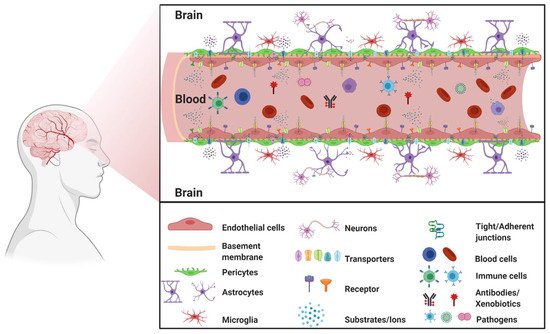
The blood–brain barrier (BBB) is a complex cellular barrier composed of a tightly sealed monolayer of specialized brain capillary endothelial cells lined on a basement membrane and surrounded by adjacent perivascular astrocytes, pericytes, and microglia. The major function of the BBB is to separate the circulating blood from the brain extracellular fluid and maintain homeostasis in the central nervous system (CNS) ().
1.1. BBB Facts and Figures
For an average adult, the human brain represents 2% of total body mass. Remarkably, despite this relatively small contribution to total body weight, the brain is composed of ~170 billion neuronal, glial, or non-neuronal cells [
1]. Under resting conditions, about 20% of total blood flow from the heart is received by the brain in healthy adults [
2,
3]. The BBB has a large surface area of almost 15 to 25 m
2, a total length of capillaries of ~400 miles, microcapillaries with a diameter of 7–10 µm, and an average inter-capillary distance of ~40 µm [
1,
4,
5,
6]. These characteristics allow very selective diffusion and penetration of compounds [
1,
4]. The large BBB surface area and its critical role in maintaining homeostasis of the brain require regulation and support from accompanying brain cells, which all together form the neurovascular unit (NVU).
1.2. The Neurovascular Unit
The NVU is a relatively new concept in neuroscience. The NVU was originally described in 1996 as a three-party unit composed of neurons, cerebral blood vessels, and astrocytes, collectively regulating cerebral blood flow [
7,
8]. In 2001, during the Stroke Progress Review Group meeting of the National Institute of Neurological Disorders and Stroke, endothelial cells were recognized as the NVU blood vessel component to emphasizes the unique relationship between brain cells and cerebral vasculature [
9,
10]. Since then, the NVU has gained much interest from the neuroscience community and provided a better understanding of the inter-play and signaling between these different cell types. The gate function of the BBB is mainly due to the tightly sealed monolayer of endothelial cells lining the brain capillaries but the interaction of these endothelial cells with surrounding neurons, astrocytes, microglia, and pericytes is key to maintaining proper integrity and function of the BBB [
11]. Furthermore, the NVU has become the center of attention for our understanding of the pathology of several neurodegenerative diseases and, consequently, may represent a potential therapeutic target [
12].
1.3. Movement across the BBB
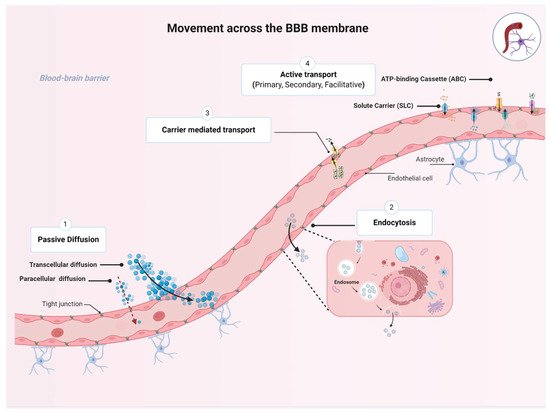
The BBB plays an essential role in maintaining proper CNS functions by selectively regulating movement of essential nutrients and toxins into and out of the CNS. While several molecules are restricted from entering the brain, many molecules can move across the BBB membrane using four basic mechanisms, namely, passive diffusion, endocytosis, and carrier-mediated transport or active transport, depending on the characteristics of the molecule and the direction of the transport () [
13].
