Due to the selectivity in the function, specific distribution, and number of family members, GPCRs possess a potential therapeutic index and are important targets in different neurological conditions; some of the GPCRs listed in are therapeutic targets for cognition and memory deficits. The important roles of different GPCRs are discussed below.
3.1. 5-HT Receptors
The serotonergic neurotransmitters play a critical role in cognitive and behavioural functions. These receptors are distributed in different brain regions, including the cortex, amygdala, and hippocampus, and are associated with learning and memory [
71]. Their roles in cognition and memory have made them important drug targets in neurodegenerative disorders. Some evidence suggests that the use of agonists of the 5-HT subtype, such as 5-HT
2A/2C or 5-HT
4, can prevent memory impairment and improve learning ability. A similar effect can also be obtained by using an antagonist of 5-HT
1A or 5-HT
3 and the 5-HT
1B receptor [
72].
The 5-HT
6 receptor (5-HT
6R) has been shown to regulate several neurotransmitter pathways, including the serotonergic, cholinergic, glutamatergic, and GABAergic systems [
73]. In addition, several studies have established that this receptor is involved in learning and memory processes. These studies also claim that 5-HT
6R antagonists could improve cognitive functions; some agents are in the preclinical stage [
74,
75]. Many of these agents are associated with substantial improvements in different cognitive tasks and enhanced memory retention or formation in rodents [
76]. To date, at least three candidates have already reached Phase II/III clinical trials as novel therapeutic agents for the treatment of AD [
77]. Indeed, the serotonin receptor 5-HT
6R is an attractive drug target for reversing memory loss and learning disabilities associated with NDDs [
77,
78,
79]. A recent study has discovered a new benzimidazole-based compound that is an antagonist of 5-HT
6R and improves the novel object identification task in memory-deficit mice [
76].
Moreover, in a Phase I trial, PRX-07034, a selective 5-HT
6R antagonist, showed high selectivity for 5-HT
6Rs over other 5-HT receptors and nonserotonin receptors. This derivative at doses of 1 and 3 mg/kg enhanced short-term memory and improved cognitive flexibility in rats [
80]. Although Phase II trials have not yet been concluded [
81,
82], this drug candidate could be useful in treating dementia in AD [
83]. Another selective antagonist, AVN-322, which is a derivative of AVN-101 and AVN-211, is undergoing Phase I trials for AD and schizophrenia. This drug has been reported to reverse the detrimental cognitive effects of scopolamine and MK-80 [
84]. The AVN-101 is a multitarget serotonin antagonist that blocks 5-HT
7R and has a low, but potential, affinity towards 5-HT
6, 5-HT
2A, 5-HT
2C, and adrenergic receptors. It is also a dynamic candidate that is an antagonist of both 5-HT
6 and 5-HT
7. Both receptors follow the same signal transduction pathway, which is important for learning, memory, and anxiety. After completing Phase I trials, the drug is poised to start Phase II trials for AD and anxiety [
85]. In addition, the benefit of AVN-101 in treating AD progression is the symptomatic relief of anxiety, depression, sleep disorders, and associated mood disorders [
85]. Another derivative, AVN-211, has been reported to be a well-tolerated antagonist at a dose of 20 mg/kg of body weight and is suggested in AD therapy for its positive impact on cognition [
86].
Idalopirdine is an antagonist of 5-HT
6R that was developed initially for schizophrenia. Later analysis revealed its positive effect on cognition, and it was recommended for use in AD-associated dementia. Idalopirdine blocks 5-HT
6R and increases acetylcholine in the CNS. In particular, it increases acetylcholine by inhibiting CYP206, an enzyme that is involved in the metabolism of donepezil. Therefore, cotreatment with donepezil increases donepezil’s bioavailability [
87]. During a trial, 90 mg of idalopirdine taken daily with donepezil (10 mg) improved cognitive function significantly [
87]. An AChE inhibitor, donepezil, and an NMDAR inhibitor, memantine, were used in a clinical trial involving VaD patients and resulted in cognitive improvement [
88]. Monotherapy with intepirdine was found to be well tolerated, but its efficacy was lower than expected in AD. Further, the results of different studies were inconsistent. For instance, one study found a significant change in global function but not cognition [
89], while another report failed to find any significant improvements [
90,
91].
