Heavy metals (HMs) are released into the environment by many human activities and persist in water even after remediation. The efficient filtration of solubilized HMs is extremely difficult. Phytoremediation appears a convenient tool to remove HMs from polluted water, but it is limited by the choice of plants able to adapt to filtration of polluted water in terms of space and physiological needs. Biomasses are often preferred. Aquatic moss biomasses, thanks to gametophyte characteristics, can act as live filtering material. The potential for phytoremediation of Hypnales aquatic mosses has been poorly investigated compared to aquatic macrophytes. Their potential is usually indicated as a tool for bioindication and environmental monitoring more than for pollutant removal. When phytoremediation has been considered, insufficient attention has been paid to the adaptability of biomasses to different needs. In this study the heavy metal uptake of moss Taxiphyllum barbieri grown in two different light conditions, was tested with high concentrations of elements such as Pb, Cd, Zn, Cu, As, and Cr. This moss produces dense mats with few culture needs. The experimental design confirmed the capacity of the moss to accumulate HMs accordingly to their physiology and then demonstrated that a significant proportion of HMs was accumulated within a few hours. In addition to the biosorption effect, an evident contribution of the active simplastic mass can be evidenced. These reports of HM accumulation within short time intervals, show how this moss is particularly suitable as an adaptable bio-filter, representing a new opportunity for water eco-sustainable remediation.
- moss
- phytofiltration
- phytoremediation
- heavy metals
- Hypnales
- Taxiphyllum barbieri
1. Introduction

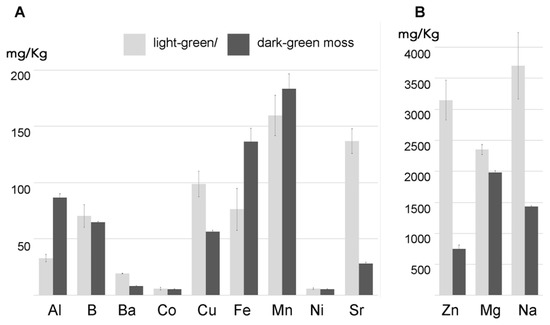
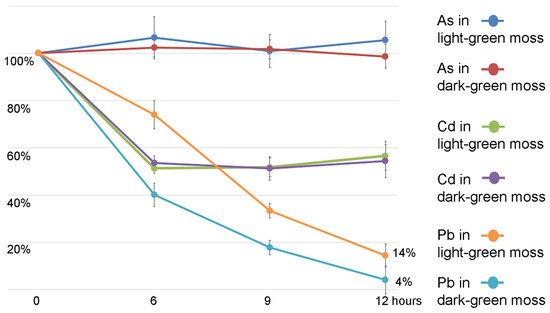
2. Mosess Grown in Different Conditions Have Different Uptake Capacities

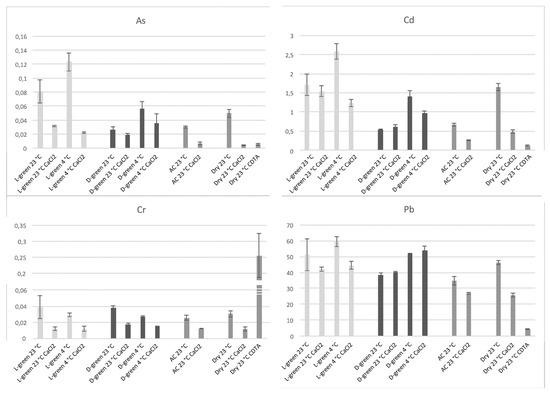
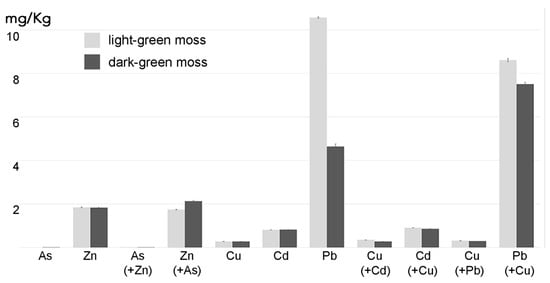
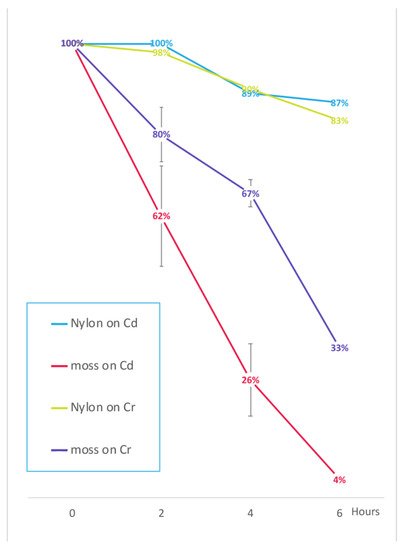
3. Influence of Pectins on HM Absorption
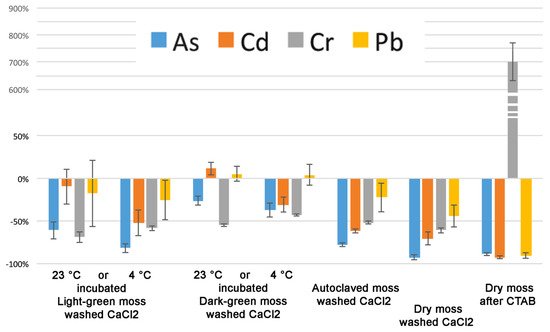
4. Discussion
This entry is adapted from the peer-reviewed paper 10.3390/ijms21134769
References
- Rahman, M.A.; Hasegawa, H. Aquatic arsenic: Phytoremediation using floating macrophytes. Chemosphere 2011, 83, 633–646.
