2.3. Tissue Expression and Cellular Localization of OBPs
Insect OBPs were originally identified in olfactory sensilla (Vogt and Riddiford, 1981) using immuno-electron microscopy, which enable the determination of their expression patterns in the different antennal sensilla types (trichoid, basiconic and coeloconic). A comparative study conducted on three moth species, the saturniid
Antheraea polyphemus, the bombycid
Bombyx mori, and the noctuid
Autographa gamma, detected PBPs in trichoid sensilla, particularly in the extracellular sensillum lymph of the hair lumen and in the sensillum-lymph cavities. Moth PBPs were also detected in secretory organelles of the trichogen and tormogen cells, supporting the hypothesis that these cells can produce and secrete PBPs into the sensillar lymph [
33].
Recently, Larter et al. focused on the ten OBPs most abundantly expressed in the Drosophila antenna. They used in situ hybridization to map their spatial distribution in the different morphological sensilla classes (a). The expression profiles of these antennal OBPs were more precisely investigated in the basiconic sensilla subtypes using double-labelling with OBP and OR markers. The expression patterns of distinct OBP subsets in different basiconic sensilla were identified. The map reveals that ab8 and ab9 basiconic sensilla express only one abundant OBP (OBP28a), while others co-express different OBPs. Moreover, some functionally distinct basiconic sensilla contain the same subset of abundant OBPs (b) [
34]. Drosphila olfactory and gustatory sensilla house three accessory cells that surround the cell body of the sensory neurons: the thecogen, tormogen and trichogen cells which are involved in insect sensilla morphogenesis and in OBP expression in the lymph. Conversely, non-neuronal cells expressing OBPs in antennal sensilla were identified using markers labelling each accessory cell. This study revealed that OBPs can be either expressed in tormogen cells or in thecogen cells. The only exception was OBP28a, which is simultaneously expressed in both types of accessory cell [
34].

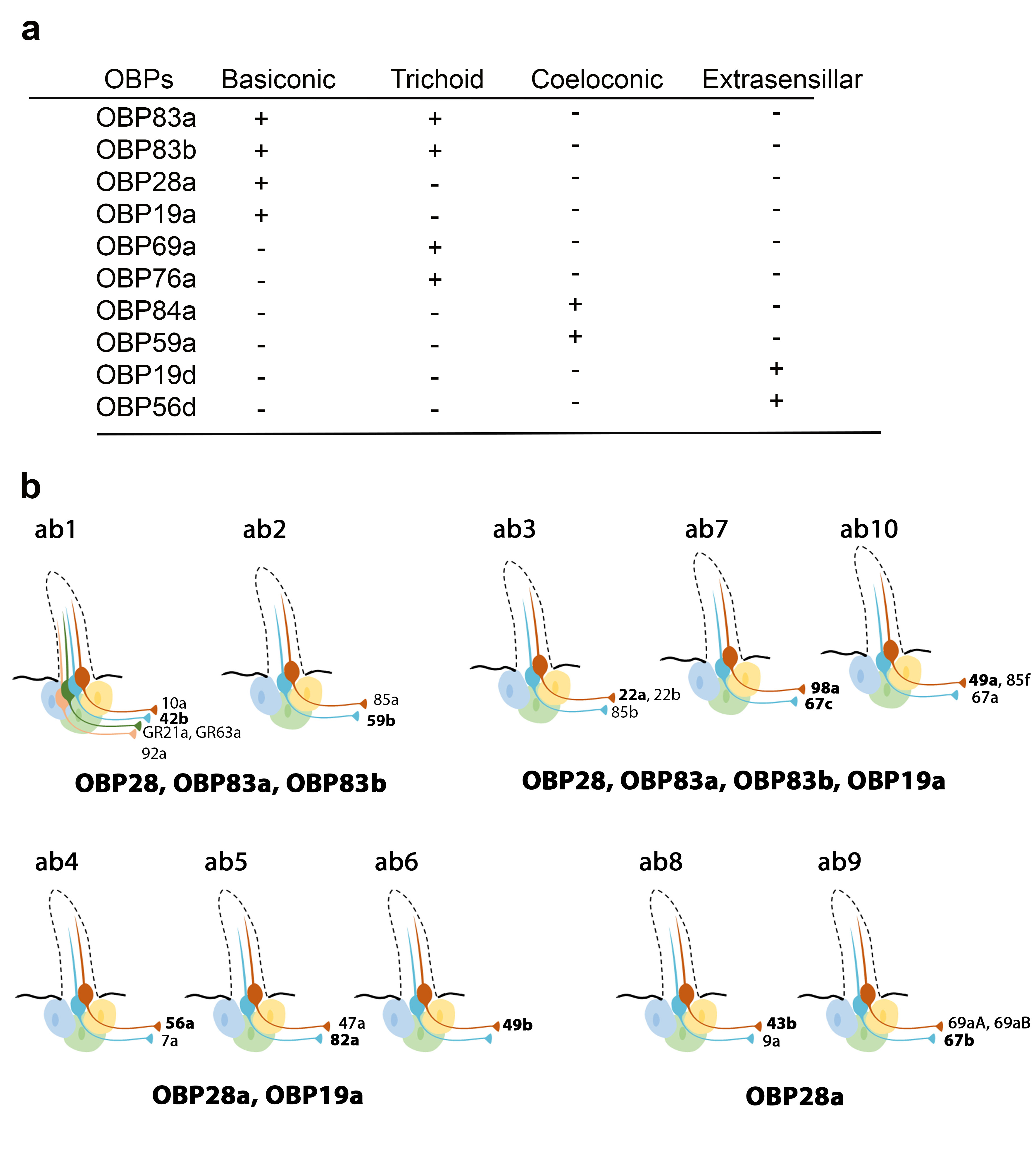
Figure 1. Expression patterns of the most abundant OBPs in Drosophila antennae. (