Figure 2. Transport pathways across the blood–brain barrier (BBB). The passage of various molecules through the brain implies four basic mechanisms allowing specific molecules to move across the BBB membrane including: (1) the passive diffusion (spontaneous movement across a concentration gradient), (2) endocytosis (receptor-mediated, adsorptive, or bulk-phase endocytosis), (3) carrier-mediated transport (movement across a concentration gradient and energy independent), and (4) active transport (movement of molecules against a concentration gradient and energy dependent). Collectively, the four mechanism plays an essential role for maintaining brain homeostasis. (Created with BioRender, accessed on 27 January 2021).
Passive diffusion is the movement across a concentration gradient from high to low concentrations, without expenditure of biological energy or involvement of a carrier protein [
14]. Passive diffusion can be divided into paracellular (i.e., between cells), transcellular (i.e., across cells), or via aqueous channels traversing the BBB lipid bilayer. Transcellular diffusion of compounds into the brain is dependent on their lipophilicity, as assessed by oil/water partition coefficients; the more lipophilic a compound is, the easier it could be to cross the BBB. For instance, such physicochemical characteristics have been used to facilitate CNS distribution of antidepressants and antipsychotics by inserting halogenic elements (chlorine, fluorine, bromine) in their chemical structure [
15]. Transcellular diffusion is non-saturable and non-competitive [
16]. Examples of drugs that can passively diffuse across biological membranes include steroids, opioids (e.g., fentanyl, methadone), benzodiazepines, and first-generation antihistamines such as diphenhydramine [
14]. Small hydrophilic molecules such as nitrate, urea, glycerol, and arsenite could enter the CNS through aqueous channels [
17]. Finally, due to the tight junctions connecting endothelial cells of the BBB, paracellular diffusion does not occur to any great extent under normal physiological conditions [
6].
The second mechanism is endocytosis, which is defined as the vesicular transport across the BBB through receptor-mediated, adsorptive, or bulk-phase endocytosis [
14,
18]. Receptor-mediated endocytosis involves internalization of the substrate–receptor complex into the intracellular compartment where dissociation happens. This process can happen at both the luminal (apical) and the abluminal (basolateral) cell membranes. Transport of insulin and transferrin (iron transport protein) are classic examples of receptor-mediated endocytosis [
19,
20]. This transport mechanism is also energy-independent.
The third mechanism is carrier-mediated transport, through which the solute molecules move from high to low concentrations across the BBB. This membrane-bound protein-mediated transport system is an energy-independent system that could be uniporter (allows transportation of one solute at a time), symporter (carries two different solutes at the same time; the transported solute and the cotransported solute move in the same direction), or antiporter (also carries two different solutes; the transported solute and the cotransported solute move in opposite directions). This type of transporter is of utmost importance for CNS access to essential nutrients such as glucose through the glucose transporter 1, GLUT1 (SLC2A1) [
14]. Centrally acting drugs may also utilize this pathway; for example, L-Dopa and gabapentin are transported through the sodium-independent large neutral amino acid transporter (LAT1 or SLC7A5) [
21].
The fourth mechanism is active transport across the BBB through a carrier protein. This process is energy-dependent and often coupled with ATP hydrolysis; it enables the movement of substances against their concentration gradient [
22]. There are several energy-dependent transporters expressed at the BBB endothelium that work to transport essential micronutrients, ions, and some endogenous compounds into the CNS. Active transporters could favor influx but predominantly facilitate efflux of drugs from the CNS and therefore work to regulate the entry of potentially toxic substances. This BBB crossing process is the target of many therapeutic drugs that are transported into or out of the CNS, such as antineoplastic agents, opioid analgesic drugs, HIV-1 protease inhibitors, and antibiotics [
14].
While the four mechanisms are essential for maintaining brain homeostasis, the transport proteins expressed on the luminal (blood-facing) and abluminal surface of the BBB magnify the protection provided and strictly regulates the transcellular transport. Unfortunately, this process is affected by aging and age-associated disorders [
23]. Both the expression level and the functionality of transport proteins go through a transformation with aging under both physiological and pathological conditions. Therefore, this review focuses on the major transporters located at the BBB and sheds light on the influence of inflammatory conditions and diseases, such as Alzheimer’s disease (AD), epilepsy, and stroke, on the expression and functionality of the BBB drug transporters, the consequential modulation of drug distribution to the brain, and its impact on drug efficacy and toxicity.
