Macrophages are innate immune cells pivotal for tissue homeostasis, removal of superfluous cells, and inflammatory responses to infections. Macrophages also play diverse roles in cancer development, ranging from antitumor activity in early progression stages to, most commonly, tumor-promoting roles in established cancer. Notably, macrophages are highly plastic cells and, depending on the microenvironmental cues in the Tumor Microenvironment (TME), can undergo marked changes in their function. In established cancers, high macrophage infiltration often strongly associates with poor prognosis or tumor progression in many types of solid tumors, including breast, bladder, head and neck, glioma, melanoma, and prostate cancer. Conversely, in colorectal and gastric cancers, high macrophage infiltration correlates with a better prognosis. These apparently opposite effects are likely related to macrophage plasticity and resultant heterogeneity of phenotype and functions in various cancers.
- tumor-associated macrophages
- immunotherapy
- tumor microenvironment
- tumor
- immune suppression
- macrophage
1. The Role of Macrophages in the Tumor-Promoting Inflammation
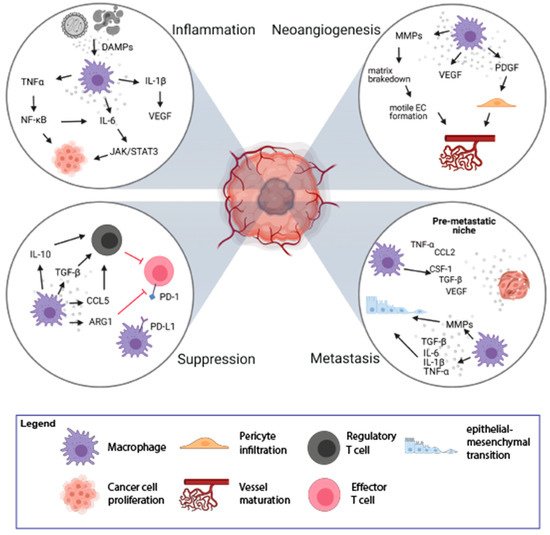
2. Macrophages and Neoangiogenesis
3. Immune Suppression and Orchestration of the Tumor Microenvironment by TAMs
4. TAMs in Tissue Invasion and Distant Metastasis
5. The Role of M1 TAMs in the Elimination of Cancer Cells
This entry is adapted from the peer-reviewed paper 10.3390/cancers13081946
References
- Glauben Landskron; Marjorie De La Fuente; Peti Thuwajit; Chanitra Thuwajit; Marcela A. Hermoso; Chronic Inflammation and Cytokines in the Tumor Microenvironment. Journal of Immunology Research 2014, 2014, 1-19, 10.1155/2014/149185.
- Jing Wang; Danyang Li; Huaixing Cang; Bo Guo; Crosstalk between cancer and immune cells: Role of tumor‐associated macrophages in the tumor microenvironment. Cancer Medicine 2019, 8, 4709-4721, 10.1002/cam4.2327.
- Florian R. Greten; Sergei I. Grivennikov; Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27-41, 10.1016/j.immuni.2019.06.025.
- Ashleigh R. Poh; Matthias Ernst; Targeting Macrophages in Cancer: From Bench to Bedside. Frontiers in Oncology 2018, 8, 49, 10.3389/fonc.2018.00049.
- Rebecca S. Cook; Kristen M. Jacobsen; Anne M. Wofford; Deborah DeRyckere; Jamie Stanford; Anne L. Prieto; Elizabeth Redente; Melissa Sandahl; Debra M. Hunter; Karen E. Strunk; et al. MerTK inhibition in tumor leukocytes decreases tumor growth and metastasis. Journal of Clinical Investigation 2013, 123, 3231-3242, 10.1172/jci67655.
- Longzheng Xia; Shiming Tan; Yujuan Zhou; Jingguan Lin; Heran Wang; Linda Oyang; Yutong Tian; Lu Liu; Min Su; Hui Wang; et al. Role of the NFκB-signaling pathway in cancer. OncoTargets and Therapy 2018, ume 11, 2063-2073, 10.2147/ott.s161109.
