2.3. Crocodylomorpha
The crocodylomorph record in the Western Tremp Syncline is dominated by eusuchians. Two skulls belonging to two different genera have been identified. The first one is
Arenysuchus gascabadiolorum Puértolas, Canudo, Cruzado-Caballero 2011 [
115] (MPZ 2011/184) (2), from the Elias site near Arén/Areny. Phylogenetically, MPZ 2011/184 was initially placed within Crocodyloidea (crown-group Crocodylia) [
115], but later cladistic studies have situated it as a more basal eusuchian within Allodaposuchidae [
116,
117,
118,
119,
120]. The second species is the allodaposuchid
Agaresuchus subjuniperus Puértolas-Pascual, Canudo, Moreno-Azanza 2014 [
121] (MPZ 2012/288) (2). MPZ 2012/288 was initially identified as a member of the genus
Allodaposuchus [
121], but it was later reassigned to
Agaresuchus [
119]. This crocodylomorph comes from the Amor 3 site near the town of Serraduy, from one of the uppermost levels of the ‘Lower Red Garumnian’. As such, it could be one of the youngest crocodylomorphs on the Ibero-Armorican island before the K-Pg extinction. In addition, allodaposuchids are also represented by isolated teeth in several sites throughout the C30n−C29r interval [
23,
64,
93,
94,
122]. All these teeth are conical with pointed crowns, showing the typical morphology of crocodylomorphs with a generalist diet. These dental morphologies have been observed in several allodaposuchid species from the Late Cretaceous of Europe (e.g., [
122]). As the presence of other crocodylomorph clades with generalist dentition cannot be ruled out, these teeth were assigned to cf. Allodaposuchidae, since this is the most abundant clade in this region and time interval.
Gavialoidea is another clade of crocodylomorphs that may be present in the upper Maastrichtian of the Western Tremp Syncline. A few elongated conical teeth with basiapical ridges have been assigned to cf.
Thoracosaurus. These are restricted to the transitional environments of the Arén Fm and the ‘Grey Garumnian’ unit of the Tremp Fm close to Arén/Areny, Beranuy and Serraduy () [
23,
64,
122].
Hylaeochampsidae are represented by tribodont teeth from the Blasi 2B site, which were identified as cf.
Acynodon (MPZ-2017/1137) [
23,
64,
72,
122]. The eusuchian record is augmented by teeth, osteoderms, and vertebrae from the Blasi and Serraduy sites, whose taxonomical position within Eusuchia is difficult to assign with precision. They are accordingly identified as Eusuchia indet. [
23,
64,
72,
122].
López-Martínez et al. (2001) [
72] pointed out the presence of “trematochampsid”-like and alligatoroid teeth from the sites of Blasi 1, 2, and 3. However, although the authors did not provide pictures or specimen numbers, the morphotypes in question probably correspond to more recently erected taxa that had not been described at the time of publication of that paper. The “trematochampsid”-like teeth may correspond to non-eusuchian crocodylomorphs more typical of the Late Cretaceous of Europe, such as
Sabresuchus or
Doratodon, and the alligatorid teeth probably correspond to allodaposuchids. In addition, Blanco et al. (2020) [
122] mentioned the presence of a conical tooth (MPZ 2010/948) with enamel striations and crenulated carinae assigned to Mesoeucrocodylia indet.
There are also crocodylomorph eggshells from the Blasi 2 site (upper part of C30n, ). These were first reported as megaloolithid eggshells [
111], but they were later [
123] described as having a crocodyloid morphotype and were identified as
Krokolithes sp. Hirsch, 1985 [
124], implying that these eggs were laid by crocodylomorphs. Something similar has occurred with the eggshells found at the Veracruz 1 site close to Biascas de Obarra,, which were first identified as hadrosaurid eggshells [
94] (4), but after a more thorough study, their crocodylomorph affinities have been ascertained, and a description is in preparation.
Finally, the crocodylomorph record also includes several swimming and plantigrade tracks from the Serraduy, Beranuy, and Campo outcrops, all within chron C29r [
23,
125,
126] (3). This is the youngest record of crocodylomorph tracks in Europe. These tracks represent digit scratch marks produced by the manus and pes of buoyant crocodylomorphs, and they have been assigned to the ichnogenus
Characichnos. One pedal impression has been assigned to cf.
Crocodylopodus, and although its assignation cannot be confirmed with certainty due to the scarce material, this is the youngest occurrence of this ichnotaxon [
125].