a) Summary of the OBP expression patterns in the three types of antennal sensilla. OBP19d and OBP56g are expressed in epidermal cells. (
b) Distribution of highly abundant OBPs expressed in Drosophila antennal basiconic sensilla. Olfactory receptor (OR) genes expressed in each olfactory receptor neuron class are indicated (all the OR genes except for GR21a and GR63a). The OR markers used for in situ hybridization for each sensilla are shown in bold [
28,
34].
The spatial and temporal expression patterns of insect OBPs have been reported in several studies, revealing that OBPs are expressed in both olfactory and taste appendages or in either chemosensory system. Gustatory OBPs have been less commonly studied than those expressed in olfactory tissues. For example, OBP57d and OBP57e are expressed in specific leg sensilla of different Drosophila species [
35], while OBP49a and OBP19b are expressed in thecogen cells of
D. melanogaster labellar sensilla [
36,
37]. The OBP19b protein was only detected in small and intermediate proboscis sensilla [
37]. Moreover, OBP19d is not only expressed in olfactory appendages of
D. melanogaster (antenna and maxillary palps) but also in adult gustatory organs (labellar bristles and pegs, legs, wings and in ventral and dorsal cibarial sense organs (VSCO)) [
38,
39] (a,b). Two
Helicoverpa armigera OBPs and one
Plutella xylostella OBP were detected in the mouthparts [
40], while OBP57e, OBP56g, OBP28a2 and OBP49a were identified in the legs of the oriental fruit fly
Bactrocera dorsalis [
41]. In
Culex pipiens quinquefasciatus adults, several OBPs were exclusively identified in olfactory tissues, while others (OBP10, OBP17, OBP18, OBP22, OBP25) were identified in taste appendages (proboscis and legs) [
42]. Similarly, taste-specific OBPs were identified in the labellum and tarsi in
Aedes aegypti and in the red flour beetle
Tribolium castaneum [
43,
44]. In the desert locust
Schistocerca gregaria, a subset of the antennal OBP repertoire is also expressed in the maxillary and the labial palps [
45]. Moreover, seven genes expressed in the labellum and tarsus of the fleshfly