- Stephen J Thomas; J A Snowden; Martin P Zeidler; Sarah Danson; The role of JAK/STAT signalling in the pathogenesis, prognosis and treatment of solid tumours. British Journal of Cancer 2015, 113, 365-371, 10.1038/bjc.2015.233.
- Kevin J. Baker; Aileen Houston; Elizabeth Brint; IL-1 Family Members in Cancer; Two Sides to Every Story. Frontiers in Immunology 2019, 10, 1197, 10.3389/fimmu.2019.01197.
- Sergei I. Grivennikov; Florian R. Greten; Michael Karin; Immunity, Inflammation, and Cancer. Cell 2010, 140, 883-899, 10.1016/j.cell.2010.01.025.
- Lisa M. Coussens; Zena Werb; Inflammation and cancer. Nature 2002, 420, 860-867, 10.1038/nature01322.
- Soyoung A. Oh; Ming O. Li; TGF-β: Guardian of T Cell Function. The Journal of Immunology 2013, 191, 3973-3979, 10.4049/jimmunol.1301843.
- Naoyo Nishida; Hirohisa Yano; Takashi Nishida; Toshiharu Kamura; Masamichi Kojiro; Angiogenesis in cancer. Vascular Health and Risk Management 2006, 2, 213-219, 10.2147/vhrm.2006.2.3.213.
- Roberta Lugano; Mohanraj Ramachandran; Anna Dimberg; Tumor angiogenesis: causes, consequences, challenges and opportunities. Cellular and Molecular Life Sciences 2019, 77, 1745-1770, 10.1007/s00018-019-03351-7.
- D Ribatti; B Nico; E Crivellato; A Vacca; Macrophages and tumor angiogenesis. Leukemia 2007, 21, 2085-2089, 10.1038/sj.leu.2404900.
- Vladimir Riabov; Alexandru Gudima; Nan Wang; Amanda Mickley; Alexander Orekhov; Julia Kzhyshkowska; Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Frontiers in Physiology 2014, 5, 75, 10.3389/fphys.2014.00075.
- Saray Quintero-Fabián; Rodrigo Arreola; Enrique Becerril-Villanueva; Julio César Torres-Romero; Victor Arana-Argáez; Julio Lara-Riegos; Mario Alberto Ramírez-Camacho; María Elizbeth Alvarez-Sánchez; Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Frontiers in Oncology 2019, 9, 1370, 10.3389/fonc.2019.01370.
- Victor Ljl Thijssen; Yvette Wj Paulis; Patrycja Nowak-Sliwinska; Katrin L Deumelandt; Kayoko Hosaka; Patricia Mmb Soetekouw; Anca M. Cimpean; Marius Raica; Patrick Pauwels; Joost J Van Den Oord; et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. The Journal of Pathology 2018, 246, 447-458, 10.1002/path.5152.
- Aline Lopes Ribeiro; Oswaldo Keith Okamoto; Combined Effects of Pericytes in the Tumor Microenvironment. Stem Cells International 2015, 2015, 1-8, 10.1155/2015/868475.
- Michele De Palma; Daniela Biziato; Tatiana V. Petrova; Microenvironmental regulation of tumour angiogenesis. Nature Cancer 2017, 17, 457-474, 10.1038/nrc.2017.51.
- Michele De Palma; Mary Anna Venneri; Rossella Galli; Lucia Sergi Sergi; Letterio S. Politi; Maurilio Sampaolesi; Luigi Naldini; Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer Cell 2005, 8, 211-226, 10.1016/j.ccr.2005.08.002.
- Kanako Shimizu; Tomonori Iyoda; Masahiro Okada; Satoru Yamasaki; Shin-Ichiro Fujii; Immune suppression and reversal of the suppressive tumor microenvironment. International Immunology 2018, 30, 445-455, 10.1093/intimm/dxy042.