2.6. The Tetrapod Fossil Record from the Upper Maastrichtian of the Ibero-Armorican Island
The tetrapod fossil record of the Western Tremp Syncline adds several unique taxa to the late Maastrichtian assemblages of the Ibero-Armorican island, yielding the youngest record of some groups prior to the Paleocene. To date, the upper Maastrichtian record of the Ibero-Armorican island is limited to the South Pyrenean Basin in northeast Spain; the Sobrepeña Fm, Torme Fm, and equivalent outcrops in northwest Spain [
130,
131]; east Spain near Tous (Valencia) [
132,
133]; and the Haute-Garonne and Aude departments in southern France [
33,
134]. During the Maastrichtian, the dinosaur faunas underwent a change in dominant herbivores during the so-called “Maastrichtian Dinosaur Turnover” [
33,
34,
135]. During the early Maastrichtian, ecosystems were inhabited by rhabdodontid ornithopods, titanosaurian sauropods and ankylosaurs, whereas in the late Maastrichtian, these communities were replaced by hadrosaurid ornithopods and new titanosaurian forms. However, nodosaurid ankylosaurians still persisted up to chron C30r, coexisting with these new assemblages for nearly 2 Myr [
33].
Lambeosaurine hadrosaurids were present in the Ibero-Armorican island from the late early Maastrichtian, mostly recorded from the Pyrenees (Spanish and French). The lambeosaurine from Els Nerets (Vilamitjana, Catalonia, NE Spain) is the oldest evidence of hadrosaurids in Europe, which was dated to within chron C31r [
81]. Lambeosaurines are also present within chron C29r, with some fossils falling very close to the K-Pg boundary. At present, there are five species of lambeosaurine hadrosaurids described from the region, comprising
Adynomosaurus from the Costa de Les Solanes site (Basturs, Catalonia) [
136],
Arenysaurus [
62],
Blasisaurus [
82] from Areny and Blasi sites (Ribagorza, Aragón, NE Spain),
Pararhabdodon from the Sant Romà d’Abella site (Lleida, Catalonia, NE Spain) [
80,
137,
138,
139], and
Canardia from the Lacarn and Tricouté sites (Haute-Garonne, southern France) [
80]. Additional hadrosauroid remains include the aforementioned lambeosaurinae from Els Nerets [
81] and other indeterminate lambeosaurines from Basturs Poble and Les Llaus (Lleida, Catalonia) [
80,
140,
141,
142]; a non-hadrosaurid hadrosauroid from Fontllonga-R (Fontllonga, Catalonia, NE Spain) [
75]; an indeterminate euhadrosaurid from Blasi 3,4 [
78]; and a small hadrosaurid from Serraduy [
79]. Outside the Pyrenees, there is a dentary from La Solana (Tous, Valencia, E Spain) that has been identified as belonging to an indeterminate hadrosaurid [
75,
143]. Finally, there is a hadrosauroid femur from the Albaina site (Laño, Condado de Treviño, Burgos, NW Spain) [
144]. With six to twelve taxa, hadrosauroids are the most speciose clade of dinosaurs in the Tremp Basin, five to eight of them being lambeosaurine hadrosaurids (, and ).
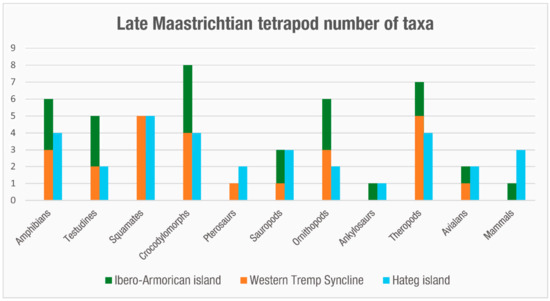
Figure 7. Bar chart with the minimum number of tetrapod taxa (genera) present in the Ibero-Armorican island and Haţeg island during the late Maastrichtian.
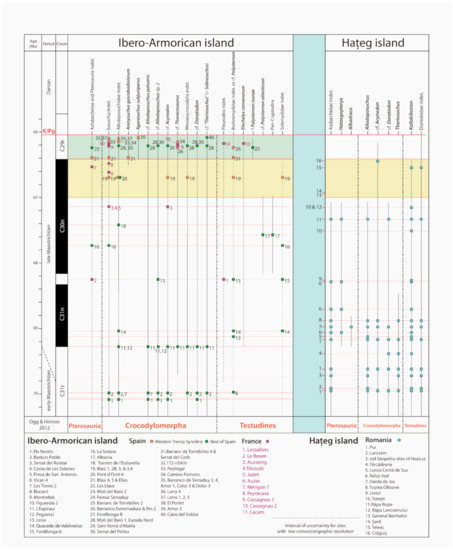
Figure 8. Dinosaur groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data are based on [
32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [
153] and calibration for the K-Pg boundary based on [
28].
Table 1. Number of tetrapod taxa present on the Ibero-Armorican and Hațeg islands during the late Maastrichtian. Red numbers mark possible additional taxa.
