Pulses root ideotype is a conceptual root system architecture for pulse plants. This ideotype is derived from root traits that have been documented to be beneficial under soil and climate constraints, in particular, the temperate dryland cropping systems.
- dicotyledon
- indeterminacy
- tolerance
- avoidance
- dryland cropping
- constraints
- morphology
1. Introduction
Pulses are a key seed crop both in Australia and globally due to their production value, their ability to increase soil nitrogen through fixation, forming a disease break in crop rotations, and for leaving residual moisture deeper in the soil profile for use by subsequent, deep-rooted crops [1][2]. In Australia, the annual pulse crop was valued at AUD $1.8 million in 2018 [3]. Furthermore, approximately 3 million tonnes of nitrogen are fixed annually via root nodules in pulse crops, which translates into approximately AUD $4 billion [4]. If constraints are addressed, predicted pulse production can reach 4.2 million tonnes, with a combined commodity value and farm system benefit of AUD 2 billion [3].
Considerable research effort over several decades has been expended on physiological improvements of crop adaptation to water stress [5][6]. A relatively recent line of enquiry has targeted features of the root system architecture (RSA) that specifically improve access to soil water and nutrients. The root system architecture of monocots has been thoroughly characterised in terminal drought conditions such as in that of the Northern Australian cropping system. However, there is comparatively little information available on dicotyledon roots, especially pulses. In contrast to cereals, pulses are an indeterminate crop; hence, they differ significantly in their interactions between phenology, root development and response to seasonal variation [3]. Understanding the RSA of pulses and exploring the genetic variability that favours constraint tolerance or avoidance when combined with appropriate management strategies could provide a direction to breeders to improve the productivity of pulses under challenging climate and soil conditions.
Recently, a “Steep, Deep and Cheap” ideotype [7] has been proposed to increase soil water and nitrogen acquisition in an effective manner in monocotyledons. Steep root angles and thick long laterals allow deep roots to reach soil water and nitrate at depth, whilst a structure with few laterals, large cortical cells and aerenchyma reduces the photosynthate demand on the plant (Figure 1a). The focus on deeper roots is supported by research that has demonstrated the value of water stored deep in the soil profile to seed yield in environments characterised by terminal drought [8], particularly in sub-tropical regions, where crops rely on water stored during summer fallows and “opportunity cropping” [9]. However, deep roots may not be as important in major cropping regions in temperate Southern Australia. Here, rain predominantly falls during the cropping season, heavy clay soils limit the ability of water to permeate deeper down the profile, and fallows are limited to summer when limited recharge occurs. This can result in a lack of stored subsoil water and deeper roots may not provide a benefit during water-deficit conditions. Furthermore, many cropping soils in the southern region contain soil physicochemical constraints such as high boron, sodicity, salinity, high soil strength/poor soil structure and nutrient deficiencies that potentially restrict root growth and access to soil water and nutrients [10]. Thus, there is a need to investigate root traits specific to water stress adaption in the dryland cropping systems of Southern Australia.
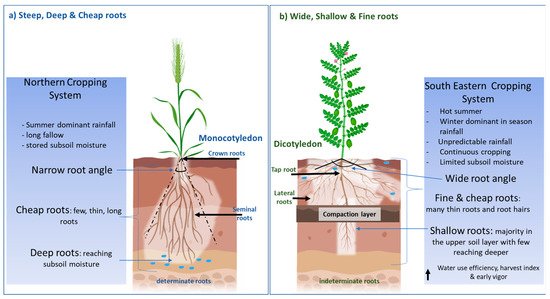
Figure 1. (a) The “Steep, Deep and Cheap” root architecture that has been largely investigated and applied to monocotyledons such as maize and wheat cultivated in environments that experience terminal drought with presence of subsoil water. (b) The proposed alternative, “Wide, Shallow and Fine” roots, for the southeastern cropping system of Australia, which experiences unpredictable rainfall and absence/unavailability of stored subsoil water.
Studies to date on pulse root system architecture (RSA) have identified key traits such as rooting depth, lateral root intensity, root length distribution, root angle and root diameter that have the potential to increase plant adaptation to water stress [11][12][13]. Ramamoorthy et al. [14] investigated some of these traits in in chickpeas and observed that genotypes with higher root length density closer to the soil surface, with greater root dry weight at depth, showed the best tolerance to drought. Similarly, Kashiwagi et al. [15] observed that genotypes with deeper roots and more soil exploration capability perform better under drought stress. Other studies, such as those by Gorim and Vandenberg [16] and Idrissi et al. [17], have examined root trait diversity in selected germplasms and found significant genetic variability in root traits and their response to drought. However, it is difficult to compare the value of various traits across existing studies due to the range of different methods that have been used. Consolidating root traits would provide breeders and agronomists with a new perspective in approaching the development of climate-ready pulse crops, especially when attempting to improve crop adaptation to specific environments.