2. Transporters Expressed at the BBB
The gatekeepers of the BBB are the endothelial cells lining its vasculature that express a variety of transporters that strictly regulate the influx and efflux of endogenous nutrients and exogenous toxins including xenobiotics and drugs [
24]. Below, an overview of the major superfamilies of transporters expressed at the BBB is provided. This includes the ATP-binding cassette (ABC) transporters and the solute carrier superfamily (SLC).
2.1. ATP-Binding Cassette (ABC) Transporters
The ATP-binding cassette (ABC) superfamily of transporters is perhaps the most studied family of active transporters that exists in nearly all living organisms [
14,
25]. The ABC family in humans includes 49 transporters grouped into seven sub-families, designated from ABCA to ABCG. Every transporter, on the basis of unique genetic and amino acid sequences, is assigned a number following the subfamily letter, e.g., ABCG2 [
26].
ABC transporters are ATP-driven transmembrane proteins that function as unidirectional pumps across the BBB for various compounds including amino acids, steroids, phospholipids, ions, polysaccharides, drugs, and xenobiotic compounds [
27]. At the BBB level, most ABC transporters act as efflux transporters (moving substrates outside of the CNS) and provide the BBB with a great clearance and defensive machinery [
28]. Many ABC transporters are well-known mediators of a multidrug resistance (MDR) phenotype described as the simultaneous resistance to multiple structurally unrelated compounds that are not due to an independent genetic mutation that confers resistance to a single xenobiotic [
14]. The most studied members of the ABC superfamily expressed at the BBB will be discussed below and include ABCB1 (P-glycoprotein, P-gp encoded by the gene
MDR1), ABCG2 (breast cancer resistance protein, BCRP), and ABCCs (multidrug-associated resistant proteins, MRPs); they are all efflux transporters, limiting therapeutic drug entry into the brain and preventing successful response to several agents intended to treat neurological disorders. Moreover, decreased expression or function of these transporters has been described in several CNS disorders [
29].
2.1.1. ABCB1
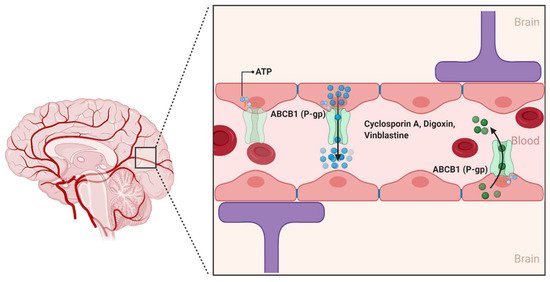
ABCB1 is the classical multidrug efflux transporter that was first described in 1974 by Dr. Victor Ling at the University of Toronto through the characterization of colchicine-resistant Chinese hamster ovary cells [
30,
31]. Since its discovery, ABCB1 has been characterized as a 170 kDa transmembrane protein that consists of two homologous halves, each with an intracellular ATP-binding site. In the human brain, ABCB1 is extensively expressed on the luminal side of the brain endothelial cells lining the BBB. ABCB1 limit the entry of a huge variety of hydrophobic amphipathic drugs, such as cyclosporin A, digoxin, and vinblastine into the CNS by pumping them from the endothelial cells back into the blood () [
32]. Unlike conventional transporters, which are the target of specific substrates, ABCB1 recognizes a wide variety of substrates that differ considerably in molecular size and structure [
31]. This includes several antineoplastic agents, calcium channel blockers, anti-depressants, anti-epileptic agents, and several HIV-1 protease inhibitors [
33].
Figure 3. The ABCB1 transporter at the blood–brain interfaces. In the human brain, the ABCB1 transporter is found on the luminal membrane side and restricts the penetration of compounds in the brain by limiting the uptake of several substrate drugs. (Created with BioRender, accessed on 27 January 2021).