Boettcherisca peregrina were identified and show sequence similarity to insect OBP genes. Homologues of these gene products were detected in
D. melanogaster taste tissues [
46]. In the legs of the two mosquito species
Anopheles gambiae and
Anopheles arabiensis, the identified OBP (agCP1564) shows high similarity to Drosophila OBP57e, which is specifically expressed in the tarsi [
47,
48] (a). Notably, 6 OBPs (OBP1, OBP2, OBP3, OBP4, OBP7 and OBP8) were found in the antenna and legs of the onion fly
Delia antiqua. Homology studies identified their
D. melanogaster homologues (OBP19d, OBP83a, OBP83b, OBP56h, OBP76a, OBP69a, respectively). Unlike
D. antiqua,
D. melanogaster homologues are only expressed in the fly antenna except for OBP19d, which is also expressed in Drosophila tarsi, and OBP56h, which is also expressed in Drosophila proboscis [
49] (a,b). Other studies have reported the expression of OBPs in insect legs: OBP7 in
B. dorsalis [
50], OBP10 in
Clostera restitura [
51] and OBP4, OBP6, OBP7, OBP8 in
Adelphocoris lineolatus [
52]. Similarly, OBPs are expressed in the legs and wings of three species of social hymenopterans (
Polistes dominulus,
Vespa crabro,
Apis mellifera) [
53]. Notably, three OBPs were identified in the anterior margin of the wings of
D. melanogaster [
39,
47] (a). The differences in OBP expression between tarsi, labellum and wings might be explained by the distinct roles of OBPs in food detection and intake.
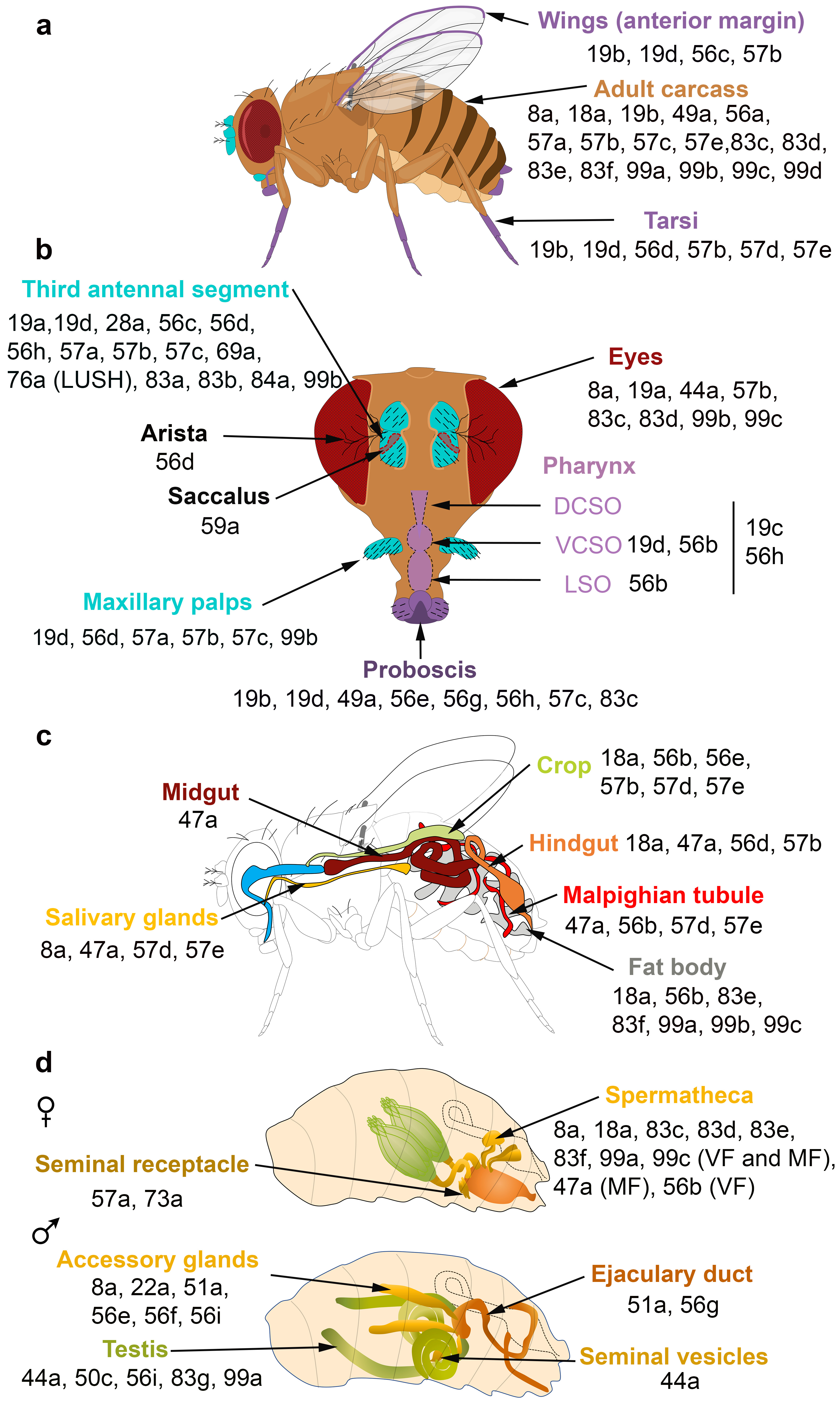
Figure 2. OBPs expressed in the olfactory, gustatory, digestive and reproductive organs of
D. melanogaster. (
a,
b) Adult taste organs (purple) consist of the proboscis, the internal structure in the pharynx (the labral sense organ (LSO), the ventral and dorsal cibarial sense organs (VCSO and DCSO)), the leg tarsi, the anterior margin of the wings and the female genitalia. Adult olfactory organs (turquoise) consist of the antenna and the maxillary palps. OBPs expressed in each organ are indicated by their number. OBPs are found in chemosensory and non-chemosensory organs. (
c) Expression patterns of OBPs in the adult digestive tract. (