Latrepirdine, or Dimebon, is an antihistamine developed originally for allergic rhinitis. Later, the drug was found to have procognitive effects, which are the result of antagonising 5-HT
6R. Latrepirdine was also reported to stabilise mitochondria and works as a neuroprotective drug [
82]. Based on fact and figures, latrepirdine was recommended for clinical trials in both AD and HD in the 2000s [
82]. After completion of a small pilot study, the drug passed its Phase I and II clinical trials. However, in the Phase III trial, use of latrepirdine was terminated due to lack of efficacy [
82,
92]. SUVN-502 is another selective 5-HT
6R antagonist. After completing Phase I trials with no notable adverse effects and better tolerance [
93], the drug is currently in Phase II clinical trials involving patients with moderate AD [
82]. This drug has been reported to promote acetylcholine and glutamate in the CNS when administered in combination with donepezil [
94,
95]. Agomelatine, a 5-HT
2C antagonist, is another drug undergoing clinical trials that promises to improve prefrontal dopaminergic tone in FTD patients [
96]. One clinical trial involving agomelatine reported that it reduces depression in AD patients significantly and decreases depression and motor symptoms in PD patients [
97,
98]. These data suggest that the 5-HT
2C receptor could also be an important target in neurodegenerative disorder therapy.
Tryptophan (TRP) is an essential amino acid and serotonin precursor, and its depletion can disrupt serotonin synthesis in the brain. That depletion could also produce a paradigm of the serotonin deficit models, which has been demonstrated in different animals, including mice [
99], rats [
100], primates [
101], and humans [
102]. Several studies have assessed the relation between TRP depletion and cognitive function. For instance, Mendelsohn et al. [
103] used a diet lacking in TRP but enriched in large neutral amino acids to produce an almost 50% downregulation of 5-HT levels in the cortex, striatum, and hippocampus [
103]. That study, however, did not find any significant effect of TRP depletion on spatial, episodic, or working memory and instead reported semantic memory improvement even in the depleted state [
103].
Moreover, despite the modest effect of TRP depletion on sustained attention, such as vigilance, it does not significantly affect specific impaired executive functions such as planning, decision making, and responding [
103,
104]. To the contrary, tryptophan-rich supplementation of a regular diet can enhance serotonin synthesis, and a clinical study found that tryptophan supplementation improved reaction times, visual memory, and attention [
105]. Chronic ingestion of docosahexaenoic acid phospholipids with melatonin and tryptophan for 12 weeks also improves mild cognition impairment in elderly patients [
106].
Vortioxetine [
13], a serotonin transporter inhibitor (SERT), also works as an antagonist of 5-HT
3, 5-HT
7, and 5-HT
1D receptors, an agonist of 5-HT
1A, and a partial agonist of 5-HT
1B. A recent meta-analysis of a 6–8-week treatment with vortioxetine (5-20 mg/day) produced an incremental reduction in depression symptoms, and an increasing effect was associated with an increase in dose [
107]. Vortioxetine was also shown to improve cognitive performance in patients with acute major depressive disorder [
107]. Similarly, a study involving chronic citalopram use (20 mg; single dose daily) resulted in improved impulse response and contextual information processing abilities on a delayed nonmatching to sample task (DNMST) in healthy controls (n = 20). Acute administration (24 hr) of that drug showed no effect on working memory or impulsive responses. The authors of the study also suggested that DNMST makes a contribution to the activation of 5-HT
1A receptors in the entorhinal cortex and hippocampus [
108].
Chronic consumption of a diet high in saturated fats and low in fibre is associated with obesity and cognitive decline. In this context, dietary supplementation can prevent cognitive decline by altering serotoninergic signalling in the brain. A recent study with high-fat, low-fibre (5% dietary fibre)-induced obese models has demonstrated an association with the upregulation of 5-HT
1AR and 5-HT
2AR binding density in the rat brain in comparison to the low-fat diet group [
109]. With the inclusion of galacto-oligosaccharides and resistant starch, receptor binding densities in the hippocampal and hypothalamic region are reduced, improving cognitive function.
