead to TGF-β-gene transcription, activation of Rho GTPases, and stimulation of the PI3/Akt and MAPK pathways (C). These events are associated with myofibroblast formation, epithelial-mesenchymal-transition (EMT)-related matrix contraction, cell differentiation, and inhibition of normal LEC pathways [
2]. As a result, the LECs are stimulated to hyper-proliferate and differentiate into myofibroblasts, leading to their eventual migration over the visual axis of the IOL (D).
Figure 1. Schematic overview of the capsular bag post cataract surgery in the development of posterior capsule opacification. (
A) Stimulated by surgery trauma, residual lens epithelial cells undergo the wound healing response, alteration of gene transcriptome and expression of chemoattracts targeting inflammatory mediators and innate immune cells. (
B) Upregulation of inflammatory mediators, i.e., IL-1β expressed by lens cells undergoes autocrine signalling to initiate the synthesis of collagen I, extracellular matrix (ECM), and epithelial-mesenchymal transition (EMT), proteins and markers. (
C) Residual lens epithelial cells start to proliferate; leukocyte infiltration attracted by the high levels of chemo attractants and inflammatory mediators activate dormant transforming growth factor beta (TGF-β) residing in the aqueous humour. TGF-β activates SMAD3 signalling pathways in the lens cells, stimulating PI3/Akt, Rho GTPases, and MAPK pathways. (
D) The transdifferentiated lens cells migrate over the intraocular lens’ optic. IL: interleukin; TGF-β: transforming growth factor beta [
2,
5,
6,
16,
17,
20,
21,
25,
26,
37,
46].
From current literature, the hypothesised key mediators involved in PCO development are IL-1β, IL-8, IL-6, and TGF-β. IL-1β can be synthesised by retinal pigment epithelial cells, LECs, corneal epithelial cells, and to some extent, corneal stromal cells when triggered during inflammatory processes [
25,
30,
31]. IL-1β promotes inflammation by acting as a proinflammatory mediator, stimulating the secretion of other cytokines such as IL-8 [
24,
25,
32]. IL-8 can be produced by peripheral blood monocytes, endothelial cells, fibroblasts, and epithelial cells [
25]. A study conducted by Ferrick [
25] exemplifies the interconnectivity of IL-1β and IL-8 in augmenting the inflammatory response. Rat eye models were first injected with IL-1β. At 20 h post injection, IL-8-activated neutrophils were detected. The rat neutrophils levels then subsided after 48 h. However, this inflammatory resolution may not occur as quickly in humans. The study concluded that IL-1β produces a stronger inflammatory response with long-lasting effects due to the cascade of pathways it initiates such as activating leukocytes, instigating proinflammatory mechanisms in local cells and promoting the synthesis of cytokines such as IL-8, whereas IL-8 acts as a specific chemoattractant for neutrophils.
The presence of IL-6 during PCO development is supported by the study conducted by Nishi [
23]. The authors found IL-6 expression to be significantly higher in the aqueous humour of LECs obtained during cataract surgery in comparison to medium controls. IL-6 is believed to be involved in the upregulation of extracellular matrix (ECM) synthesis, the contraction of the capsular bag, and EMT of LECs, as shown by Ma [
5]. Within 24 h of IL-6 stimulation, the human LEC-B3 (HLE-B3) line initiated a significant increase in ECM synthesis and activation of the JAK/STAT3 pathway was observed [
5]. Cells treated with JAK/STAT3 inhibitor WP1066 showed significantly inhibited expression of collagen, fibronectin, and TGF-β2 (
p > 0.01). This suggests that the JAK/STAT3 pathway influences the expression of ECM proteins in HLE-B3 cells. The authors also showed that IL-6 works in synergy with TGF-β to promote EMT, since HLE-B3 cells treated with IL-6 and TGF-β2 showed significantly increased expression of the markers of EMT, alpha smooth muscle actin (α-SMA), collagen, and fibronectin in comparison to cells treated with either IL-6 or TGF-β2.
