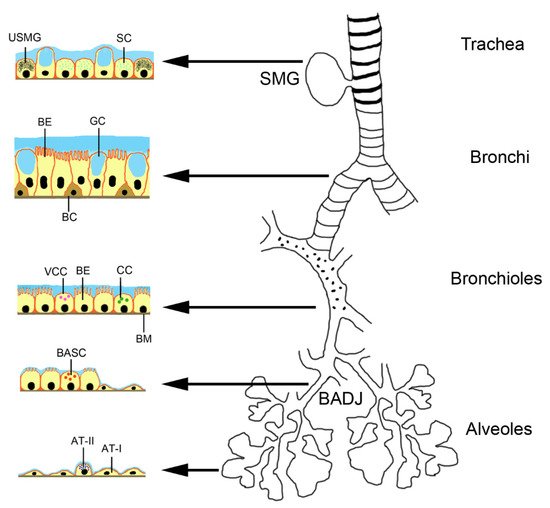
2.1. Biological Characteristics of MSCs
The use of embryonic stem cells is not undisputed and regulation for their use vary between the countries [
24]. Ethical concern of destroying embryonic cells and issues, such as tumorigenicity, risk of rejection and difficulty to obtain uniform differentiation in the target tissue, were the main reasons for preferring adult stem cells to embryonic stem cells. Adult stem cells, although they are less flexible regarding differentiation, available in smaller amounts and have a finite life span, are the preferred source for stem cell therapy. Due to their inherent plasticity, adult stem cells can differentiate into parenchymal cells. Sources for adult stem cells are blood, bone marrow, eye, brain, skeletal muscle, dental pulp, liver, gastrointestinal tract, pancreas and skin. MSCs have the advantage that, due to the lack of Major Histocompatibility Complex (MHC) class II and low class I expression, they are considered immune-evasive. They are not immune-privileged because allogenic MSCs caused a systemic inflammatory response 2h after infusion and induced antibody generation [
25]. There may be even inter-species compatibility because human MSCs were effective in rats with PAH and in hyperoxic neonatal lung injury of mice [
26,
27]. MSCs can also activate the complement activation and induce thrombosis. However, since the reaction is usually mild and the retention of the MSCs in the body is low, allogenic MSCs can be used without major problems. Another issue of concern is the potential tumorigenic action of MSCs. The reason for the concern was sarcoma formation upon MSC administration reported in a mouse study [
28]. Genetic instability (transformation) of the cells upon expansion in vitro was not experimentally verified and no tumor formation after repeated administration of MSCs to NOD mice and cynomolgus monkeys reported [
29]. Although no tumor formation has been reported in humans, studies with long-term follow up are needed to enable a definite statement on their safety.
2.2. Modes of Action of MSCs
The mechanism of the beneficial effects of MSCs is not completely understood but several modes of action have been identified. Firstly, MSCs may differentiate to replace the damaged cells. This mechanism has been reported for renal tubular epithelial cells, epidermal keratinocytes and endothelial cells [
30]. Integration into the walls of endothelial vessels has been observed after intraarterial injection of MSCs in the rat cremaster muscle microcirculation model [
31] and in a similar way, MSCs delivered directly into the lung may also integrate into the alveolar epithelium [
32]. Alternatively, cell fusion may take place because the very low frequency of cell fusion is increased in pathologic conditions. Alterations of the lipid bilayer upon inflammation are hypothesized as potential mechanism. Polymer chain reaction analysis from organs of patients, who received MSCs for various indications, showed that cell fusion contributed to the beneficial effect of MSCs, although to very low extent [