d) Expression patterns of OBPs in the female and male reproductive organs. The expression of OBPs in female spermatheca depends on the sexual state. Some OBPs are expressed in virgin female (VF) and mated female (MF) spermatheca, while the expression of some OBPs is mating-dependent [
36,
37,
38,
47,
54,
55,
56,
57,
58,
59].
Proteomic and transcriptional studies confirmed the expression of a subset of insect OBPs in non-sensory organs. In the honeybee, 9 of the 21 OBPs predicted by the genomic sequence were detected in the mandibular glands [
60]. OBPs can be expressed in female and male reproductive organs. In
D. melanogaster, six OBPs (especially the abundant OBP56f and OBP56g) were detected among the seminal fluid proteins transferred to females during copulation, and three of these OBPs were found in the seminal receptacle [
55,
61,
62,
63]. Similarly, OBP10 is highly abundant in the seminal fluid of the two Lepidopteran species
Helicoverpa armigera and
H. assulta [
24]. The OBPs present in seminal fluid could be carriers of oviposition deterrents. In addition, OBP22 of the mosquito
A. aegypti [
64,
65], OBP9 of
A. mellifera [
66] and two OBPs of
Tribolium castaneum [
67] are also present in sperm. Proteomic analysis revealed OBP expression in mosquito ovaries and eggshell [
68,
69,
70]. RNAseq analyses and RT-PCR data also revealed the presence of OBPs in the ovaries of the stemborer
Sesamia nonagrioides [
23]. We can hypothesize that their accumulation in the ovaries is involved in oocyte maturation. These OBPs might also bind chemo-attractant molecules, resulting in sperm attraction. Moreover, in the oriental fruit fly
B. dorsalis, OBP44a, OBP49a, and OBP56g are highly expressed in the male testis and OBP19c is highly expressed in the female ovary [
41]. Examination of the FlyAtlas expression database reveals that OBP44a, OBP50c, OBP56i, OBP83g, and OBP99a are expressed in
D. melanogaster male testis (d). Among the 32 OBP genes annotated in the Hessian fly
Mayetiola destructor, 24 and 25 of them were found to be expressed in female and male terminal abdomens, respectively. Only OBP31 (in female) and OBP11, OBP24 and OBP32 (in male) showed relatively higher expression levels in the terminal abdomen than in the antennae [