TGF-β is a well-established key mediator within PCO development [
1,
6,
16,
17,
29,
33,
34]. The isoforms TGF-β1 and -β2 are found in ciliary processes and within the limbal epithelium where they are hypothesised to be involved in the transdifferentiation of conjunctival to corneal epithelium. As a consequence, it is believed these isoforms are locally synthesised due to their predominance in the aqueous humour [
6,
35]. Moreover, Nishi [
36] found that LECs express TGF-β2. The binding of TGF-β to its receptor activates kinase domains within the receptor, augmenting phosphorylation cascades which initiate SMAD3 transcription factors [
2,
28]. The TGF-β receptor kinase has many roles within the ocular tissue, in both normal and pathological conditions [
37,
38]. TGF-β is important in maintaining corneal integrity, in tissue repair, and in the regulation of cell proliferation and death [
37,
38]. Nevertheless, hyper-activation of the growth factor can lead to an exaggerated wound-healing response and increased ECM deposition by cells, which may cause fibrotic corneal disease, fibrosis of lens and retinal epithelium, and loss of vision [
6,
28,
35,
37].
Short-term exposure to TGF-β can lead to long-term impact. Possible mechanisms behind this are the positive feedback loop on the upregulation of TGF-β gene expression stimulated by the exposure to TGF-β or the ability of TGF-β to bind to collagen IV, a predominant component of the capsular bag. In PCO development, TGF-β is also involved in the activation of the EMT pathway in LEC; the differentiation of the epithelial cells to migratory spindle-like myofibroblasts [
29,
39,
40,
41,
42]. The activation of TGF-β inhibits the proliferation of LECs in favour of initiating transdifferentiation by upregulating the expression of α-SMA genes [
6,
28,
43]. Therefore, α-SMA can be used as a biomarker for myofibroblasts [
44]. Furthermore, TGF-β has been shown to increase the secretion of ECM components by LECs. In this process, ECM turnover is disturbed; the upregulation of fibronectin, collagen I and IV, and inhibitors of matrix metalloproteinases (MMPs) cause a greater production of ECM with reduced degradation. The increased ECM leads to capsule wrinkling and thickening of the posterior capsule [
6,
43,
45]. The presentation of PCO can differ between individuals, depending on patient specific risk factors affecting the patients.
1.2. Risk Factors for Developing Posterior Capsule Opacification
PCO typically develops in the first 2–5 years post-surgery [
7,
11,
14,
47,
48]. There are several risk factors that can make patients more susceptible to developing PCO. These are patient-associated risk, surgical-associated risk, and IOL-associated risk, as discussed.
1.3. Therapeutic Interventions for Posterior Capsule Opacification
PCO is managed with Nd:YAG laser capsulotomy. The agglomerated cells are targeted, creating an opening which restores visual acuity. A retrospective study of 806 patients found the requirement for Nd:YAG capsulotomy was 10.6% after one year, 14.8% after two, and 28.6% after four years [
89]. The procedure can lead to complications such as retinal detachment, cystoid macular oedema, IOL displacement, mild anterior uveitis, and transient intraocular pressure [
90]. Developing preventative measures to inhibit the biomolecular pathways of PCO could reduce the risk to patients, lower the requirement for additional treatment post cataract surgery, and ease the burden on healthcare services. Thus far, improvements in surgical technique and the square edge optic design have shown a reduction in PCO incidence [
4,
9,
72,
76,
91]. Non-pharmacological methods to reduce PCO development include gene therapy and inducing osmotic changes in LECs [
2]. Pharmacological approaches to preventing PCO include cytostatic drugs, anti-inflammatory drugs, and antagonists to key molecules within the developmental pathways of PCO as exemplified by Shao [
92]. Shao [
92] stimulated the HLE-B3 cell line SRA01/04 with TGF-β2 to induce EMT then treated the cells with fasudil, an inhibitor of the Rho-kinase activated during TGF-β signalling. Fasudil significantly reduced cell proliferation and migration, down-regulated α-SMA expression, and prevented the suppression of epithelial marker Connexin43. Nevertheless, despite extensive research into therapeutic solutions for PCO, no clinical treatment exists other than Nd:YAG.