33]. MSC donor DNA was detected in one or more tissues at levels of 1‰ to 1% of the cells. A study on co-culture of heat-shocked small airway epithelial cells and MSCs reported that up to 1% of the MSCs fused with epithelial cells [
34]. Organelle transfer (mitochondria, lysosomes) may occur via thick (0.7 µm) tunneling nanotubes [
35] from MSCs to macrophages, bronchial epithelial cells and alveolar epithelial cells. However, based on the fact that survival of MSCs in the body is short and that alginate-encapsulated MSCs acted similar to not encapsulated MSCs, it is assumed that MSCs act mainly by paracrine action [
36]. This action may occur either by soluble molecules or by proteins and microRNAs (miRs) contained in extracellular vesicles (EVs). It is not easy to differentiate between soluble and vesicle-enclosed molecules because the isolation method of the EVs does not allow a good separation between the two. EVs can be released upon stimulation or after cell disruption due to shear stress in the blood circulation [
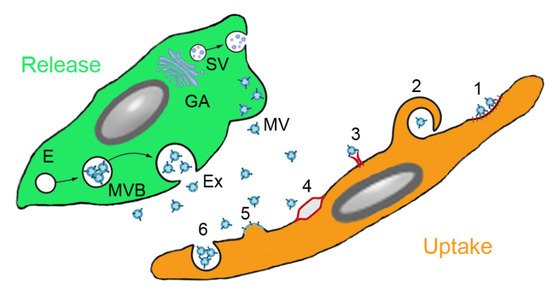
37]. Paracrine interaction of MSCs with target cells is illustrated in . Soluble factors are transported from the Golgi stacks as secretory vesicles to the plasma membrane, where they are released. Microvesicles are released by membrane budding and exosomes originate from endosomes and multivesicular bodies. Uptake by the target cells occurs either by clathrin- or caveolin-mediated endocytosis, macropinocytosis, receptor-mediated uptake, lipid rafts, fusion with the plasma membrane and (for phagocytic cells) phagocytosis.
Figure 2. Paracrine secretion by mesenchymal stem cells (MSCs) and action on target cells. MSCs produce soluble molecules at the Golgi apparatus (GA) and release them as secretory vesicles (SV). Extracellular vesicles (EVs) are secreted either from endosomes (E) via multivesicular bodies (MVB) as exosomes (Ex) or by membrane budding as microvesicles (MV). EVs can be ingested by clathrin-or caveolin-mediated endocytosis (1), macropinocytosis (2), receptor-mediated uptake (3), lipid rafts (4), fusion with the plasma membrane (5) and phagocytosis (6) (routes according to [
38]).
MSCs acted beneficial in asthma by release of EVs containing miRs to inhibit Th2 cells, in IPF by combination of inhibition of Th2 cells, stimulation of regulatory T cells (Treg) and inhibition of leucocyte infiltration via secretion of interleukin (IL)-1RA. Therapeutic effect of MSCs in COPD include secretion of epithelial growth factor (EGF), hepatocyte growth factor (HGF), keratinocyte growth factor (KGF), vascular endothelial growth factor (VEGF) to inhibit alveolar epithelial cell damage and apoptosis and release of EVs containing miR-100, miR-146a and miR-146-5b [
39]. Effects of MSCs have been studied particularly in ARDS and can be grouped into improvement of the epithelial barrier by protection of epithelial cells, tightening of the intercellular functions, improvement of clearance function and decrease of inflammation by immunomodulatory effects () [
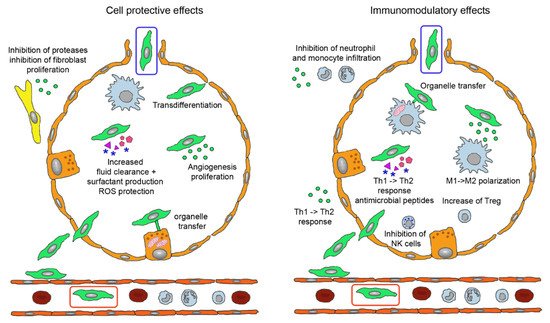
40].