- Dario A. A. Vignali; Lauren W. Collison; Creg J. Workman; How regulatory T cells work. Nature Reviews Immunology 2008, 8, 523-532, 10.1038/nri2343.
- Roy Noy; Jeffrey W. Pollard; Tumor-Associated Macrophages: From Mechanisms to Therapy. Immunity 2014, 41, 49-61, 10.1016/j.immuni.2014.06.010.
- Manuela Liguori; Chiara Buracchi; Fabio Pasqualini; Francesca Bergomas; Samantha Pesce; Marina Sironi; Fabio Grizzi; Alberto Mantovani; Cristina Belgiovine; Paola Allavena; et al. Functional TRAIL receptors in monocytes and tumor-associated macrophages: A possible targeting pathway in the tumor microenvironment. Oncotarget 2016, 7, 41662-41676, 10.18632/oncotarget.9340.
- Kurt Alex Schalper; Daniel Carvajal-Hausdorf; Joseph McLaughlin; Vamsidhar Velcheti; Lieping Chen; Miguel Sanmamed; Roy S Herbst; David L Rimm; Clinical significance of PD-L1 protein expression on tumor-associated macrophages in lung cancer. Journal for ImmunoTherapy of Cancer 2015, 3, P415-P415, 10.1186/2051-1426-3-s2-p415.
- Muhammad Zaeem Noman; Giacomo Desantis; Bassam Janji; Meriem Hasmim; Saoussen Karray; Philippe Dessen; Vincenzo Bronte; Salem Chouaib; PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. Journal of Experimental Medicine 2014, 211, 781-790, 10.1084/jem.20131916.
- Andrew M. Intlekofer; Craig B. Thompson; At the Bench: Preclinical rationale for CTLA-4 and PD-1 blockade as cancer immunotherapy. Journal of Leukocyte Biology 2013, 94, 25-39, 10.1189/jlb.1212621.
- K. Vandenborre; S. W. Van Gool; A. Kasran; J. L. Ceuppens; M. A. Boogaerts; P. Vandenberghe; Interaction of CTLA-4 (CD152) with CD80 or CD86 inhibits human T-cell activation. Immunology 1999, 98, 413-421, 10.1046/j.1365-2567.1999.00888.x.
- Petar J. Popovic; Iii Herbert J. Zeh; Juan B. Ochoa; Arginine and Immunity. The Journal of Nutrition 2007, 137, 1681S-1686S, 10.1093/jn/137.6.1681s.
- Malgorzata Czystowska-Kuzmicz; Anna Sosnowska; Dominika Nowis; Kavita Ramji; Marta Szajnik; Justyna Chlebowska-Tuz; Ewa Wolinska; Pawel Gaj; Magdalena Grazul; Zofia Pilch; et al. Small extracellular vesicles containing arginase-1 suppress T-cell responses and promote tumor growth in ovarian carcinoma. Nature Communications 2019, 10, 1-16, 10.1038/s41467-019-10979-3.
- Paulo C. Rodriguez; David G. Quiceno; Augusto C. Ochoa; l-arginine availability regulates T-lymphocyte cell-cycle progression. Blood 2006, 109, 1568-1573, 10.1182/blood-2006-06-031856.
- Yuxin Lin; Jianxin Xu; Huiyin Lan; Tumor-associated macrophages in tumor metastasis: biological roles and clinical therapeutic applications. Journal of Hematology & Oncology 2019, 12, 1-16, 10.1186/s13045-019-0760-3.
- Zhe Ge; Shuzhe Ding; The Crosstalk Between Tumor-Associated Macrophages (TAMs) and Tumor Cells and the Corresponding Targeted Therapy. Frontiers in Oncology 2020, 10, 1-23, 10.3389/fonc.2020.590941.