- Diaz, S.; Villares, R.; Carballeira, A. Uptake Kinetics of As, Hg, Sb, and Se in the Aquatic Moss Fontinalis antipyretica Hedw. Water Air Soil Pollut. 2012, 223, 3409–3423.
- Esposito, S.; Sorbo, S.; Conte, B.; Basile, A. Effects of heavy metals on ultrastructure and HSP70S induction in the aquatic moss Leptodictyum riparium Hedw. Int. J. Phytoremediat. 2012, 14, 443–455.
- Esposito, S.; Loppi, S.; Monaci, F.; Paoli, L.; Vannini, A.; Sorbo, S.; Maresca, V.; Fusaro, L.; Karam, E.A.; Lentini, M.; et al. In-field and in-vitro study of the moss Leptodictyum riparium as bioindicator of toxic metal pollution in the aquatic environment: Ultrastructural damage, oxidative stress and HSP70 induction. PLoS ONE 2018, 13, e0195717.
- Koch, I.; Wang, L.; Ollson, C.A.; Cullen, W.R.; Reimer, K.J. The predominance of inorganic arsenic species in plants from Yellowknife, Northwest Territories, Canada. Environ. Sci. Technol. 2000, 34, 22–26.
- Sandhi, A.; Landberg, T.; Greger, M. Phytofiltration of arsenic by aquatic moss (Warnstorfia fluitans). Environ. Pollut. 2018, 237, 1098–1105.
- Itouga, M.; Hayatsu, M.; Sato, M.; Tsuboi, Y.; Kato, Y.; Toyooka, K.; Suzuki, S.; Nakatsuka, S.; Kawakami, S.; Kikuchi, J.; et al. Protonema of the moss Funaria hygrometrica can function as a lead (Pb) adsorbent. PLoS ONE 2017, 12, e0189726.
- Prasad, M.N.V.; De Oliveira Freitas, H.M. Metal hyperaccumulation in plants: Biodiversity prospecting for phytoremediation technology. Electron. J. Biotechnol. 2003, 6, 285–321.
- Shanab, S.; Essa, A.; Shalaby, E. Bioremoval capacity of three heavy metals by some microalgae species (Egyptian isolates). Plant Signal. Behav. 2012, 7, 392–399.
- Edelstein, M.; Ben-Hur, M. Heavy metals and metalloids: Sources, risks and strategies to reduce their accumulation in horticultural crops. Sci. Hortic. 2018, 234, 431–444.
- Srivastava, N.K.; Majumder, C.B. Novel biofiltration methods for the treatment of heavy metals from industrial wastewater. J. Hazard. Mater. 2008, 151, 1–8.
- Bolisetty, S.; Mezzenga, R. Amyloid-carbon hybrid membranes for universal water purification. Nat. Nanotechnol. 2016, 11, 365–371.
- Shaffer, D.L.; Tousley, M.E.; Elimelech, M. Influence of polyamide membrane surface chemistry on gypsum scaling behavior. J. Membr. Sci. 2017, 525, 249–256.
- Ye, C.C.; An, Q.F.; Wu, J.K.; Zhao, F.Y.; Zheng, P.Y.; Wang, N.X. Nanofiltration membranes consisting of quaternized polyelectrolyte complex nanoparticles for heavy metal removal. Eng. J. 2019, 359, 994–1005.