2. Ideotype Context and Traits
2.1. Soil, Climate and Constraints
Pulse cultivation in the temperate regions of Australia with low- to mid-rainfall zones is mainly undertaken in dryland systems. Rainfall in the low-rainfall zone is <350 mm, medium 350–550, and in the high-rainfall zone, it is >550 mm. Pulses in temperate Australia are typically sown in late autumn and experience cool wet winters, with terminal drought often ending the season in early summer [18]. Existing cultivars of these crops are best adapted to alkaline soils and mostly grown on Vertosols, Sodosols and Calcarosols with a high-clay subsoil. Faba bean appears to be able to tolerate more acidic soil and waterlogging in high-rainfall areas. However, most other pulses cannot and are more limited to higher soil pH in medium- and low-rainfall areas. Topsoils can be both clay and loam, although sands become increasingly dominant in some of the lower rainfall areas. Environmental conditions in Southern Australia drylands result in a range of physicochemical and nutrient constraints, including water stress, subsoil compaction, salinity, sodicity, boron, extremes in pH and nitrogen and phosphorous deficiencies.
Despite the numerous abiotic and biotic constraints to pulse productivity in dryland cropping systems, the principal abiotic constraint is drought [10][19]. Dryland cropping systems in the medium- and low-rainfall zones of Australia typically experience unpredictable rainfall, often in insufficient amounts to replenish soil water throughout the potential rooting zone of annual crops, whilst in higher-rainfall locations, high precipitation: evaporation rates often lead to waterlogging during winter. Furthermore, the large episodic events that can occur in summer are also inefficient in recharging subsoil water storage due to high runoff and evaporation rates [18][20]. Verburg, McBeath, Armstrong, Tavakkoli, Wilhelm, Mclaughlin, Haling, Richardson, Mason, Kirkegaard and Sandral [20] found that, in certain areas, insufficient and irregular rainfall resulted in low soil water, starting at depths of only 10 cm in the soil profile.
In the case of dryland cropping systems in Southern Australia, where subsoil water can be limited due to unpredictable rainfall and continuous cropping, traits associated with developing deep roots may provide little benefit in most temperate cropping systems and may even reduce productivity as excess energy is used in developing roots that provide little value. However, critical knowledge can be extrapolated from these traits and applied to root system architecture suited to alternative seasonal rainfall patterns and soil types.
2.2. A New Root Ideotype—Wide, Shallow and Fine
Given the propensity for many pulse cropping regions to experience moderate in-season droughts, a new root ideotype that combines a range of root architectural traits is proposed. This ideotype has a root length density that is concentrated in the upper soil layers to capture in-season rainfall before it is lost due to evaporation. This high root length density would be achieved through more hypocotyl roots, a wide root angle allowing for lateral root growth in a shallow soil profile, thin lateral roots and thin xylem diameter with a high number of root hairs to increase surface area to volume ratio. Lynch [7] proposed cheap roots in terms of construction costs, by going deep quickly, and this was important to ensure a balance on the photosynthate demand on the plant. Shallow roots are inherently cheap to produce as there is no wastage on extensive root production whilst searching for water; shallow roots are already at the optimal position for water and nutrient uptake in ephemeral rainfall systems. A high water use efficiency (WUE)/transpiration efficiency and harvest index are also preferred for the optimal extraction of soil moisture and to convert it effectively to biomass and yield. Furthermore, the development of early root vigour, aimed at establishing extensive colonisation of upper soil layers when soil water is available and root penetration is easier, would likely benefit productivity. Early vigour above ground can also help to trap soil moisture by reducing evaporative losses. Early root and shoot vigour would accelerate the accumulation of shoot biomass early in the season, before water deficiency reaches limiting levels, and could act in concert with efficient translocation of assimilation to seed later in the season. The proposed ideotype would also help in competitiveness against weeds. Ramamoorthy, Lakshmanan, Upadhyaya, Vadez and Varshney [14] have also proposed a more profuse root length density in the topsoil and few thick roots at depth as adaption traits for water stress. The possession of high hereditability and plasticity in this ideotype is also important, so that in years with above-average rainfall, yield potential is still able to be fully achieved.
This entry is adapted from the peer-reviewed paper 10.3390/plants10040692
References
- Campbell, C.A.; Zentner, R.P.; Basnyat, P.; Wang, H.; Selles, F.; McConkey, B.G.; Gan, Y.T.; Cutforth, H.W. Water use efficiency and water and nitrate distribution in soil in the semiarid prairie: Effect of crop type over 21 years. Can. J. Plant Sci. 2007, 87, 815–827.
- Gan, Y.; Campbell, C.; Liu, L.; Basnyat, P.; McDonald, C. Water use and distribution profile under pulse and oilseed crops in semiarid northern high latitude areas. Agric. Water Manag. 2009, 96, 337–348.
