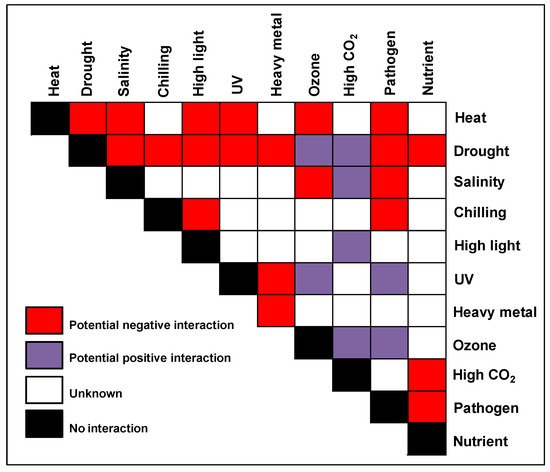
| Negative response |
Drought + salinity |
Wheat |
|
[54] |
| |
|
Maize |
|
[55] |
| |
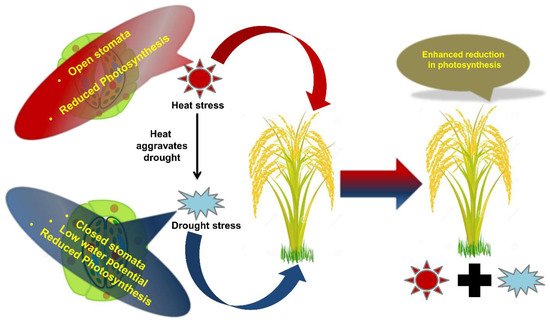
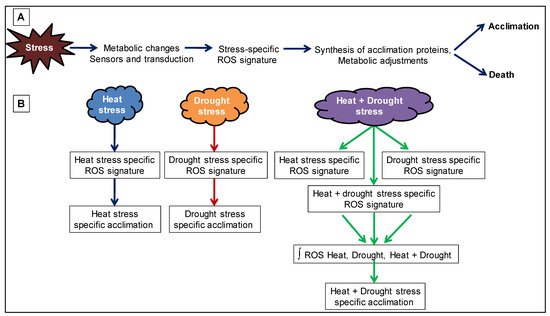
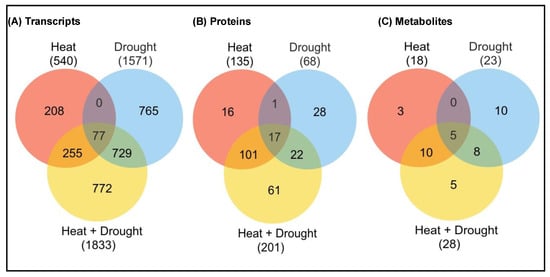
Drought + high temperature |
Tobacco |
-
Stomatal closure, reduced photosynthesis, enhanced respiration, and leaf temperature.
-
Some transcripts induced during individual stress while suppressed during combined stress. Few transcripts were specifically induced during combined stress.
-
Overlap between different transcription factors during individual and combined stress.
|
[16] |
| |
|
Arabidopsis |
-
454 transcripts were specifically expressed during combined stress.
-
Sucrose, maltose and glucose were highly accumulated under combined stress.
-
Proline only accumulated during drought stress.
|
[17] |
| |
|
Wheat |
-
Photosynthesis rate declined under High temperature > Drought > combined stress.
-
High temperature significantly affects grain number, while drought affects grain weight and combined stress affects leaf chlorophyll content, spikelet fertility, total dry weight, and harvest index.
|
[8] |
| |
|
Arabidopsis |
-
Root allocation increased during drought, while reproductive allocation, hyponasty and specific leaf area increased under high temperature.
-
Origin of accession plays a significant role during individual and combined stress.
|
[56] |
| |
|
Maize |
-
Combined stress in comparison to a single stress strongly affected the seminal lateral roots, reducing the dry weight, length, surface area and root mass ratio (RMR).
|
[57] |
| |
Drought + chilling |
Sugarcane |
-
Effect of abiotic stress is cultivar-dependent where the sensitive genotypes were more affected by combined stress than tolerant genotypes.