1.4. Capsular Devices to Prevent Posterior Capsule Opacification Development
Capsular devices were developed to facilitate cataract surgery by improving capsular bag stability and intraocular lens centration [
93,
94]. The introduction of the capsular device by Hara [
95] in 1991 has since led to the evolution of endocapsular devices that have aided capsular support and stability leading to a subsequent reduction in PCO development. The equator ring (E-ring) introduced by Hara [
95] was a closed silicone circle with square-edge design and an inner groove to allow IOL fixation. The squared edges were hypothesised to delay PCO formation. Hara [
96] compared 51 eyes, 14 of which were implanted with the E-ring, and found the E-ring significantly reduced PCO development. This design was built upon by Nishi who developed a capsular tension ring (CTR) made from PMMA with sharp rectangular edges. Although the implantation of a CTR has shown reduced PCO formation in comparison to patients implanted with an IOL alone, the discontinuous capsular bend created allows the possibility for LEC migration [
93,
97,
98]. This CTR design was adapted into an open-capsule device for the purpose of expanding and opening the capsular bag, separating the anterior and posterior capsules [
93,
99]. The open-capsule device can be made of hydrophobic or hydrophilic material and is a closed ring with a square-edge design. Alon [
100] showed that the open-capsule ring reduced PCO development, regardless of base material. New Zealand white rabbits were assigned either to two control groups, each implanted with hydrophobic or hydrophilic IOLs without an open-capsule ring, or to four study groups, each implanted with a hydrophobic or hydrophilic IOL and a hydrophobic or hydrophilic open-capsule ring. A clinical evaluation found a 69% reduction in the eyes implanted with the device relative to the control eyes. This reduction has been linked to the 360° squared edge and the delivery of aqueous humour to the capsule equator through windows in the rim of the device that prevents LEC migration and proliferation, respectively [
93]. An additional endocapsular device was developed by Sharklet Technologies, Inc. (Aurora, CO, USA) to act as an artificial capsular bag. The protective silicone membrane has a square-edge haptic ring which provides a ridge for the IOL haptics. The signature feature of this design is the composite sharkskin-inspired microtopography which inhibits bioadhesion and has been hypothesised to prevent LEC migration [
93,
101]. These capsular devices, in addition to the other risk factors discussed, present the complexity of understanding and treating PCO. Experimental models are utilised in PCO research for insight into the underlying pathophysiology, testing newly developed IOL technology in aid of device optimisation, and investigating potential therapeutic interventions.
2. Experimental Models to Investigate Posterior Capsule Opacification
2.1. In Vitro Models
Whilst in vitro models are not representative of the physiological environment they have been used as a rapid and cost-effective route to ascertain the molecular mechanisms underlying PCO formation. Additionally, cell models are typically used in the first stage of investigating possible therapeutic interventions due to the reduced ethical constraints and ability to provide fundamental toxicology data prior to using relatively more expensive in vivo and ex vivo models. Cell culture models have shown the role of fibronectin, growth factors such as TGF-β, and enzymes including aldose reductase in the development of PCO [
102,
103,
104]. The creation of three-dimensional cell culture models has broadened the application of in vitro models to encompass structural replicability.
2.2. In Vivo Models
In vivo models can be exploited in many aspects of PCO research. Such applications include the investigation of intraocular lenses, underlying biomolecular mechanisms, and the efficacy of surgical interventions, and the testing of therapeutic inhibitors of developmental pathways [
17,
109,
110,
111,
112]. Animal donors include murine, rabbit, and porcine species. However, caution is required when comparing and extrapolating pathological responses between animals and humans due to differences in species biology [
1,
17]. Furthermore, each animal donor type has its own limitations.
2.3. Ex Vivo Models
Ex vivo models have a benefit over in vivo models as the explant system maintains the natural structure of the surrounding tissue; therefore, these models provide insight into the localised impact of treatments with less stringent ethical constraints [
17,
120]. A study conducted by Kassumeh [
121] exemplifies this as this study investigated slow-releasing methotrexate (MTX)-loaded poly (lactic-co-glycolic) biomatrices as a IOL coating to reduce PCO. MTX was selected as the best candidate from a review of potential pharmacological drugs that were cross-referenced against those approved by the FDA or EMA. The study found no toxic effects on the corneal cell line CEC-SV40. Moreover, the study performed open-sky cataract surgery on twelve cadaver eyes then implanted the coated IOLs. The capsular bags were kept pinned in culture flasks and kept under standard cell culture conditions. At 9.3 days, the control IOLs showed full cell coverage whereas at this time point, the MTX-coated IOLs showed residual LECs were only visible at the outer edges and took 51.0–51.3 days to achieve full coverage. The application of ex vivo models is further supported by D’Antin [
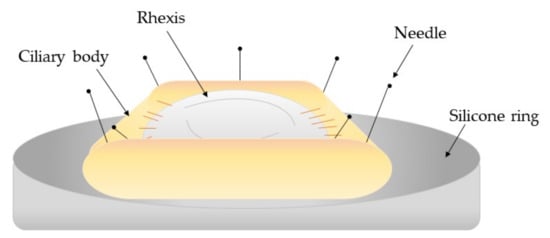
122] who used human eye donor tissue to investigate the treatment of hydrogen peroxide and distilled water to prevent cell proliferation within PCO development. The capsule bag-ciliary body complex was dissected from the donor eyes and transferred to sterile petri dishes onto a silicone ring mould where the iris was removed. Continuous circular capsulorrhexis, hydrodissection, and hydroexpression was performed before individual treatment using a silicone irrigation ring (). The study found that both treatments significantly delayed cell growth by day 28 whereas the control donor eyes reached cell confluence by day 11.