Modulation of ABCB1 expression at the BBB has been described under various conditions [
29]. These include upregulation of ABCB1 when exposed to certain drugs or hormones in vitro. In addition, a number of in vivo studies suggest that ABCB1 expression is elevated in case of epilepsy and neurodegenerative disorders such as amyotrophic lateral sclerosis (ALS) [
34,
35]. An increase in ABCB1 expression was reported to induce progressive pharmacoresistance to riluzole, the only FDA-approved drug thus far for ALS [
36]. In contrast to ALS, downregulation of ABCB1 expression and function has been observed with aging and in patients with Alzheimer’s disease (AD) [
37]. Several studies also suggested that ABCB1 function is associated with AD progression, as ABCB1 plays an important role in the clearance of amyloid-β (Aβ), the hallmark of AD [
37,
38]. Activity level of ABCB1 is modulated by mutations in
ABCB1, and many recent reports have shown an association between
ABCB1 mutations and AD [
39]. Some of these variants are the
3435C > T, the
2677G > T/A and the
1236C > T polymorphisms [
39]. While some studies have shown no association between the
3435C > T polymorphism and AD risk, Van Assema et al. showed that the
1236C > T polymorphism may contribute to increased Aβ deposition in the brain [
39].
2.1.2. ABCG2
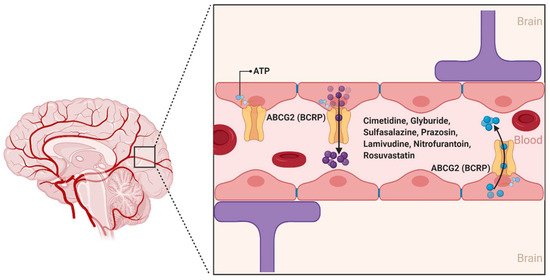
ABCG2 was first described in 1990 by Chen et al., and was further characterized in 1998 by Doyle et al. as a polytopic plasma membrane transporter protein of 75 kDa. Doyle et al. demonstrated using the human breast carcinoma cell line MCF-7/AdrVp—which does not express the efflux transporters ABCB1 or ABCC1—the multidrug resistance aspects of ABCG2 for substrates such as the anthracycline doxorubicin () [
40,
41]. While there are at least five members of the ABCG subfamily identified in humans, ABCG2 is the primary form involved in the transport of substances across the BBB [
42].
Figure 4. The ABCG2 transporter at the blood–brain interfaces. ABCG2 was originally identified as the breast cancer resistance protein (BCRP). In the human brain, ABCG2 is expressed on the luminal membrane side and plays an important role in limiting exposure of drug molecules to the CNS. (Created with BioRender, accessed on 27 January 2021).
ABCG2 is expressed on the luminal side of BBB capillary endothelial cells and acts as an ATP hydrolysis-dependent efflux transporter of a wide range of structurally and chemically distinct compounds, including physiological substrates such as sulfate and glucuronide conjugates of estrone and dehydroepiandrosterone [
42]. In addition, several antineoplastic agents, namely, daunorubicin, doxorubicin, imatinib, methotrexate, mitoxantrone, and topotecan are substrates of ABCG2 [
42,
43]. Other ABCG2 substrates from various drug classes include camptothecin derivatives, cimetidine, glyburide, sulfasalazine, prazosin, pantoprazole, methotrexate, lamivudine, nitrofurantoin, and rosuvastatin [
44]. A number of photosensitizers including pheophorbide A, protoporphyrin IX, and associated compounds are also substrates of ABCG2, suggesting a potential association between ABCG2 expression and cellular resistance to photodynamic therapy [
45].