3.2. Dopamine Receptors
In the late 1950s, Carlsson identified dopamine as a potential neurotransmitter in the brain. Later, it was discovered that a progressive decrease in dopamine is associated with the pathophysiology of PD [
51,
110,
111]. This finding introduced levodopa, the metabolic precursor of dopamine, into the symptomatic treatment of PD. In the past few years, several investigations have been conducted on dopamine and cognitive function [
112,
113,
114,
115]. Although dopamine neurons are very few in number compared to the total neuronal population in the brain (< 1/100,000), they are involved in neuroendocrine regulation, mood, motivation, and psychological processes, including working memory and learning [
111,
112]. Although no specific mechanism has yet been confirmed, the inhibition of tyrosine hydroxylase activity and tyrosine conversion into dopamine and norepinephrine are involved in long-term memory formation [
116]. For example, vasopressin is a hypothalamic neuropeptide found to improve memory and does so by interacting with dopamine in the amygdala and serotonin of the hippocampus [
117].
As previously mentioned, some evidence suggests that dopamine modulates working memory, but its specific role is not yet fully defined, and its effect in memory processing has not been clarified. Haloperidol, a D2R antagonist, has recently been investigated for its effect on working memory improvement as well as for its distracting-information-ignoring capability. The study was designed with two testing sessions. In one session, participants took a placebo tablet, and in the other, they took Haloperidol 2.5 mg [
118]. The study showed that the deleterious effect of haloperidol on response conflict is associated with the negative effect of the drug on ignoring. The authors also suggest that D2R protects memory content from distraction through a general process, and inhibition of D2R could result in the impairment of response conflict as well as reduced quality of recall [
118]. D1R and D2R have been found to be important targets in FTD as well. Both DR antagonists (antipsychotics) and agonists (specifically, D2R) are used frequently in FTD treatment. Commonly used antipsychotics, such as olanzapine, quetiapine, and risperidone, have a high affinity for D2R and dissociate rapidly, resulting in very few side effects [
119]. A lack of presynaptic dopaminergic nerve terminal and postsynaptic D2R binding in the striatum is prevalent in FTD patients, and most of them complain of rigidity and bradykinesia. Therefore, FTD patients are currently being treated with the DR agonists carbidopa and levodopa to improve behavioural and psychotic symptoms.
However, SK609, a recently designed selective-small-molecule agonist of D3R, selectively inhibits norepinephrine reuptake as well as increases dopamine by 160% at an i.p. (intraperitoneal) dose of 4 mg/kg. SK609 improved rats’ performance in an attention task. Additionally, SK609 has been reported to improve cognitive function in low-performing rats. Interestingly, the molecule did not produce any side effects mediated by DA transporter (DAT) activity [
120], such as spontaneous locomotor activity.
3.3. Cannabinoid Receptors
Extensive evidence indicates that the endocannabinoid system has modulatory effects on cognitive function, and these effects have a substantial role in memory acquisition, consolidation, and extinction [
121]. However, the introduction of cannabinoid drugs often induces opposite effects, when used in anxiety, cognition, and other behavioural deficiencies, depending on the stress level and the aversiveness of the context [
121,
122]. It has been demonstrated that the use of an endocannabinoid receptor inhibitor under different environmental conditions has a substantial influence on cognitive function without affecting locomotor or emotional behaviour [
122]. Although it is difficult to define the exact role of the cannabinoid receptor, cannabinoid signalling influences memory processing. This influence has been demonstrated in several studies [
121,
122,
123].
In particular, Δ9-tetrahydrocannabinol (THC) is a cannabinoid receptor agonist that acts at CB1R to produce a wide range of biological and behavioural activity. This ligand’s association with cognitive deficits and poor decision making has been identified. Using Wistar rats in a rat gambling task study, it has been reported that a high dose of THC reduces premature responses, while another synthetic agonist produced the opposite reaction [
124]. Although acute or limited chronic use of THC does not affect subjects, long-term exposure can impair impulse control and attentional function [
125]. Therefore, chronic activation of CB1R or antagonism can impair or improve task performance, which makes it an interesting target. For example, in an animal study, the CB1 antagonist rimonabant produced an improvement in decision making [
124]. Meanwhile, cannabinoid receptor agonists, such as AEA (N-arachidonoyl ethanolamine) and noladin, have been reported to protect against Aβ-induced neurotoxicity in the human teratocarcinoma cell line NTERA-2/cl-D1. The effect was exerted through the CB1R- and mitogen-activated protein kinase (MAPK) pathway [
126]. Treatment with the CB1R antagonist SR141716A failed to protect against Aβ-induced amnesia [
127]. Another study with a triple transgenic mouse model of AD (3xTg-AD) reported that CB1R is up-regulated in the anterior thalamus at the age of 4 months, while the CB1R activity decreased gradually in the nucleus basalis of Meynert at 15 months of age [
128].
