The microbial diversity in anaerobic digestion (AD) is important because it affects process robustness. High-throughput sequencing offers high-resolution data regarding the microbial diversity and robustness of biological systems including AD; however, to understand the dynamics of microbial processes, knowing the microbial diversity is not adequate alone. Advanced meta-omic techniques have been established to determine the activity and interactions among organisms in biological processes like AD. Results of these methods can be used to identify biomarkers for AD states. This can aid a better understanding of system dynamics and be applied to producing comprehensive models for AD.
- microbial diversity
- next generation sequencing
- proteomics
- metabolomics
- community diversity
- anaerobic digetion
- biogas
1. Introduction
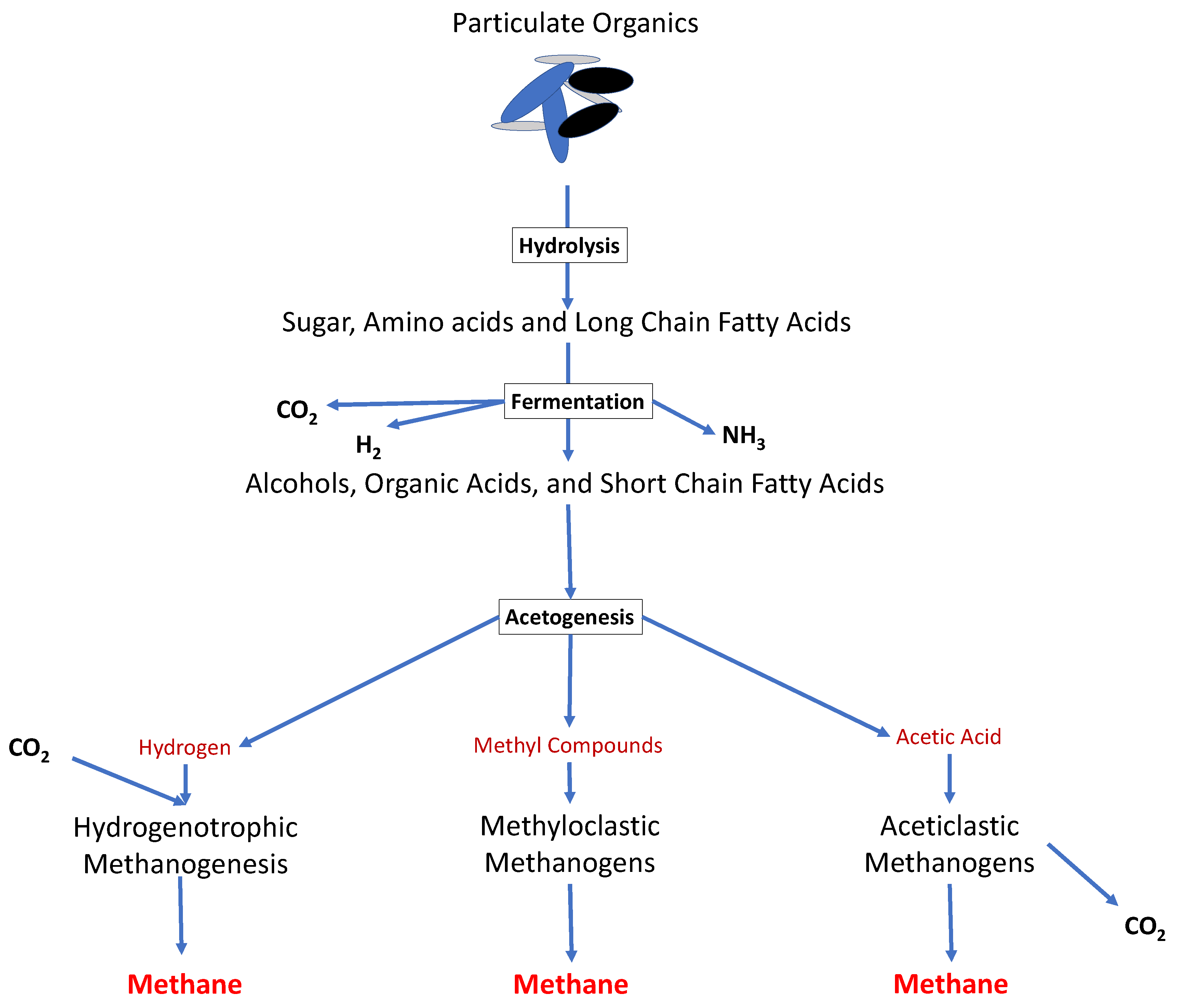 Figure 1. The overall metabolic pathways of anaerobic digestion. Polymeric organic materials go through the four stages of anaerobic digestion (hydrolysis, acidogenesis, acetogenesis and methanogenesis), in order to produce raw biogas.