-
Low root temperature combined with drought severely affects PSII activity.
|
[58] |
| |
Drought + pathogen |
Arabidopsis |
-
Interaction among ABA, JA, and ethylene signaling pathways regulate pathogen-, wound-, and dehydration-response and one signaling pathway may dominate over others, depending on the stress conditions.
|
[59] |
| |
|
Arabidopsis |
-
11 genes differentially regulated, 23 genes specifically regulated, and reduced expression of R-gene mediated response were observed under combined heat, drought, and turnip mosaic virus stress.
|
[60] |
| |
Drought + UV |
Plants |
|
[61] |
| |
Drought + high light |
Arabidopsis |
-
Mutant seedlings deficient in alternative oxidase (AOX) showed accumulation of anthocyanins in leaves, alterations in photosynthetic efficiency, increased superoxide radical and reduced root growthunder combined stress.
|
[62] |
| |
Drought + low N |
Wheat |
-
Low N stress can lead to accumulation of ABA in wheat seedlings.
-
Combined stress was found to have significant interaction in regulation of genes for nitrate signaling, uptake and assimilation.
|
[63] |
| |
Drought + heavy metals |
Red maple |
-
Combined stress has additive effect in both stems and roots, reductions in hydraulic conductance, xylem-specific conductivity, and leaf-specific conductivity.
|
[64] |
| |
Drought + soil compaction + mechanical stress |
Tobacco |
|
[65] |
| |
Drought + nutrient |
Mungbean |
-
Under combined stress, a significant reduction in gas exchange traits (photosynthesis, stomatal conductance, transpiration, instantaneous water use efficiency), and P uptake in seed and shoot were observed under combined stress as compared to individual stress.
|
[66] |
| |
Salinity +high/low temperature
Drought + high/low temperature |
Wheat |
-
Root and shoot elongation significantly reduced under individual stress.
-
HT/LT treatment possess additive effect on growth inhibition under salt stress.
-
α-tocopherol significantly increased under drought and salt stress but significantly decreased under HT stress.
|
[67] |
| |
Salinity + high temperature |
Suaeda salsa |
-
Combined stress suppressed CO2 assimilation and photosystem II efficiency.
-
57 differentially expressed proteins were observed under individual and combined stress.
-
Expression of nucleoside diphosphate kinase 1, chlorophyll a/b binding protein, and ABC transporter I family member 1 was specifically induced during combined stress.
|
[68] |
| |
Salinity + pathogen |
Rice |
-
Downregulation of OsMAPK5 expression enhanced pathogenesis-related (PR) genes expression and significantly enhanced resistance to fungal (Magnaporthe grisea) and bacterial (Burkholderia glumae) pathogens but reduced tolerance to drought, salt, and cold. In contrast, overexpression lines exhibited increased OsMAPK5 kinase activity and increased tolerance to drought, salt, and cold stresses.
|
[69] |
| |
High temperature + ozone |
Silver birch (Betula pendula) |
-
O3 reduces, while temperature increase tree growth and growth may be counteractive during combined stress. R:S ratio decreases under O3 exposure.
-
Temperature increase may stimulate soil respiration rates and total biomass, while O3 could have opposite effect.
-
Elevated O3 decreases C assimilation, foliar C content and productivity.
|
[70] |
| |
High temperature + pathogen |
Arabidopsis |
-
NB-LRR type of R or R-like protein is the temperature-sensitive component of plant defense responses.
-
Alterations in the R-like gene SNC1 and the R gene N can change temperature sensitivity of defense responses.
|
[71] |
| |
High temperature + UV-C |
Strawberry |
-
Both heat and combined treatments, decreased hue and delayed changes in the colorimetric parameters.
-
The combined stress treatment reduced fungal infections and delayed in vitro germination of Botrytis cinerea conidia.
-
Neither the heat nor UV-C irradiation modified the total sugar content, although the combined treatment decreased it slightly relative to the control.
-
The combination of UV-C and heat treatments enhanced the benefits of applying each treatment separately and could be useful to improve and extend strawberry fruit postharvest life.
|
[72] |
| |
High temperature + high light |
Sunflower
(Helianthus annuus) |
-
Comparative expression analysis of leaves and immature seeds revealed that 89, 113 and 186 genes were differentially expressed in response to HL, HT and HL + HT, respectively.
|
[73] |
| |
High temperature + CO2 |
Soybean and maize |
-
Increased photosynthetic rates in response to CO2 enrichment, while C4 cycle is largely unresponsive to increased response to CO2 enrichment.