Activation of CB1R and CB2R was reported to affect the upregulation of PPARγ signalling in an animal model study [
129], where Aβ-induced neuroinflammation, neurodegeneration, and spatial memory impairment was attenuated. Activation of CB2R using a lower-dose agonist, JWH-015, eradicated native Aβ from human tissue and cleared a synthetic pathogenic Aβ peptide in a human macrophage cell line (THP-1). This effect was attributed to the antagonist SR144528, which is selective for CB2R, and the plaque removal effect induced by JWH-015 was reversed [
130]. A selective CB2R agonist 1-((3-benzyl-3-methyl-2,3-dihydro-1-benzofuran-6-yl) carbonyl) piperidine (MDA7) has shown a modulatory effect on cognitive impairment induced by bilateral microinjection of Aβ (1-40) fibrils into the hippocampal CA1 area of rats. Intraperitoneal treatment of MDA7 (15 mg/kg) for 14 days reduced microglial CD11b expression, promoted Aβ clearance, and restored synaptic plasticity, cognition, and memory [
131].
The increased expression of CB2R has been noted with reduced CB1R in late-onset HD, mainly in glial cells [
132]. The selective CB2R antagonist, SR144528, provides striatal neuron protection in HD rats; the mechanism includes glial cells [
133]. Genetic deletion of CB2R also exacerbates HD, but CB2R-selective agonists can reduce striatal neurodegeneration through microglial activation [
134]. SR141716 has also been reported to aggravate malonate-induced striatal pathology in HD rats [
135]. However, some studies report that cannabidiol, a phytoconstituent and an allosteric modulator of CB1R [
136], can rescue neuronal loss induced by THC by preventing THC-induced CB1R loss [
137]. Similarly, VCE-003.2, a cannabigerol derivative, improved the antioxidant barrier in 3-nitropropionic-acid-induced HD mice brains [
138]. This agent also showed neuroprotection in SOD1
G93A transgenic amyotrophic lateral sclerosis (ALS) mice by targeting CB2R and inhibiting endocannabinoid inactivation [
139]. Based on different reports, it is speculated that CBR antagonists may provide a better therapeutic insight into HD treatment.
3.4. Cholinergic Receptors
Cholinergic receptors are divided into two classes, including the muscarinic and nicotinic receptor families. These two families are further subclassed according to the occurrence of many ACh receptor subtypes, and their differential dendritic, somatic, axonal, and synaptic localisation contributes to the varied roles that these receptors play in the CNS [
140]. Five subtypes of mAChRs (M1 to M5) have been defined and pharmacologically characterized in the CNS. Their expression has been detected at very high levels in the subcortical structures and the cerebral cortex [
141]. A modest level of mAChR expression was also reported in the frontal cortex, parietal cortex, temporal cortex, entorhinal cortex, occipital cortex, and insular and cingulate cortexes, with the highest values recorded for the temporal and occipital cortexes [
141]. M1 receptors are the most abundantly expressed mAChRs [
141].
Nicotinic AChRs (nAChRs) constitute the second cholinergic receptor type. This type of receptor relies on ligand-gated ion channels and contains five subunits, which are assembled into homomeric or heteromeric subunit combinations. The pharmacological and biological characteristics of these receptors are determined by this combination [
142]. The nAChR subunits are composed of α4, α6, α7, β2, and β3 subunits and are mostly expressed in the striatum [
143,
144]. They are expressed on glutamatergic and dopaminergic neuron terminals, GABAergic interneurons, and cholinergic interneurons (ChIs) but are not present in medium spiny neurons (MSNs) [
145,
146]. However, nAChRs can express in both pre- and post-synaptic neurons. Thus, they can depolarise and increase excitability, causing glutamate, DA, and GABA to release [
147,
148].