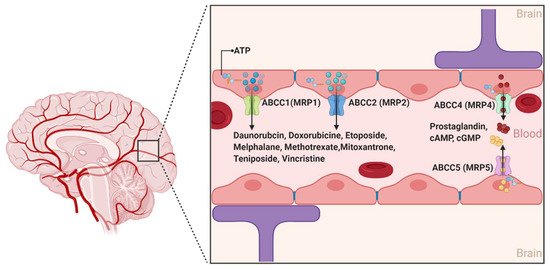
2.1.3. ABCCs
The ABCCs are ~190 kDa proteins whose physiological functions include organic anions and nucleotide-based analogs transport, signal transduction, and toxin secretion [
14]. Although at least 12 ABCC members have been identified to data, only 9 members (ABCC1-9) are involved in the efflux transport of drugs impacting their pharmacokinetics [
46]. While several ABCC members have long been associated with drug resistance across different organs of the human body, ABCC1, 2, 4, and 5 are the only isoforms with confirmed expression on the luminal side of the BBB endothelial cells. Moreover, the expression of ABCC1 and ABCC4 was also described at the basolateral side of the BBB () [
47]. Conflicting results have been reported for the expression of ABCC3 and ABCC6 [
48,
49,
50,
51]. Collectively, the localization of the aforementioned four different ABCC isoforms at the BBB suggests that the ABCCs play an important role in regulating the efflux of drugs from the brain back to the blood and restricting the influx of several compounds into the brain parenchyma [
52].
Figure 5. The ABCC transporters at the blood–brain interfaces. In the human brain, ABCC transporters are preferentially expressed on the luminal sides. ABCC proteins are active efflux transporters playing a role in the extrusion of several molecules. (Created with BioRender, accessed on 27 January 2021).
In contrast to ABCB1 and ABCG2, ABCC’s substrate structural profile is relatively restrictive [
14]. Notably, the ABCC’s isoforms generally transport unconjugated anionic drugs, organic anions, and their conjugated metabolites, such as glutathione conjugates (leukotriene C4 or LTC4), glucuronides (estradiol-17-
β-glucuronide), and glutathione disulfide (GSSG) [
53]. The fact that at least nine isoforms of ABCCs show drug efflux functions with substantial overlapping substrate affinity makes it difficult to differentiate each isoforms’ specific characteristics and function [
27]. This includes their potential role in the pathophysiology of several neurological diseases and response to treatment [
49,
54]. ABCC1 and ABCC4 exhibit a high affinity for the endogenous inflammatory mediator LTC4, suggesting a role in the transport of this compound across the BBB and a possible contribution to immune responses. In vitro studies have also shown that ABCC4 is a unique efflux transporter of prostaglandin E1 (PGE1) and PGE2 and that their transport was inhibited by nonsteroidal anti-inflammatory drugs such as indomethacin [
55]. In addition, a number of antineoplastic purine nucleotide anion analogs are effluxed by ABCC4, specifically the active metabolites of 6-mercaptopurine and 6-thioguanine, and bis(pivaloyloxymethyl)-9-[2-(phosphonomethoxy)ethyl]-adenine (PMEA) [
27]. ABCC4 and ABCC5 are the primary active transporters of endogenous nucleotides such as cAMP and cGMP, and may play a role in regulating their levels in the brain [
56].
2.2. The Solute Carrier (SLCs) Superfamily
The SLC superfamily of transporters represents the largest studied superfamily of secondary active and facilitative transporters [
57]. The SLC superfamily consists of almost 395 unique transporter genes divided into 65 sub-families (SLC 1-65) [
58]. Among the 52 identified SLC families, the SLCO family (which includes the Organic Anion Transporting Polypeptide subfamily (OATP)), SLC1A (high-affinity glutamate transporters, EAATs), SLC2A (glucose transporters, GLUTs), SLC7A, SLC15A, SLC16A, SLC21A, SLC22A (which includes the organic anion transporters (OATs) and organic cation transporters (OCTs)), SLC28A, and SLC29A families have been identified at the BBB [
59]. Unlike the ABC transporters, SLCs do not need ATP to transport substrates across biological membranes. Instead, the transport machinery is powered by an electrochemical gradient (e.g., Na
+) or a concentration gradient, depending on the solute being transported [
60]. These transporters are responsible for the transport of a wide range of substrates including organic cations and anions, peptides, monocarboxylates, steroids, signaling molecules, drugs, and drug conjugates [
59].