- Meggy Suarez-Carmona; Julien Lesage; Didier Cataldo; Christine Gilles; EMT and inflammation: inseparable actors of cancer progression. Molecular Oncology 2017, 11, 805-823, 10.1002/1878-0261.12095.
- Anne-Katrine Bonde; Verena Tischler; Sushil Kumar; Alex Soltermann; Reto A Schwendener; Intratumoral macrophages contribute to epithelial-mesenchymal transition in solid tumors. BMC Cancer 2012, 12, 35-35, 10.1186/1471-2407-12-35.
- Jeffrey Wyckoff; Weigang Wang; Elaine Y. Lin; Yarong Wang; Fiona Pixley; E. Richard Stanley; Thomas Graf; Jeffrey W. Pollard; Jeffrey Segall; John Condeelis; et al. A Paracrine Loop between Tumor Cells and Macrophages Is Required for Tumor Cell Migration in Mammary Tumors. Cancer Research 2004, 64, 7022-7029, 10.1158/0008-5472.can-04-1449.
- Qing Chen; Xiang H.-F. Zhang; Joan Massagué; Macrophage Binding to Receptor VCAM-1 Transmits Survival Signals in Breast Cancer Cells that Invade the Lungs. Cancer Cell 2011, 20, 538-549, 10.1016/j.ccr.2011.08.025.
- Binzhi Qian; Yan Deng; Jae Hong Im; Ruth J. Muschel; Yiyu Zou; Jiufeng Li; Richard A. Lang; Jeffrey W. Pollard; A Distinct Macrophage Population Mediates Metastatic Breast Cancer Cell Extravasation, Establishment and Growth. PLOS ONE 2009, 4, e6562, 10.1371/journal.pone.0006562.
- Rosandra N. Kaplan; Rebecca D. Riba; Stergios Zacharoulis; Anna H. Bramley; Loïc Vincent; Carla Costa; Daniel D. MacDonald; David K. Jin; Koji Shido; Scott A. Kerns; et al. VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 2005, 438, 820-827, 10.1038/nature04186.
- Hyunho Kim; Hyewon Chung; Jaehoon Kim; Dong‐Hee Choi; Yoojin Shin; Yong Guk Kang; Beop‐Min Kim; Sang‐Uk Seo; Seok Chung; Seung Hyeok Seok; et al. Macrophages‐Triggered Sequential Remodeling of Endothelium‐Interstitial Matrix to Form Pre‐Metastatic Niche in Microfluidic Tumor Microenvironment. Advanced Science 2019, 6, 1900195, 10.1002/advs.201900195.
- Christina E. Arnold; Peter Gordon; Robert N. Barker; Heather M. Wilson; The activation status of human macrophages presenting antigen determines the efficiency of Th17 responses. Immunobiology 2015, 220, 10-19, 10.1016/j.imbio.2014.09.022.
- Dorothée Duluc; Murielle Corvaisier; Simon Blanchard; Laurent Catala; Philippe Descamps; Erick Gamelin; Stéphane Ponsoda; Yves Delneste; Mohamed Hebbar; Pascale Jeannin; et al. Interferon-γ reverses the immunosuppressive and protumoral properties and prevents the generation of human tumor-associated macrophages. International Journal of Cancer 2009, 125, 367-373, 10.1002/ijc.24401.
- Khue G. Nguyen; Maura R. Vrabel; Siena M. Mantooth; Jared J. Hopkins; Ethan S. Wagner; Taylor A. Gabaldon; David A. Zaharoff; Localized Interleukin-12 for Cancer Immunotherapy. Frontiers in Immunology 2020, 11, 575597, 10.3389/fimmu.2020.575597.
- Xiang Zheng; Kati Turkowski; Javier Mora; Bernhard Brüne; Werner Seeger; Andreas Weigert; Rajkumar Savai; Redirecting tumor-associated macrophages to become tumoricidal effectors as a novel strategy for cancer therapy. Oncotarget 2017, 8, 48436-48452, 10.18632/oncotarget.17061.