-
CO2 enrichment can mitigate the effects of moderately elevated temperatures on crop yield.
|
[74] |
| |
Low temperature + pathogen |
Plants |
-
Both virus and transgene-triggered RNA silencing are inhibited at low temperature. Thus, plants become more susceptible to viruses. RNA silencing-based phenotypes of transgenic plants are lost. However, temperature does not influence the accumulation of micro (mi) RNAs, which play a role in developmental regulation.
|
[75] |
| |
Low temperature + high light |
Dunaliella salina |
-
Low temperature and combined high light-low temperature decreased chlorophyll and β-carotene indicating that these treatments cause photo-oxidative stress.
-
High light, low temperature and combined high light-low temperature treatments increased the total ascorbate pool by 10–50% and the total glutathione pool by 20–100% with no consistent effect on their redox state.
|
[76] |
| |
Pathogen + nutrient |
Arabidopsis |
|
[77] |
| |
UV-B + Heavy metals |
Pea |
-
Combined dose (UV-B + 0.01 mM Ni) caused inhibitory effects.
-
Nickel at high doses strongly inhibited PSII activity and the inhibition was further intensified when chloroplasts were simultaneously exposed to UV-B radiation.
-
High doses of Ni (0.1 and 1.0 mM) and UV-B alone interrupted electron flow at the oxygen evolving complex. Similar damaging effects were caused by 0.01 and 0.1 mM Ni together with UV-B, but the damage extended to PSII reaction center, in case of 1.0 mM Ni in combination with UV-B.
|
[78] |
| |
Nutrient + high CO2 |
Panicum maximum Jacq. ‘Mombaça’ (Guinea grass) |
|
[79] |
| |
Heavy metals + heavy metals |
Tomato |
-
Zn supply clearly reduced Cd accumulation in leaves and simultaneously increased Zn concentration.
-
Cd-induced oxidative stress in leaves as indicated by an increase in thiobarbituric acid-reactive substances (TBARS) level and chlorophyll breakdown.
-
Zn supplementation, at low level, restored and enhanced the functional activity of these enzymes (SOD, CAT, APX and GR) as compared to Cd-alone-treated plants.
-
The beneficial effect of adequate Zn level on Cd toxicity was confirmed by a significant decrease in TBARS level and restoration of chlorophyll content. However, when Zn was added at a high level in combination with Cd, there was an accumulation of oxidative stress, which was higher than that for Cd or excess Zn alone treatments.
|
[80] |
| Positive response |
Drought + ozone |
Birch |
-
Combined stress increases the N concentration in the leaves, the thickness of the upper epidermal cell wall, the number of pectinaceous projections of mesophyll cell walls, and the vacuolar tannin-like depositions and phenolic droplets, which are regarded as signs of activated stress defense mechanisms.
-
The increase in specific foliage mass, cytoplasmic lipids (younger leaves), and a condensed appearance of the upper epidermal mucilaginous layer were caused by both drought and ozone but were not additive.
|
[81] |
| |
|
beech trees (Fagus sylvatica) |
-
Photosynthesis (Amax), stomatal conductance (gs), and electron transport rate (ETR) were lowered during drought rather than ozone, whereas chlorophyll levels did not differ.
-
Comparison of AOT40 [Accumulated Ozone exposure over a threshold of 40 ppb ((80 µg/m³)], an O3 exposure-based risk index of O3 stress, and cumulative ozone uptake (COU) yielded a linear relationship throughout humid growth conditions. The findings support the hypothesis that drought protects plants from O3 injury by stomatal closure, which restricts O3 influx into leaves and decouples COU from high external ozone levels.
-
High AOT40 erroneously suggested high O3risk under drought. Enhanced ozone levels did not aggravate drought effects in leaves and stem.
|
[82] |
| |
|
Medicago truncatula |
-
Medicago truncatula cultivar Jemalong that is sensitive to ozone and drought stress when applied singly, showed tolerance when subjected to a combined application of these stresses.