Figure 1. The overall metabolic pathways of anaerobic digestion. Polymeric organic materials go through the four stages of anaerobic digestion (hydrolysis, acidogenesis, acetogenesis and methanogenesis), in order to produce raw biogas.2. Application of Microbial Diversity Analysis in AD Models
2.1. Metabolic Models of AD
2.2. Development of Metabolic Models and Main Gaps in the Field
2.3. Examples of Hybrid Cellular-Level Modeling/Biochemical Process Modeling (CLM/BPM) for Enhanced Predictivity of AD Models
3. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms9061162
References
- Su, C.; Lei, L.; Duan, Y.; Zhang, K.-Q.; Yang, J. Culture-independent methods for studying environmental microorganisms: Methods, application, and perspective. Appl. Microbiol. Biotechnol. 2012, 93, 993–1003.
- Lamb, J.J. Microbiology of AD. In Anaerobic Digestion: From Biomass to Biogas; SCIO Publishing: Trondheim, Norway, 2020; pp. 53–88.
- Gregorie, E.F.J.; Lamb, J.J.; Lien, K.M.; Pollet, B.G.; Burheim, O.S. Hydrogen and biogas. In Micro-Optics and Energy: Sensors for Energy Devices; Springer: Berlin/Heidelberg, Germany, 2020.
- Metcalf, I.; Eddy, G.; Tchobanoglous, H.; Stensel, R.; Tsuchihashi, F. Burton, Wastewater Engineering: Treatment and Resource Recovery, 5th ed.; McGraw-Hill: New York, NY, USA, 2014; Volume 1.
- Sarker, S.; Nordgård, A.S.; Lamb, J.J.; Lien, K.M. Biogas and hydrogen. In Hydrogen, Biomass and Bioenergy; Academic Press: Cambridge, MA, USA, 2020.
- Ziganshin, A.M.; Liebetrau, J.; Pröter, J.; Kleinsteuber, S. Microbial community structure and dynamics during anaerobic digestion of various agricultural waste materials. Appl. Microbiol. Biotechnol. 2013, 97, 5161–5174.
- Zamri, M.; Hasmady, S.; Akhiar, A.; Ideris, F.; Shamsuddin, A.; Mofijur, M.; Fattah, I.M.R.; Mahlia, T. A comprehensive review on anaerobic digestion of organic fraction of municipal solid waste. Renew. Sustain. Energy Rev. 2021, 137, 110637.
- De Vrieze, J.; Pinto, A.J.; Sloan, W.T.; Ijaz, U.Z. The active microbial community more accurately reflects the anaerobic digestion process: 16S rRNA (gene) sequencing as a predictive tool. Microbiome 2018, 6, 1–13.
- Rudakiya, D.M.; Narra, M. Microbial Community Dynamics in Anaerobic Digesters for Biogas Production; Springer: Singapore, 2021; pp. 143–159.
- Lamb, J.J.; Hill, R.E.; Eaton-Rye, J.J.; Hohmann-Marriott, M.F. Functional role of PilA in iron acquisition in the cyanobacterium Synechocystis sp. PCC 6803. PLoS ONE 2014, 9, e105761.
- Lamb, J.J.; Hohmann-Marriott, M.F. Manganese acquisition is facilitated by PilA in the cyanobacterium Synechocystis sp. PCC 6803. PLoS ONE 2017, 12, e0184685.
- Vanwonterghem, I.; Jensen, P.D.; Ho, D.P.; Batstone, D.J.; Tyson, G.W. Linking microbial community structure, interactions and function in anaerobic digesters using new molecular techniques. Curr. Opin. Biotechnol. 2014, 27, 55–64.
- Gaspari, M.; Treu, L.; Zhu, X.; Palù, M.; Angelidaki, I.; Campanaro, S.; Kougias, P.G. Microbial dynamics in biogas digesters treating lipid-rich substrates via genome-centric metagenomics. Sci. Total Environ. 2021, 778, 146296.
- Nelson, M.C.; Morrison, M.; Yu, Z. A meta-analysis of the microbial diversity observed in anaerobic digesters. Bioresour. Technol. 2011, 102, 3730–3739.
- Sundberg, C.; Abu Al-Soud, W.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626.
- Lyberatos, G.; Skiadas, I.V. Modeling of Anaerobic Digestion—A Review. Glob. NEST Int. J. 1999, 1, 63–76.
- Batstone, D.; Hülsen, T.; Oehmen, A. Metabolic modelling of mixed culture anaerobic microbial processes. Curr. Opin. Biotechnol. 2019, 57, 137–144.