Figure 3. Biological effects of mesenchymal stem cells (MSCs) administered by intravenous (green cell in red box) and inhalation route (green cell in blue box). Injected MSCs have to extravasate and either locate in perivascular position or permeate the epithelial layer to reach the alveolar lumen. They perform tissue-protective and anti-fibrotic effects (left) by integration into the epithelial layer, release of extracellular vesicles (EVs) containing Ang-1, KGF, HGF, VEGF, EGF, mRNAs, miRs, membrane components and DNA for anti-apoptotic effects, increase of lung fluid clearance, epithelial cell proliferation and angiogenesis, transfer of mitochondria to alveolar epithelial cells via nanotubules and secretion of cytokines and other proteins. EVs containing inhibitors for metalloproteinases and for fibroblast proliferation act in the perialveolar space. Immunomodulatory functions (right) comprise organelle transfer to alveolar macrophages via nanotubules, release of TGF-β, PGE2, IDO, IL-10 to decrease T cell activation, TGF-β and HLA-G5 to stimulate Treg, PGE2 and TSG-6 to inhibit M1 activation and secretion of antimicrobial proteins LL-37 and lipocalin-1. The involved mediators either are contained in EVs or present as soluble factors. Abbreviations: Ang-1, angiopoietin-1; EGF, epithelial growth factor; HGF, hepatocyte growth factor; HLA-G5, human leukocyte antigen G5; IDO, indoleamine 2,3-dioxygenase; IL-10, interleukin 10; KGF, keratinocyte growth factor; LL-37, cathelicidin; miR, micro RNA; PGE2, prostaglandin E2; Treg, regulatory T cells; TGF-β, tumor growth factor-beta; TSG-6, TNF-stimulating gene 6; VEGF, vascular endothelial growth factor.
The contributions of the specific molecules and mediation by free or membrane-enclosed molecules differ between the studies. The majority of the reported effects, namely restoration of epithelial and endothelial function by increased cellular repair and decreased rate of apoptosis, higher surfactant production, increased resorption of lung fluid, restoration of tight junctions and reduced fibrin production may be mediated by EVs. EVs are also supposed to be involved in reduction of pro-inflammatory and enhancement of anti-inflammatory cytokine secretion, reduction of neutrophil infiltration and M2 polarization of alveolar macrophages [
41]. Preclinical data identified cell-to-cell contact via programmed death-1 for MSC-T cell interaction, changes in amino acid and lipid metabolism by indoleamine 2,3-dioxygenase (IDO) expression, prostaglandin E2 (PGE2) production, tumor growth factor-beta (TGF-β) and HGF and increased expression of leukocyte protease inhibitor via EGF and HGF as important mechanisms [
42]. Epithelial repair was mediated mainly by angiopoietin-1 (Ang-1), TNF-stimulating gene 6 (TSG-6) and lipoxin A4 (LXA4) and enhanced alveolar fluid clearance through activation of epithelial sodium channel by KGF [
43]. Antimicrobiotic effects of MSCs in bacterial ARDS included increased phagocytosis of macrophages through mitochondria transfer, increased release of cathelicidin (LL-37) and of IL-10 and decreased tumor necrosis factor alpha (TNF-α) secretion mediated by PGE2 and LX4.
In bacterial/lipopolysaccharide (LPS)-induced ARDS, EVs containing Ang-1 and miR30b-3p for inhibition of epithelial damage are released by MSCs. miR-145, mitochondria and KGF stimulate oxygen consumption and phagocytosis of macrophages. MSCs inhibit secretion of IL-6 and TNF-α secretion, stimulate TSG-6 and LL-37 secretion and inhibit leukocyte invasion [
39]. MSCs display on the one hand anti-inflammatory properties but, on the other, cause positive effects in animal models for bacterial pneumonitis. Preservation of neutrophilic granulocytes activity as the first line defense is likely and transfer of mitochondria to macrophages may make them more energetic and active in antibacterial defense. Rather than a standard release profile of MSCs the described mechanisms represent a panel of potential mechanisms, which can vary according to type and generation of MSC and to disease condition.
2.3. MSC-Derived Products
Conditioned media or secretome is a mixture of all organic and inorganic products secreted by cells. Its composition is similar to plasma, which contains free and vesicle-bound molecules mainly released from epithelial cells, endothelial cells and blood cells. Isolation of EVs from MSC-derived secretomes have been used in preclinical studies of lung diseases, where conditioned media from bone marrow-derived MSCs of rat, murine and human origin acted beneficial in murine and rat models of ARDS, murine asthma, murine and rat BPD/hyperoxia and rat fibrosis models [
44]. Efficacy was obtained upon administration by intratracheal, intravenous, intranasal and intraperitoneal route. The priming of MSCs caused variable effects; while priming of MSCs by culture in hypoxia did not increase the efficacy of the secretome in ARDS model, addition of a Toll-like receptor 3 agonist had a positive effect. Differences between soluble proteins, different vesicles and whole cells may be related to disease-specific requirements: delivery of VEGF is most relevant in BPD, while fibroblast growth factor 2 (FGF-2) is the most important molecule in the treatment of COPD [
45].