-
Lowered stomatal conductance may be a vital tolerance mechanism to overcome combined ozone and drought.
-
Sustained increases in both reduced ascorbate and glutathione in response to combined stress may play a role in lowering reactive oxygen species and nitric oxide toxicity.
-
Transcriptome analysis indicated that genes associated with glucan metabolism, responses to temperature and light signaling may play a role in dampening ozone responses due to drought-induced stomatal closure during combined occurrence of these two stresses.
-
Gene ontologies for jasmonic acid signaling and innate immunity were enriched among the 300 differentially expressed genes unique to combined stress.
-
Differential expression of transcription factors associated with redox, defense signaling, jasmonate responses and chromatin modifications may be important for evoking novel gene networks during combined occurrence of drought and ozone.
-
The alterations in redox milieu and distinct transcriptome changes in response to combined stress could aid in tweaking the metabolome and proteome to annul the detrimental effects of ozone and drought in Jemalong.
|
[83] |
| |
Drought + high CO2 |
Plants |
-
Elevated atmospheric CO2 cause an increase in leaf and canopy photosynthesis, especially in C3 plants, with minor changes in dark respiration. Additional CO2causes an increase in biomass without marked alteration in dry matter partitioning, reduced transpiration of most plants and improvement in WUE. However, spatiotemporal variation in these attributes impact agronomic performance and crop water use in a site-specific manner. Nutrient acquisition is closely associated with overall biomass and strongly influenced by root surface area.
|
[84] |
| |
salinity + High temperature |
tomato |
-
The combination of heat and salinity provides a significant level of protection to tomato plants from the effects of salinity. We observed a specific response of plants to the stress combination, which included accumulation of glycine betaine and trehalose. The accumulation of these compounds under the stress combination was linked to the maintenance of a high K+concentration and thus a lower Na+/K+ratio, with a better performance of the cell water status and photosynthesis as compared with salinity alone.
|
[14] |
| |
Salinity + hypoxia |
Salix |
|
[85] |
| |
Salinity + high CO2 |
lettuce |
-
Elevated CO2 and its combination with salinity or high light increases biomass production.
-
Elevated CO2 and its combination with salinity or high light increases the antioxidant capacity, while high light treatment alone increased the antioxidant capacity of red-leaf lettuce, but not of green-leaf lettuce.
|
[86] |
| |
Salinity + boron |
Zea mays |
-
Under salt stress, the activity of specific membrane components can be influenced directly by boron, regulating the water uptake and water transport through the functions of certain aquaporin isoforms.
|
[87] |
| |
Ozone + pathogen |
Plants |
|
[88] |
| |
|
Microbes |
-
The acetate, propionate, and butyrate buffered aqueous ozone combinations had a significant 3–4 log reduction of S. aureus (p < 0.05) colony forming unit (CFU), while citrate or oxalate buffered aqueous ozone, statistically significant versus buffer alone, had less activity.
|
[89] |
| |
Ozone + UV |
Escherichia coli |
|
[90] |
| |
Ozone + high CO2 |
Rice |
-
Elevated CO2 (627ppm) increases rice yields by 23%. Modest increases in grain mass and larger increases in panicle and grain number contributed to this response.
-
The response of rice to elevated CO2 varied with fumigation technique. The more closely the fumigation conditions mimicked field conditions, the smaller was the stimulation of yield by elevated CO2.
-
Free air concentration enrichment (FACE) experiments showed only a 12% increase in rice yield.
-
When compared with rice grown in charcoal-filtered air, rice exposed to 62° ppb O3 showed a 14% decrease in yield. Many determinants of yield, including photosynthesis, biomass, leaf area index, grain number and grain mass, were reduced by elevated O3.
|
[91] |
| |
Pathogen + UV |
Various plants |
|
[88] |
| |
High CO2 + high light |
lettuce |
-
High light treatment alone increased production in green-leaf lettuce but not in red-leaf lettuce. On the other hand, elevated CO2and its combination with salinity or high light increased the antioxidant capacity, while high light treatment alone increased the antioxidant capacity of red-leaf lettuce, but not of green-leaf lettuce.
|
[86] |