- Šuňovská, A.; Hasíková, V.; Horník, M.; Pipíška, M.; Hostin, S.; Lesný, J. Removal of Cd by dried biomass of freshwater moss Vesicularia dubyana: Batch and column studies. Desalin. Water Treat. 2015, 57, 2657–2668.
- Kadukova, J.; Štofko, M. Biosorption of heavy metal ions from aqueous solutions. In Environmental Research Trends; Cato, M.A., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2007; pp. 133–152. ISBN 978-1-60021-556-8.
- Sag, Y.; Kutsal, T. Determination of the biosorption activation energies of heavy metal ions on Zoogloea ramigera and Rhizopus arrhizus. Biochem. Eng. J. 2000, 6, 145–151.
- Dixit, S.; Singh, D.P. Phycoremediation: Future Perspective of Green Technology. In Algae and Environmental Sustainability; Singh, B., Bauddh, K., Bux, F., Eds.; Springer: New Delhi, India, 2015; pp. 9–21. ISBN 978 81-322 2641-3.
- Kayalvizhi, K.; Vijayaraghavan, K.; Velan, M. Biosorption of Cr (VI) using a novel microalga Rhizoclonium hookeri: Equilibrium, kinetics and thermodynamic studies. Desalin. Water Treat. 2014, 56, 194–203.
- Lombi, E.; Tearall, K.L.; Howarth, J.R.; Zhao, F.-J.; Hawkesford, M.J.; McGrath, S.P. Influence of Iron Status on Cadmium and Zinc Uptake by Different Ecotypes of the Hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2002, 128, 1359–1367.
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.S.P. Alleviation of cadmium toxicity in Brassica juncea L. (Czern. & Coss.) by calcium application involves various physiological and biochemical strategies. PLoS ONE 2015, 10, e0114571.
- Tian, S.; Lu, L.; Zhang, J.; Wang, K.; Brown, P.; He, Z.; Liang, J.; Yang, X. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress. Chemosphere 2011, 84, 63–69.
- Basic, N.; Salamin, N.; Keller, C.; Galland, N.; Besnard, G. Cadmium hyperaccumulation and genetic differentiation of Thlaspi caerulescens populations. Biochem. Syst. Ecol. 2006, 34, 667–677.
- Walker, D.J.; Bernal, M.P. The effects of copper and lead on growth and zinc accumulation of Thlaspi caerulescens J. and C. Presl: Implications for phytoremediation of contaminated soils. Water Air Soil Pollut. 2004, 151, 361–372.
- De Caroli, M.; Furini, A.; Dalcorso, G.; Rojas, M.; Di Sansebastiano, G.-P. Endomembrane Reorganization Induced by Heavy Metals. Plants 2020, 9, 482.
- Ma, L.Q.; Komar, K.M.; Tu, C.; Zhang, W.; Cai, Y.; Kennelley, E.D. A fern that hyperaccumulates arsenic. Nature 2001, 409, 579.
- Chen, G.; Liu, X.; Brookes, P.C.; Xu, J. Opportunities for Phytoremediation and Bioindication of Arsenic Contaminated Water Using a Submerged Aquatic Plant: Vallisneria natans (lour.) Hara. Int. J. Phytoremediat. 2015, 17, 249–255.
- Zhao, F.-J.; McGrath, S.P.; Meharg, A.A. Arsenic as a Food Chain Contaminant: Mechanisms of Plant Uptake and Metabolism and Mitigation Strategies. Annu. Rev. Plant Biol. 2010, 61, 535–559.
- Bergqvist, C.; Greger, M. Arsenic accumulation and speciation in plants from different habitats. Appl. Geochem. 2012, 27, 615–622.
- Nowak, J.; Frérot, H.; Faure, N.; Glorieux, C.; Liné, C.; Pourrut, B.; Pauwels, M. Can zinc pollution promote adaptive evolution in plants? Insights from a one-generation selection experiment. J. Exp. Bot. 2018, 69, 5561–5572.
- Zhang, X.; Hu, Y.; Liu, Y.; Chen, B. Arsenic uptake, accumulation and phytofiltration by duckweed (Spirodela polyrhiza L.). J. Environ. Sci. 2011, 23, 601–606.
- Carbonell-Barrachina, A.A.; Aarabi, M.A.; DeLaune, R.D.; Gambrell, R.P.; Patrick, W.H. The influence of arsenic chemical form and concentration on Spartina patens and Spartina alterniflora growth and tissue arsenic concentration. Plant Soil 1998, 198, 33–43.