Ankylosaurs are represented during the late Maastrichtian by isolated and fragmentary material referred to nodosaurids from several sites in the Southern Pyrenees within the Lleida province (Catalonia, NE Spain), including Els Nerets [
145], Fontllonga-6 [
111], and Biscarri [
146]. They are also present at the Lestaillats site, in the Petite Pyrénées (Haute-Garonne, southern France) [
134]. Their last occurrence is documented at the Fontllonga-6 and Lestaillats sites, dated to within chrons C30r and C30n ().
Titanosaurs from the upper Maastrichtian of the Ibero-Armorican island consist mainly of three undetermined but distinct taxa represented by three femur morphotypes [
71] ( and ). The femur from Serraduy corresponds to a large titanosaur, whereas the other two femora represent small-medium titanosaurs. Although not formally described, these titanosaurs represent different taxa from those of the early Maastrichtian assemblage [
71]. This distinction is additionally supported by the distinct ootaxa association reported from the pre- and post-turnover assemblages, respectively [
33,
99].
Theropods from the late Maastrichtian of the Ibero-Armorican island are mainly abelisaurids and maniraptorans [
64,
93,
108,
147,
148], and they have been found only in the South-Pyrenean Basin. Due to their fragmentary and incomplete nature, the number of taxa is difficult to determine. Based on tooth morphotypes from the Southern Pyrenees, at least one abelisaurid taxon inhabited the island during the late Maastrichtian (Theropoda indet. 1 and 2 or cf.
Arcovenator; [
108]) ( and ). Maniraptorans are also represented mainly by teeth from the Southern Pyrenees, with at least three taxa identified in this way (
Richardoestesia,
Paronychodon, and Dromaeosauridae indet. from [
108]), and by the troodontid
Tamarro insperatus Sellés, Vila, Brusatte, Currie and Galobart 2021 [
149], which were recently described on the basis of skeletal remains (, and ). However, the real abundance of theropods is hard to establish, since there are several fragmentary skeletal remains attributable to undetermined dromaeosaurids, and an oological record comprising several ootaxa of maniraptoran-like eggshells, including
Prismatoolithus trempii Sellés, Vila, Galobart 2014 [
150], and
Pseudogeckoolithus Vianey-Liaud and López-Martínez 1997 [
151] ([
98,
113,
150]). Avialan dinosaurs are represented by the giant ornithuromorph bird from Beranuy [
110] and a putative enantiornithine from southern France [
152] (, and ).
The pterosaur record in the late Maastrichtian is scarce, with some isolated and fragmentary bones from the Pyrenees of France [
154,
155,
156] and Spain [
114] and from the upper Maastrichtian outcrops near Valencia (Spain) [
157,
158] (). All of them have been identified as belonging to undetermined giant azhdarchids.
Figure 9. Pterosaur, crocodylomorph, and testudines groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data based on [
32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [
153] and calibration for the K-Pg boundary based on [
28].
During the late Maastrichtian, the crocodylomorphs of the Ibero-Armorican island show great abundance, with a similar number of taxa to that during the early Maastrichtian [
23]. The best-represented clade is the eusuchian Allodaposuchidae, with two taxa described from the Western Tremp Syncline (
Agaresuchus subjuniperus and
Arenysuchus), and probably
Allodaposuchus palustris, whose characteristic teeth have been found up to chron C29r [
122,
148]. In addition, Blanco et al. (2020) [
122] described
Allodaposuchus sp. 2 on the basis of a dentary from the Fontllonga-6 site (C30r), which seems to be different from the allodaposuchids previously described and could represent a new taxon. Finally, there are plenty of isolated teeth of allodaposuchids [
64,
121,
148,
159] that due to their conical generalist shape are difficult to ascribe to specific taxa.
Gavialoidea is represented by a skull and other associated remains from the site of Cassagnau (Haute-Garonne, southern France). These have been ascribed to
Thoracosaurus neocesariensis [
160]. This assignment has been debated, as the remains could belong to a new taxon [
161]. In addition, more teeth referred to cf.
Thoracosaurus have been found in the Spanish Pyrenees, including the record of the Western Tremp Syncline [
64,
122].
The diversity of hylaeochampsids, “atoposaurids”, and notosuchians during the late Maastrichtian is difficult to determine, since most of their fossils are isolated teeth. There are several teeth referred to cf.
Acynodon from France [
162,
163] and the Spanish Pyrenees, including the Western Tremp Syncline [
122]. “Atoposaurids” are represented by teeth identified as cf.
Theriosuchus; these are from the Spanish Pyrenees [
122,
148] but not the Western Tremp Syncline. “Atoposauridae” is here written in quotes, since Tennant et al. (2016) [
164] have argued that some taxa assigned to this clade, such as
“Theriosuchus” ibericus and
“Theriosuchus” sympiestodon, belong to Paralligatoridae and have accordingly grouped these taxa under the new genus
Sabresuchus. There are also some teeth from the Spanish Pyrenees identified as the notosuchian cf.