SLC transporters are expressed on both the luminal and abluminal sides of the BBB and, like other transporters, contribute to keeping the brain protected from toxic substances while extracting essential components from the blood [
61]. Although transporters in this family are capable of bidirectional transport, generally SLC transporters are considered to favor cellular uptake of drugs rather than their efflux [
59]. In addition, SLC transporters can stimulate the synthesis of acetylcholine, reduce oxidative stress, act upon carnitine, and prevent neurodegeneration [
62]. As such, these transporters are involved in the pathogenesis of several neurological disorders [
63].
2.2.1. Organic Anion Transporting Polypeptides (SLCOs and Formally OATPs)
The organic anion transporting polypeptides (SLCOs and formally OATPs) are a group of multi-specific sodium-independent membrane solute carrier transporters classified within the SLC21 family. The human SLCOs consists of 11 members, localized in several organs such as the kidney and the liver, and mediating the transport of a wide range of amphipathic substrates, including bile acids, steroid conjugates, thyroid hormones, and anionic peptides, in addition to several drugs and xenobiotics such as 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase inhibitors (i.e., statins), angiotensin receptor blockers, angiotensin-converting enzyme inhibitors, antibiotics, and other chemotherapeutic agents [
63,
64,
65]. While the expression and function of the Slcos in the BBB of rodents has been well documented, the expression and function of SLCOs at the human BBB is not clearly understood [
14]. Specifically, the expression of organic anion transporting polypeptide 1a4 (Slco1a4), Slco2a1, and Slco1c1 has been reported in capillary endothelial cells, capillary-enriched microvessels, and whole-brain microvessels [
66]. It has also been suggested that Slco1a4, a rodent ortholog of SLCO1A2, is the primary drug transporting Slco isoform expressed at the rat BBB [
67]. Recent studies using Slco1a4 (Oatp1a4) knockout mice also provide evidence for the Slco-mediated uptake of statins as knockout animals showed a reduced brain uptake of statins compared to age-matched wild-type control mice [
68]. Thompson et al. showed that the function expression of Slco1a4 at the rat BBB was significantly increased after induction of inflammation and pain, which was also reflected by the increased atorvastatin brain delivery via Slco1a4 [
66]. Animal studies also showed that Slco1c1 is primarily responsible for the uptake of thyroxine and conjugated sterols at the BBB, while Slco1a2 is a key player in mediating the CNS prostaglandin homeostasis [
67].
In vitro data using human brain endothelial cells suggested that SLCO1A2 (OATP1A2) plays an important role in drug delivery across the BBB (including removal of organic metabolites). Expression of SLCO1A2 was the first SLCO identified in human brain endothelial cells [
31,
59]. Moreover, using paraffin-embedded sections of human brain endothelial cells, Gao et al. showed that SLCO2B1 (OATP2B1) is abundantly expressed at the brain capillary endothelial cells and may be involved in mediating the transport of several neuropeptides and neurosteroids across the BBB [
69]. Additionally, immunohistochemical localization studies showed that SLCO1A2 is expressed on the luminal side of the BBB while Slco1A4 (rodent ortholog of human SLCO1A2) is expressed on both luminal and abluminal sides [
27,
70]. Additional studies are required to further understand the important role of SLCOs at the human BBB.
2.2.2. Organic Anion Transporters (SLC22s Formally OATs)
The organic anion transporters (OATs)—which include SLC22A6-8 (OAT1-3), SLC22A9 (OAT7), SLC22A10 (OAT5), SLC22A11 (OAT4), the human urate transporter 1 (URAT1, also known as SLC22A12), SLC22A13 (OAT10), and SLC22A20 (OAT6)—are another important multi-specific family of transporters that belong to the SLC22 superfamily [
71]. Similar to SLCOs, OATs mediate the transport of a broad range of chemically distinct endogenous and exogenous substrates such as hormones, prostaglandins, urate, dicarboxylates, and anionic neurotransmitter metabolites, as well as drugs and their associated metabolites [
72].