- Kim, W.J.; Kim, H.U.; Lee, S.Y. Current state and applications of microbial genome-scale metabolic models. Curr. Opin. Syst. Biol. 2017, 2, 10–18.
- González-Cabaleiro, R.; Lema, J.M.; Rodríguez, J. Metabolic energy-based modelling explains product yielding in anaerobic mixed culture fermentations. PLoS ONE 2015, 10, e0126739.
- Kim, T.Y.; Sohn, S.B.; Bin Kim, Y.; Kim, W.J.; Lee, S.Y. Recent advances in reconstruction and applications of genome-scale metabolic models. Curr. Opin. Biotechnol. 2012, 23, 617–623.
- Thor, S.; Peterson, J.R.; Luthey-Schulten, Z. Genome-Scale Metabolic Modeling of Archaea Lends Insight into Diversity of Metabolic Function. Archaea 2017, 2017, 1–18.
- Angelidaki, I.; Ellegaard, L.; Ahring, B.K. A comprehensive model of anaerobic bioconversion of complex substrates to biogas. Biotechnol. Bioeng. 1999, 63, 363–372.
- Henze, M.; Gujer, W.; Mino, T.; Van Loosedrecht, M. Activated Sludge Models ASM1, ASM2, ASM2d and ASM3; IWA Publishing: London, UK, 2006; Volume 5.
- Batstone, D.; Keller, J.; Angelidaki, I.; Kalyuzhnyi, S.; Pavlostathis, S.; Rozzi, A.; Sanders, W.; Siegrist, H.; Vavilin, V. The IWA Anaerobic Digestion Model No 1 (ADM1). Water Sci. Technol. 2002, 45, 65–73.
- Cai, Y.; Zheng, Z.; Wang, X. Obstacles faced by methanogenic archaea originating from substrate-driven toxicants in anaerobic digestion. J. Hazard. Mater. 2021, 403, 123938.
- Kalyuzhnyi, S.; Fedorovich, V. Mathematical modelling of competition between sulphate reduction and methanogenesis in anaerobic reactors. Bioresour. Technol. 1998, 65, 227–242.
- Regueira, A.; González-Cabaleiro, R.; Ofiţeru, I.; Rodríguez, J.; Lema, J. Electron bifurcation mechanism and homoacetogenesis explain products yields in mixed culture anaerobic fermentations. Water Res. 2018, 141, 349–356.
- Mosey, F.E. Mathematical modelling of the anaerobic digestion process: Regulatory mechanisms for the formation of short-chain volatile acids from glucose. Water Sci. Technol. 1983, 15, 209–232.
- Hoelzle, R.D.; Virdis, B.; Batstone, D.J. Regulation mechanisms in mixed and pure culture microbial fermentation. Biotechnol. Bioeng. 2014, 111, 2139–2154.
- Ahmed, W.; Rodríguez, J. Modelling sulfate reduction in anaerobic digestion: Complexity evaluation and parameter calibration. Water Res. 2018, 130, 255–262.
- TStorck, T.; Virdis, B.; Batstone, D.J. Modelling extracellular limitations for mediated versus direct interspecies electron transfer. ISME J. 2016, 10, 621–631.
- Ramirez, I.; Volcke, E.I.; Rajinikanth, R.; Steyer, J.-P. Modeling microbial diversity in anaerobic digestion through an extended ADM1 model. Water Res. 2009, 43, 2787–2800.
- Weinrich, S.; Koch, S.; Bonk, F.; Popp, D.; Benndorf, D.; Klamt, S.; Centler, F. Augmenting biogas process modeling by resolving intracellular metabolic activity. Front. Microbiol. 2019, 10, 1095.
- Abram, F.; Enright, A.-M.; O’Reilly, J.; Botting, C.; Collins, G.; O’Flaherty, V. A metaproteomic approach gives functional insights into anaerobic digestion. J. Appl. Microbiol. 2011, 110, 1550–1560.
- Hanreich, A.; Schimpf, U.; Zakrzewski, M.; Schlüter, A.; Benndorf, D.; Heyer, R.; Rapp, E.; Pühler, A.; Reichl, U.; Klocke, M. Metagenome and metaproteome analyses of microbial communities in mesophilic biogas-producing anaerobic batch fermentations indicate concerted plant carbohydrate degradation. Syst. Appl. Microbiol. 2013, 36, 330–338.
- Jimenez, J.; Charnier, C.; Kouas, M.; Latrille, E.; Torrijos, M.; Harmand, J.; Patureau, D.; Sperandio, M.; Morgenroth, E.; Béline, F.; et al. Modelling hydrolysis: Simultaneous versus sequential biodegradation of the hydrolysable fractions. Waste Manag. 2020, 101, 150–160.