Doratodon [
122,
148]. It should further be noted that plenty of undetermined eusuchian and crocodylomorph remains have been discovered in the French and Spanish Pyrenees (see [
23] and references therein), as well as fossil tracks of crocodylomorphs [
125], but due to their limited diagnostic value, it is difficult to ascertain their taxonomic status more precisely. There are also indeterminate eusuchian remains from La Solana (Valencia) [
132] and Quecedo de Valdivielso (Burgos, NW Spain) [
165,
166]. Thus, Ibero-Armorican crocodylomorphs are represented during the late Maastrichtian by a minimum of eight taxa (, and ).
The record of testudines during the late Maastrichtian of the Ibero-Armorican island is poorer than during the early Maastrichtian. In the Pyrenees, pleurodiran turtles are represented by the bothremydid
Elochelys convenarum Laurent, Tong, Claude [
167] from southern France and another bothremydid turtle from Isona. This represents the species
Polysternon isonae Marmi, Luján, Riera, Gaete, Oms, Galobart [
168], although Pérez-García [
169] considers this a
nomen dubium, lacking enough diagnostic characters for a new species, and classifies the remains as Foxemydina indet. Isolated remains of indeterminate bothremydids are also present in other sites in the Pyrenees [
128,
170] and in the northwestern Spanish sites of Urria and Quecedo de Valdivielso (Burgos) [
165,
166]. In the fossil site of Albaina, there is a plate identified as cf.
Polysternon atlanticum [
144]. Pan-cryptodirans are represented by the remains of solemydid turtles from the Pyrenees, from the sites of Blasi and Fontllonga-6 [
128,
170], and from La Solana (Valencia) [
132,
133]. Pereda-Suberbiola et al. (2015) [
144] describe a plate from a putative pan-cryptodiran that differs from solemydids. This makes a minimum of three pan-pleurodirans and two pan-cryptodirans in the Ibero-Armorican island during the late Maastrichtian (, and ).
Small-sized upper Maastrichtian tetrapods from the Ibero-Armorican island are represented only by amphibians and squamates from the Spanish and French Pyrenees [
129,
134,
152,
171] and from Valencia [
172] (). The first group consists of albanerpetontids, with at least one taxon present, identified in Blasi 2 as
Albanerpeton aff.
nexuosum [
129], plus several albanerpetontid remains from the L’Espinau and Serrat del Rostiar 1 sites (Lleida, Catalonia, NE Spain) [
171], Cassagnau 1 (Haute-Garonne, southern France) [
134,
152], and La Solana [
172]. In La Solana, the presence of a salamandrid is also documented [
172]. Anurans may be represented by at least four different groups with one discoglossid and one palaeobatrachid recognized at Blasi 2, L’Espinau, and Serrat del Rostiar [
129,
171], and an alytid and a putative pelobatid or gobiatid at L’Espinau [
171]. It is noteworthy that there are remains of a palaeobatrachid from Valencia [
172] that shows differences from the Blasi 2 taxon and could represent another taxon.
Figure 10. Squamate, amphibian and mammal groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data based on [
32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [
153] and calibration for the K-Pg boundary based on [
28].
Upper Maastrichtian squamates are represented by the previously described fossils from Blasi 2 [
129], with two undetermined lizards, one anguid lizard, and an alethinophid snake. Additionally, in the Pyrenees, the site of Serrat del Rostiar 1 (Lleida, Catalonia) has yielded several squamate remains [
171] including geckos, anguid, and “scincomorph” lizards, and an indeterminate iguanid. An indeterminate iguanid can also be found at L’Espinau. The Serrat del Rostiar 1 site is dated to within chron C31r in the early Maastrichtian, but due to its stratigraphic position, it lies very close to the boundary with the late Maastrichtian, so we have extended its faunal assemblage to the lower part of the late Maastrichtian (). In the French Pyrenees, there is also evidence of a large varanoid, “scincomorph” lizards, and other indeterminate squamates [
134,
152]. Outside the Pyrenees, there are also undetermined squamate remains at the La Solana site [
132].
It is interesting to note that during the late Maastrichtian, there is almost no evidence of mammals in the Spanish record of the Ibero-Armorican island, despite the fact that their presence is documented during the early Maastrichtian [
173,
174,
175] and the earliest Paleocene [
176,
177]. The only evidence of mammals during the late Maastrichtian is some tribosphenic teeth from the Peyrecave site, in the Petites Pyrénées (Haute-Garonne, southern France). These would have belonged to a therian mammal [
152,
178].