The nAChRs form good targets in the treatment of neurodegeneration. In recent years, several agonists and partial agonists of the α7 subunit have been evaluated in the treatment of NDDs. Daily nicotine administration in a PD animal model produced improvements in motor coordination. That treatment also showed a beneficial effect on neuronal survival, as well as on microglial and astrocytic activation, providing neuroprotection against MPTP/MPP
+ toxicity [
149]. DMXBA (GTS-21) and ABT-107, both α7nAChR agonists, have also produced beneficial effects by attenuating nigrostriatal damage in 6-hydroxydopamine (6-OHDA)-induced rats [
51]. Another agonist, PHA 543613, attenuates early-stage HD induced by striatal quinolinic acid lesions. PHA 543613 reduced microglial activation to protect neurons [
150]. In another study, α7nAChR-agonist-treated 3xTg-AD mice showed improved cognitive function [
151]. However, PNU-282987 has also produced improved motor activity, anxiety, and learning and memory development in the B6C3-Tg mice model for AD [
152]. Similarly, two other agonists, AR-R17779 and ABBF (N-[(3R)-1-azabicyclo [2.2.2]oct-3-yl]-7-[2-(methoxy) phenyl]-1-benzofuran-2-carboxamide), are also reported to improve learning and memory [
153,
154].
ANAVEX2-73, a σ1R (sigma 1 receptor) agonist and muscarinic receptor ligand, has been studied on the Aβ25-35-injected AD mice model [
155]. Because Aβ25-35 injection modulates mitochondrial respiration in the hippocampus, ANAVEX2-73 (0.01-1 mg/kg IP) restored respiration to normal and prevented Aβ25-35-induced increases in lipid peroxidation levels, Bax/Bcl-2 ratio, and cytochrome c release into the cytosol [
155].
3.5. Metabotropic Glutamate Receptors
Metabotropic glutamate receptors (mGluRs) are members of the C GPCR family, and they consist of eight subtypes (mGlu1 to mGlu8) that are further subdivided into three groups depending on their amino acid sequences, G-protein coupling, and pharmacological characteristics. Group I consists of mGlu1 and mGlu5 receptors, which are coupled to G
q/G
11 [
156]. The Group II (mGlu2, mGlu3) and Group III (mGlu4, mGlu6, mGlu7, mGlu8) subtypes are coupled to G
i/G
o. Both of these groups regulate adenylate cyclase negatively and can also activate the MAP kinase and PI-3-kinase pathways [
156,
157]. However, overexpression of mGlu5 has been reported in different neurodegenerative disorders [
157]. Specifically, Aβ plaques have been found in the surroundings of astrocytes, spinal cord lesions, and MS lesions, ALS, PD, and in the hippocampal astrocytes of Down syndrome patients [
158,
159].
Several mGluRs agonists, antagonists, and positive and negative allosteric modulators have been studied using different neurodegenerative animal models. Collectively, these data on mGluRs provide an insight into the development of therapeutics for treating NDDs. However, LY341495 (an antagonist to Group I/II mGluR) was reported as blocking Aβ-enhanced long-term depression and improving synaptic plasticity [
160]. The same study also reported that pretreatment with an mGluR1/5 agonist, 3,5-dihydroxyphenylglycine (DHPG), decreased Aβ-enhanced long-term depression [
160]. SIB1757, a noncompetitive antagonist of mGluR5, prevented Aβ oligomer-induced synaptic N-Methyl-D-aspartic acid receptor NMDAR reduction [
161]. A comparison study targeting Group II mGluRs showed that an mGlu2R positive allosteric modulator, N-4′-cyano-biphenyl-3-yl)-N-(3-pyridinylmethyl)-ethanesulfonamide hydrochloride (LY566332), amplified Aβ-induced neurodegeneration [
162]. Treatment with the antagonist (2S,1′S,2′S)-2-(9-xanthylmethyl)-2-(2′-carboxycyclopropyl) glycine (LY341495) of mGlu2/3R prevented this effect. Similarly, the dual mGlu2/3 receptor agonist (−)-2-oxa-4-aminobicyclo [3.1.0] exhane-4,6-dicarboxylic acid (LY379268) exhibited neuroprotection via a paracrine mechanism mediated by transforming growth factor-β1 [
162]. Therefore, dual activation of mGlu2R and mGlu3R may be a prime target for providing neuroprotection against Aβ-induced toxicity, and negative modulation of mGlu5R would also be a good target for PD and AD treatment.