71]. Moreover, four OBPs (OBP1, OBP4, OBP8, OBP10) were identified in the
B. dorsalis abdomen, which houses the reproductive organs. These OBPs share high sequence homology with their
D. melanogaster analogues (OBP8, OBP56d, OBP83ef and OBP99c, respectively).
D. melanogaster analogues are also expressed in different reproductive organs present in the abdomen (d) [
50]. In
Culex quinquefasciatus and
Anopheles funestus, OBP expression was also detected in the abdomen [
42,
72]. Moreover, OBP22a, OBP51a, OBP56e, OBP56f, OBP56i are highly expressed in
D. melanogaster male accessory glands (d). All these data suggest that OBPs may (i) serve to bring odorants or pheromones next to the odorant receptors present in the female reproductive tract or (ii) carry male-specific molecules into female tissue to elicit a behavioral response. It is not yet known whether these OBPs are related to fertility and fecundity features.
The FlyAtlas expression database reveals that 6 OBPs are expressed in
D. melanogaster eyes (b). Similarly, several OBPs were identified in the eyes of the lepidopteran
H. armigera [
40]. Together with other proteins, these OBPs may be implicated in the complex mechanism of vision, specifically in the generation, transport and recycling of visual pigments.
In D. melanogaster, some OBPs are expressed in both larva and adults, while others are only expressed in adults (). The majority of OBPs expressed in larva show similar expression patterns in adult tissues ( and ).
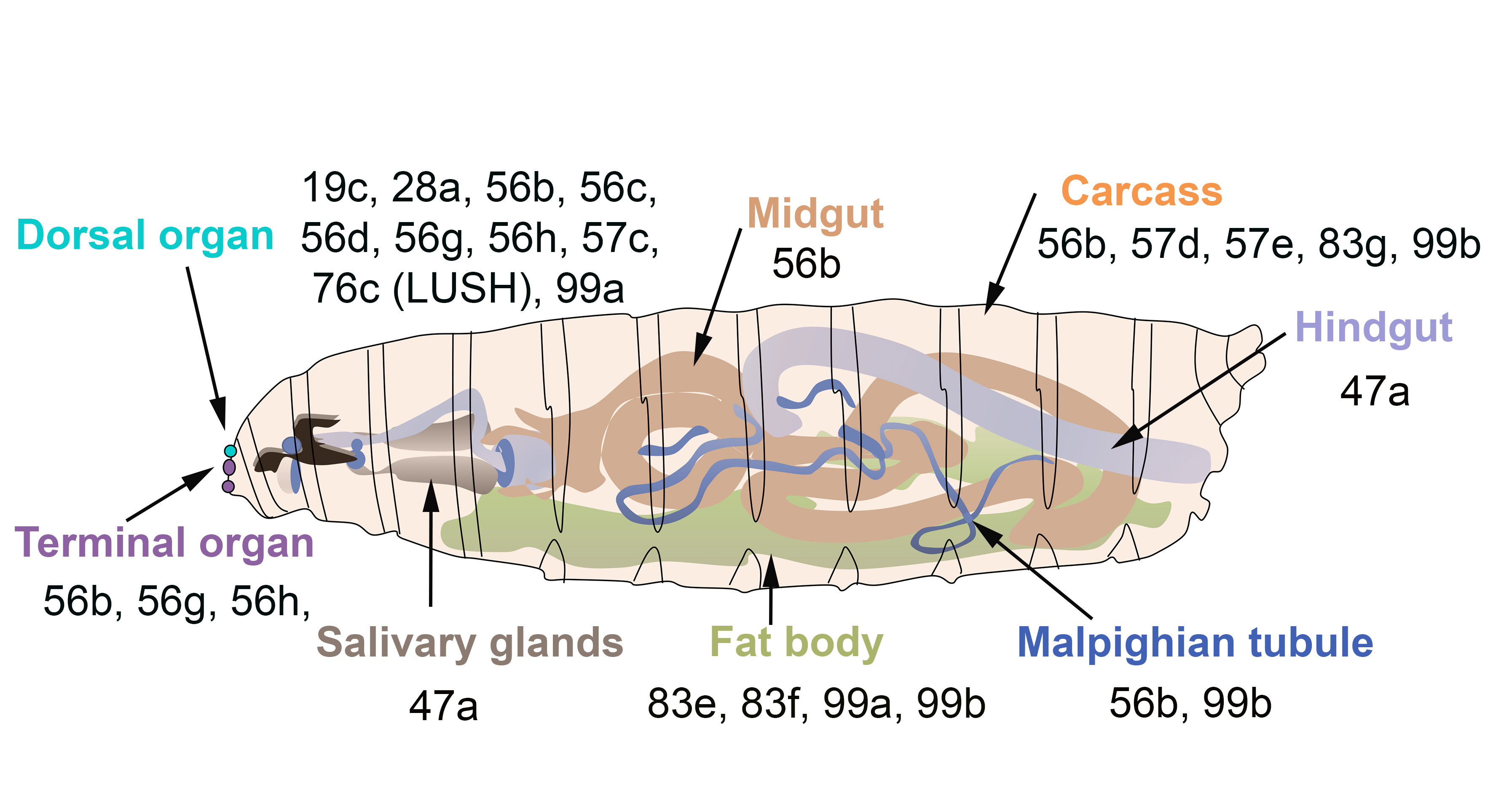
Figure 3. OBPs expressed in the olfactory gustatory and digestive organs of D. melanogaster larvae. The larval dorsal organ (turquoise) detects volatile odorants, while the larval terminal organ (purple) detects both soluble and volatile chemicals.
Surprisingly, several OBPs are expressed in the venom glands of the parasitic wasps
Leptopilina heterotoma and
Pteromalus puparum and of the honeybee
A. mellifera [
73,
74,
75]. Few studies have reported the expression of OBPs in the insect digestive tract. For instance, PregOBP56a was detected in the oral disk of the blowfly
Phormia regina [
76], while OBP56d was identified in the hindgut of
D. melanogaster flies [