- Pulse Australia. Pulse Australia. Available online: (accessed on 1 October 2019).
- Drew, E.; Herridge, D.; Ballard, R.; O’Hara, G.; Deaker, R.; Denton, M.; Yates, R.; Gemell, G.; Hartley, E.; Phillips, L.; et al. Inoculating Legumes: Apracticle Guide; Grains Research and Development Organisation. 2012. Available online: (accessed on 1 October 2019).
- Sreeman, S.M.; Vijayaraghavareddy, P.; Sreevathsa, R.; Rajendrareddy, S.; Arakesh, S.; Bharti, P.; Dharmappa, P.; Soolanayakanahally, R. Introgression of physiological traits for a comprehensive improvement of drought adaptation in crop plants. Front. Chem. 2018, 6, 92.
- Kebede, A.; Manjit, S.K.; Endashaw, B. Advances in mechanisms of drought tolerance in crops, with emphasis on barley. In Advances in Agronomy; Donald, L.S., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 156, pp. 266–300.
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357.
- Lilley, J.M.; Kirkegaard, J.A. Seasonal variation in the value of subsoil water to wheat: Simulation studies in southern New South Wales. Aust. J. Agric. Res. 2007, 58, 1115–1128.
- Carroll, C.; Halpin, M.; Burger, P.; Bell, K.; Sallaway, M.M.; Yule, D.F. The effect of crop type, crop rotation, and tillage practice on runoff and soil loss on a Vertisol in central Queensland. Soil Res. 1997, 35, 925.
- Adcock, D.; McNeill, A.M.; McDonald, G.K.; Armstrong, R.D. Subsoil constraints to crop production on neutral and alkaline soils in south-eastern Australia: A review of current knowledge and management strategies. Aust. J. Exp. Agric. 2007, 47, 1245–1261.
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166.
- Liu, L.; Gan, Y.; Bueckert, R.A.; Van Rees, K. Rooting systems of oilseed and pulse crops. II: Vertical distribution patterns across the soil profile. Field Crop. Res. 2011, 122, 248–255.
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13.
- Ramamoorthy, P.; Lakshmanan, K.; Upadhyaya, H.D.; Vadez, V.; Varshney, R.K. Root traits confer grain yield advantages under terminal drought in chickpea (Cicer arietinum L.). Field Crop. Res. 2017, 201, 146–161.
- Kashiwagi, J.; Krishnamurthy, L.; Purushothaman, R.; Upadhyaya, H.; Gaur, P.; Gowda, C.; Ito, O.; Varshney, R. Scope for improvement of yield under drought through the root traits in chickpea (Cicer arietinum L.). Field Crop. Res. 2015, 170, 47–54.
- Gorim, L.Y.; Vandenberg, A. Evaluation of wild lentil species as genetic resources to improve drought tolerance in cultivated lentil. Front. Plant Sci. 2017, 8, 1129.
- Idrissi, O.; Houasli, C.; Udupa, S.M.; De Keyser, E.; Van Damme, P.; De Riek, J. Genetic variability for root and shoot traits in a lentil (Lens culinaris Medik.) recombinant inbred line population and their association with drought tolerance. Euphytica 2015, 204, 693–709.
- Nuttall, J.G.; Armstrong, R.D.; Connor, D.J.; Matassa, V.J. Interrelationships between edaphic factors potentially limiting cereal growth on alkaline soils in north-western Victoria. Soil Res. 2003, 41, 277–292.
- Sadras, V.O.; Hayman, P.T.; Rodriguez, D.; Monjardino, M.; Bielich, M.; Unkovich, M.; Mudge, B.; Wang, E. Interactions between water and nitrogen in Australian cropping systems: Physiological, agronomic, economic, breeding and modelling perspectives. Crop. Pasture Sci. 2016, 67, 1019–1053.
- Verburg, K.; McBeath, T.M.; Armstrong, R.D.; Tavakkoli, E.; Wilhelm, N.; Mclaughlin, M.J.; Haling, R.E.; Richardson, A.E.; Mason, S.D.; Kirkegaard, J.A.; et al. Soil water dynamics as a function of soil and climate to inform phosphorus placement strategies. In Proceedings of the Soil Science Australia and New Zealand Scoiety of Soil Science Joint Conference, Cains, Australia, 29 November 2020.
- Verburg, K.; McBeath, T.M.; Armstrong, R.D.; Tavakkoli, E.; Wilhelm, N.; Mclaughlin, M.J.; Haling, R.E.; Richardson, A.E.; Mason, S.D.; Kirkegaard, J.A.; et al. Soil water dynamics as a function of soil and climate to inform phosphorus placement strategies. In Proceedings of the Soil Science Australia and New Zealand Scoiety of Soil Science Joint Conference, Cains, Australia, 2020.