There are presently no approved MSC-conditional media but convalescent plasma has been used in the treatment of several virus-based diseases, namely for the Spanish Flu pandemic and infections with the SARS, MERS and influenza virus [
46]. Plasma in addition to serum proteins (albumin and immunoglobulins) contains between 10
7 and 10
9 EVs/mL plasma in healthy individuals [
47]. The reported beneficial effects in these diseases stimulated clinical trials in severely ill COVID-19 patients. Results from the several small case studies and the large trails, however, did not show mortality benefit or reduced progression to severe disease in patients admitted to hospital with moderate COVID-19 [
48]. There is the possibility that concentrations both of transfused antibodies and of EVs was too low to induce the desired effect.
EVs are hypothesized to represent the most important active component of the secretome, although it is difficult to differentiate between soluble and membrane-enclosed molecules. EVs is a broad term to describe different types of vesicles secreted from cells. The function is communication between cells but they may also act as dustcarts. Exosomes are vesicles released by exocytosis of multivesicular bodies whereas ectosomes are assembled vesicles released at the plasma membrane [
49]. Other classification is based on size with apoptotic bodies (0.1–5 µm), microvesicles (100–1000 nm) and exosomes (35–120 nm) as the main groups. Exosomes are further classified into small (60–80 nm), large (80–120 nm) and nanosized (~35 nm) vesicles [
50]. The three types of EVs differ not only regarding size but also by origin, surface markers, content and uptake mechanism by the target cells, which is summarized in .
Table 1. Characterization of MSC-derived vesicles according to origin, size, surface marker, content and uptake mechanism by the target cells (according to [
51]). Abbreviations: ESCRT, endosomal sorting complex required for transport; TSG101, tumor susceptibility gene 101 protein.
Due to the surrounding lipid membrane, EVs prevent degradation of the transported lipid mediators (e.g., eicosanoids), proteins (cytokines, chemokines, growth factors), genetic material (mRNA, long non-coding RNAs, short non-coding RNAs/miRs, nuclear and mitochondrial DNA) and organelles (e.g., mitochondria) by enzymes. EVs in the bronchoalveolar fluid (BALF) are mainly lung-specific exosomes, suggesting that exosomes predominantly serve for local signaling [
52] with most cross talk taking place between alveolar epithelial cells and alveolar macrophages. Lung cells communicate intensely via EVs, which are released by respiratory cells at the apical and at the basal site, by alveolar macrophages, by other immune cells in the lungs and by fibroblasts [
53]. In the healthy lung, EVs released from the macrophages down-regulate secretion of cytokines by alveolar epithelial cells. In respiratory lung diseases, EVs with higher content of pro-inflammatory cytokines, metalloproteinases and lower content of suppressor molecules are found in BALF. On the other hand are lung epithelial cell derived EVs also found in blood suggesting an exchange of EVs across the air-blood barrier and effects over longer distances. The exchange of EVs between blood and lung can explain why intravenously administered EVs can influence lung physiology. Information on the mechanism of intercellular signaling by miRs and proteins contained in EVs are available in several reviews (e.g., [
52,
54,
55]).
EVs act on target cells either by release of soluble mediators, by receptor binding or by endocytosis [
56]. While receptor-mediated uptake suggests specificity, the inter-species activity of EVs supports the hypothesis of stochastic uptake. It is possible that disease-induced alterations of the cell surface affect EV uptake. Similarly, the expression of the main uptake routes, clathrin-mediated uptake, caveolin-mediated uptake, clathrin and caveolin independent uptake, macropinocytosis and phagocytosis, may be linked to cellular differences in the uptake of EVs. Clearance of EVs from air space and blood occurs usually within minutes but with prominent inter-individual differences. In addition to various mRNAs and proteins, miR-145, miR-221, miR-133b, miR-223, miR-146a and miR-let-7c are important components of EVs for the treatment of lung diseases [
57]. Despite the prominent role, which is presently attributed to miRs, calculations based on the amount of miRs in MSC-derived EVs and the possible number of EVs that could be taken up by target cells obtained concentrations, that were too low to cause relevant biological effects [
58]. These data suggest that proteins are the drivers of the biological effects of EVs.