54]. Fluorescent binding assays revealed that PregOBP56a binds palmitic, stearic, oleic, and linoleic acids. These data indicate that PregOBP56a might solubilize and deliver fatty acids to the midgut during feeding [
76]. Similarly, the midgut of
Rhodnius prolixus also expresses OBPs [
77]. Other studies have shown that the expression of OBPs can be altered depending on the insect’s diet. Indeed, the expression of one OBP of female
Culex nigripalpus increased in the midgut, thorax and abdomen after a bloodmeal, suggesting a possible role in blood feeding [
78,
79]. Moreover, a diet change in
Anoplophora glabripennis can affect gut-expressed OBPs together with other genes implicated in digestion, detoxification and nutrient acquisition. The feeding of
A. gabripennis larvae in a host with documented resistance (
Populus tomentosa) induced the downregulation of 5 OBP genes. It is not known whether alteration of the gut OBP gene expression is directly linked to the resistance of
A. gabripennis to the
Populus tomentosa plant [
80]. Moreover, bacterial symbionts increase the gut expression of tsetse’s OBP6 and of OBP28a in
D. melanogaster [
81]. The roles of these OBPs are discussed in a later section of this review. Notably, nineteen
B. dorsalis OBPs and seven
D. melanogaster OBPs are highly expressed in the fat body [
41] (c), although their roles in the fat body remain unclear. It is important to acknowledge that the expression of OBPs in specific organs does not represent a proof of function. Further physiological studies are needed to fully investigate the role(s) of OBPs in the different parts of the insect body. In the absence of selection against OBPs expression, some OBPs still become expressed despite having no obvious function. This phenomenon could lead to rapid evolution of novel functions.
3. Biochemical Properties of OBPs
The structure and binding properties of Insect OBPs were investigated. Insect OBPs are small soluble proteins classified based on the number of amino acid residues found in their primary structure: long-chain OBPs (~160 residues), medium-chain OBPs (~120 residues) and short-chain OBPs (~100 residues). The similarity between the amino acid sequences of OBPs from the same species is low (<10% identity). The protein sequences of insect OBPs include highly conserved cysteines with a specific number of amino acid residues (AAs) between them [82,83].