Human SLC22A6, SLC22A7, SLC22A8, SLC22A11, SLCA12, and SLC22A13 are predominantly expressed in the human kidneys; SLC22A7, SLC22A9, and SLC22A10 are expressed in the liver; SLC22A6 and SLC22A11 are expressed in the placenta; SLC22A6, SLC22A8, SLC22A11, and SLC22A13 are expressed in the brain; and SLCA22A20 is expressed in the olfactory mucosa [
70,
73,
74]. Proteomic analysis of human brain endothelial cells revealed differential expression of the SLC22s at the BBB level: SLC22A11 is known to be expressed at the BBB but its exact location is unclear; SLC22A6 and SLC22A8 are either not detected or have levels below the limit of quantification [
60,
75]. On the basis of animal studies, research found that Slc22a8 (Oat3) is expressed on the basolateral side of BBB [
76]. Western blot analysis of brain capillary-enriched fraction revealed higher Slc22a8 expression with greater molecular mass compared to kidney, suggesting that its major function is to clear substrates from the brain [
76]. Functional expression studies using mammalian cells showed that Slc22a8 has a broad range of substrate specificities including amphipathic organic anions, such as 17 β-glucuronide, dehydroepiandrosterone sulfate and estrone sulfate, hydrophilic organic anions such as benzylpenicillin and para-aminohippurate, and the organic cations ranitidine and cimetidine [
71]. Therapeutic agents that could be transported out of the brain by SLC22A8 include several antivirals and antibiotics [
59,
71]. SLC22A11 is also known to be expressed at the BBB, but its exact localization is yet to be confirmed. SLC22A11 acts as a bidirectional and sodium-independent transporter for substrates such as prostaglandins, estrone sulfate, and ochratoxin A [
77,
78].
2.2.3. Organic Cation Transporters (SLC22s Formally OCTs)
Depending on their transport capabilities, transporters that translocate organic cations are categorized into two main groups: oligospecific organic cation transporters and polyspecific organic cationic transporters [
79]. Oligospecific organic cation transporters enable transport of a single main substrate (or its analogs); this group includes sodium glucose co-transporters (SLC5-family), sodium chloride co-transporters for neurotransmitters (SLC6-family), and high-affinity thiamine (vitamin B1) transporters (SLC19-family) [
80,
81,
82,
83]. In contrast, polyspecific organic cation transporters transport organic cations from a diversity of chemical structures. This includes endogenous compounds, such as dopamine, serotonin, monoamine neurotransmitters, acetylcholine and histamine, drugs and xenobiotics such as cimetidine, metformin, and phenformin, acyclovir, ganciclovir, memantine, and quinidine, and other compounds such as creatinine and guanidine [
79].
Polyspecific organic cation transporters belong to a large transporter subfamily named organic cation transporters (OCTs). The transporters are part of the solute carrier family SLC22A, with three members currently described in humans: SLC22A1 (OCT1), SLC22A2 (OCT2), and SLC22A3 (OCT3); the organic cation and carnitine transporters (OCTNs) have two designated isoforms reported in humans: SLC22A4 (OCTN1) and SLC22A5 (OCTN2) [
58,
82]. By definition, OCTs are involved in the transport of organic cations or weakly alkaline substrates, including several orally administered drugs that contain positive charges at physiological pHs [
84]. SLC22A1, SLC22A2, and SLC22A3 constitute the first SLC22 subgroup of functionally characterized transporters mostly localized to the basolateral membrane of polarized cells including the BBB endothelial cells and choroid plexus epithelial cells [
14]. In vitro studies have demonstrated the expression of SLC22A1 using human brain endothelial cells that mediated the transport of the antiepileptic drug lamotrigine [
85]. Other studies also linked OCTs to the transport of a number of drugs at the BBB including neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), amisulpride, oxycodone, and sulpiride [
85,
86,
87]. Regarding the OCTNs, functional studies using in vivo and in vitro models involving the transport of acetyl-L-carnitine showed that SLC22A5 is localized at the luminal side of the BBB, while SLC22A4 has not been reported in human brain and BBB. [
88,
89].