2.4. Differences in the Actions of MSCs and EVs
EVs due to their small size can permeate epithelial barriers. Furthermore, clearance of EVs from the organism is slower. MSCs are cleared within 24 h from the circulation but EVs were detected one hour after injection in parenchymal cells and macrophages of the damaged tissue and remained there for up to 7 days [
59]. While MSCs are damaged by shear stress and retained in the pulmonary vasculature, EVs are not affects by these processes. Another advantage is the ability of storage of EVs in the absence of DMSO, which may have biological effects. Further, the absence of HLA I - and HLA II expression provides them with lower immunogenicity than MSCs, which express HLA I constitutively and HLA II after stimulation with IFN-γ [
60]. EVs do not activate the complement system or induce opsonization of antigen-presenting cells like MSCs. The immune effects are due to MSCs disrupted by shear stress in the blood circulation [
37]. Although also MSC-derived EVs have strong prothrombotic effects, MSCs induce more thrombosis because they also obstruct small blood vessels due to the large size of the cells and cell agglomeration [
61]. Thromboembolism occurs mainly in the lung, where MSCs are retained by binding to vascular cell adhesion protein 1 (VCAM-1) of endothelial cells. Despite better penetration of epithelial barriers, EVs and conditioned media may act less beneficial because they may not able to reproduce all beneficial effects of MSCs. For instance were MSCs able to prevent ischemia-perfusion kidney damage, while conditioned media was not effective [
35]. It has also been reported that exosomes were less efficient in maintaining endothelial barrier function in vitro. They acted, however, similarly efficient in a hemorrhagic shock and laparotomy induced lung injury model in vivo [
62]. The lack of cell-to-cell contact and mitochondrial transfer through tunneling nanotubes or microvesicles may explain the lower efficacy [
63]. As compensation for the lower efficacy, EVs can be loaded with small molecules, miRs, proteins and other macromolecules. This loading could be either occur exogenously after EV isolation or endogenously during biogenesis of the EVs [
64]. Exogenous loading occurs via electroporation, simple incubation, surfactant treatment, sonication, extrusion and freeze thawing. These procedures result often in aggregation of EVs or cargo and alter physicochemical properties and morphology of the EVs. In the endogenous procedure, the parent cells are transfected or co-incubated and cultured to produce EVs. Cells can also be engineered to express the molecule of interest, which is then contained in the released EVs. More details on the different loading techniques for EVs can be obtained for instance in reviews dedicated to this topic [
65,
66,
67]. Currently, the spectrum of loaded exosomes is limited to cytostatic agents like celastrol, paclitaxel, gemcitabine and taxol for use in cancer and EVs for few other indications such as spinal cord injury, Alzheimer disease and periodontal defects [
40].
One limitation may be cell-specificity of the action because exosomes from lung spheroids outperformed EVs from MSCs. Lung-resident MSCs were shown to represent a unique population with a different phenotypic and gene expression pattern than MSCs derived from other tissues [
68]. Lung MSCs compared to bone marrow MSCs expressed epithelial genes to greater extent and more remarkably differentiated to epithelial cells in retinoic acid treatment [
69]. Further, some concerns remain about the potential tumor growth promoting effects of EVs. Tumor-promoting effects of MSC-derived exosomes were observed by activation of extracellular signal-regulated kinase 1/2 (ERK1/2) signaling, transfer of tumor suppressor miR-15a, miR-410, protection against cell stress, decrease of tumor cell apoptosis, exchange of MMP-2 and ecto-5′ nucleosidase and promotion of angiogenesis [
70]. In addition, the immunomodulatory action of EVs may favor tumor growth.