3.6. Orphan GPCRs
More than 140 GPCRs remain mysterious and are referred to as ‘orphans’ (oGPCRs); most of them do not have any known ligands. Although very little is known about their endogenous or exogenous ligands, these so-called oGPCRS have gained considerable attention as drug targets [
163]. Several findings have postulated the critical role of oGPCRs in the cognitive deficits in disorders such as AD and schizophrenia [
164,
165,
166,
167]. For example, an expression map of the mouse brain has listed 78 oGPCRs and showed that many of them are relevant to cognition, motivation, and emotional processing [
168]. That study reported that oGPCRs, such as GPR17, GPR27, GPR37, GPR39, GPR63, GPR85, GPR88, GPR123 (Adgra1), GPR125 (Adgra3), GPR153, GPR176, and GPRc5c, are highly expressed in the prefrontal cortex (PFC) region of the mouse brain, which is involved in cognition and learning. Both mouse and human brain analyses have shown that four oGPCRs are highly expressed, including GPR88, GPR123, GPR149, and GPR151, but information about these oGPCRs is minimal, with the exception of GPR88 [
168].
However, GPR3 has exhibited stable and enhanced expression throughout the ageing process in ten different regions of the healthy human brain, which has been linked to AD pathogenesis in multiple cohort studies [
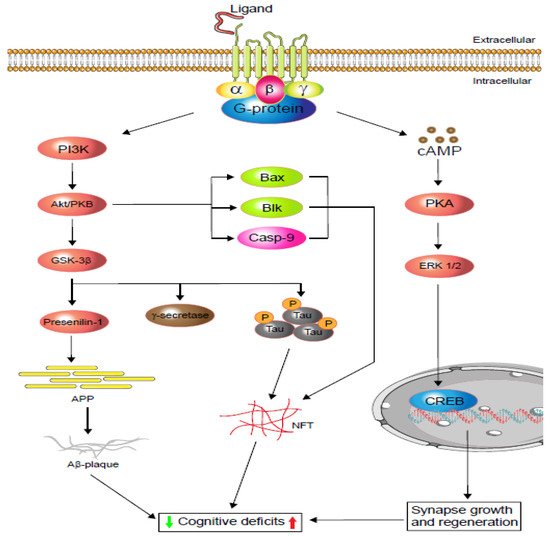
166]. Additionally, GPR3, independent of G-protein coupling, recruits β-arrestin 2 and promotes γ-secretase activity, which increases Aβ precursor protein (APP) cleavage and accelerates Aβ peptide production and accumulation [
169]. Thus, deletion of GPR3 in the AD mouse alleviated cognitive impairment and restored memory [
166]. Although GPR158 improves memory through transducing osteocalcin and regulating IP3 and BDNF in the CA3 neurons [
167], GPR158 overexpression in PFC has a potential role in depressive behaviour, which is reversed by its depletion [
170].
GPR52 is a nonodorant GPCR and is colocalized with both D1 and D2 receptors in the basal ganglia neurons. Histological experiments suggest that GPR52 promotes and regulates the
Cre-lox system and may also modulate cognition and emotion, as it has involvement with both dopaminergic and glutaminergic neurotransmission [
171]. Thus, the incorporation of the GPR52 antagonist may potentiate cognitive improvement and exert anxiolytic activity in psychiatric disorders [
171]. GPR3 knockout also produced anxiety and depressive behaviour, with no noticeable locomotor impairment under stressful conditions. However, the lack of GPR3 has no preventive action in the learning involved in fear memory in a similar stressful condition in mice [
172]. GPR3 also regulates serotonin (5-HT) and dopamine (DA) synthesis and reuptake, which makes it a primary target as well. A study has reported the possibility that serotonin reduction in the frontal cortex and hippocampus causes aggressive behaviours in GPR3 knockout mice [
172]. This finding indicates that GPR3 modulates the serotonergic and dopaminergic system, which makes it a potential target in the therapy of AD or schizophrenia.
GPR55 is highly expressed in the pyramidal cells in the hippocampal CA1 and CA3 layers and modulates the synaptic plasticity of pyramidal cells [
173]. However, GPR85 is highly expressed in the dentate gyrus region of the hippocampus [
174,
175] and prominently expresses in the phases of neuronal differentiation in the developing cerebral cortex [
176]. This expression suggests a possible role of GPR85 in cognition, and this receptor could become a potential drug target as well.