Figure 2. Schematic diagram of the capsule-ciliary body complex attached to a silicone ring mount [
122].
In contrast, Taiyab [
29] used rat ex vivo models to investigate the role of TGF-β in the EMT pathway within PCO development. In their first study, the explant models were treated with TGF-β with or without SIS3, a SMAD3 inhibitor. The untreated samples showed no α-SMA and stress F-actin fibre expression whereas lens explants treated with TGF-β displayed a greater staining of these markers. The explants treated with TGF-β and SIS3 reduced the expression of α-SMA and stress F-actin fibre expression which corresponded with the positive control ICG-001, an inhibitor of TGF-β-induced EMT. Following on from this work, the study investigated the E-cadherin/β-cadherin complex within the cell periphery that undergoes rearrangement during EMT-related PCO. On treating the explants with TGF-β, a reduction in E-cadherin and increased nuclear translocation of β-catenin was observed which led to a loss in cell structure. With the addition of SIS3, the staining of E-cadherin did not alter in comparison to the untreated control explants and only a partial reduction in β-catenin nuclear translocation was seen.
Ex vivo models have also been used to improve the reproduction of the clinical environment of PCO as shown by Eldred [
123] who used a graded culture media in a human capsular bag model to assess the impact of IOL design on its propensity to initiate PCO development. Mock surgeries were completed on donor human eye tissue after which the eyes were implanted with either Hoya Vivinex or Alcon Acrysof lenses. The capsular bag and surrounding ciliary tissue were secured to a silicone ring within a culture disk. The capsular bag was kept in a graded culture system to reflect the clinical environment, beginning with 5% human serum and 10 ng/mL TGF-β2 and reducing to serum-free media by day 15–28. A matched pair design was used to assess the implanted IOL and graded culture serum. The findings showed that the graded culture produced quicker posterior coverage, matrix contraction, and significantly increased light scatter. Cell growth over the central optic was greater in the Alcon Acrysof and significantly increased light scatter occurred.
Experimental models provide inexpensive, readily available replicas of the ocular tissue environment to reproduce the pathology of PCO and investigate therapeutic interventions and new IOL technology. Nevertheless, the differences in the wound-healing response and inflammation pathways between species prevents direct comparison to human studies. Experimental models can be exploited prior to clinical studies and present alternative, easily accessible methods for scientific investigation; however, clinical studies utilise a longitudinal design to provide important insight into the dissemination of PCO and the long-term risk factors of novel medical devices and treatments.
2.4. Clinical Studies
Building upon the evidence of preliminary experimental models, clinical studies provide a final evaluation of newly developed IOL technology or therapeutic interventions by using human participants and longitudinal reviews [
124]. Clinical studies have been used in PCO research not only for the previously discussed applications but also in determining distribution of PCO and Nd:YAG capsulotomy rates. Nevertheless, these studies also present their own limitations and their specific scope or population demographic can be difficult to translate to the general population, preventing broad applicability. The different study designs and outcomes of recent clinical studies are discussed hereafter.
3. Conclusions
In conclusion, this review presented a revised overview of the developmental pathways of PCO, from the initial wound-healing response of the LEC to the formation of a secondary cataract. Additionally, the review examined the individual impact of the risk factors underlying PCO development, establishing the square optic edge as an effective design to prevent PCO and provided clarity on the current understanding on the impact of the IOL material’s wettability. Finally, this review demonstrated the latest experimental models used in PCO research and how the model types can be exploited for different applications. Future work should focus on developing current understanding of the pathophysiology of PCO, providing in-depth analysis of the risk factors leading to PCO, specifically IOL design and function and conducting clinical studies with larger population sizes to ensure reviews provide valid and appropriate conclusions.
This entry is adapted from the peer-reviewed paper 10.3390/pharmaceutics13060860