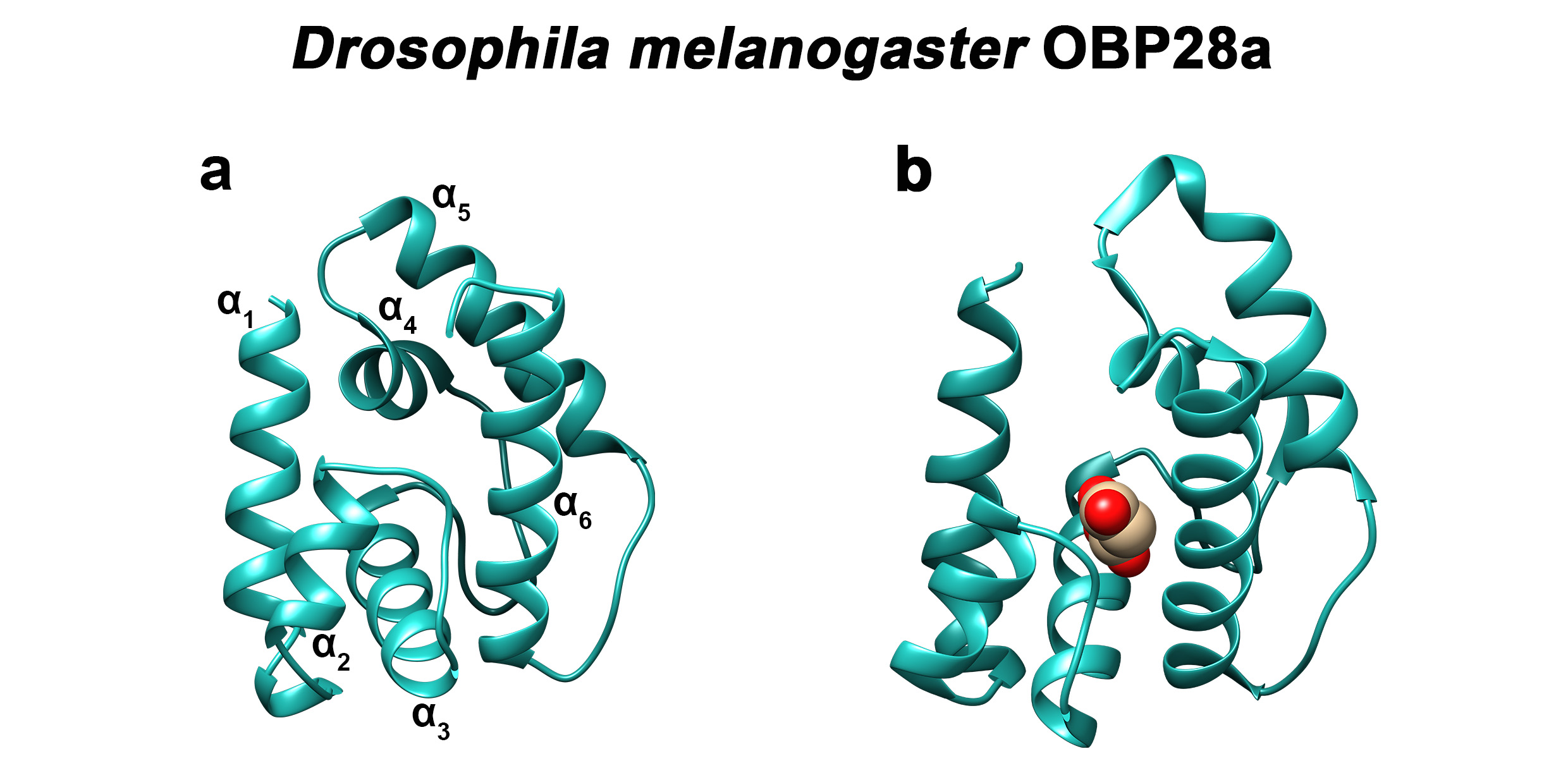
The three-dimensional structure of classic OBPs consists of a six α-helical domain forming a hydrophobic cavity [94] (Figure 4). The structural stability of insect OBPs depends on the presence of three interlocked disulphide bridges linking conserved cys- teines [95,96,97]. Although the AA sequences of insect OBPs are highly divergent between and within species, the structure of insect OBPs is highly conserved. To date, crystal or NMR structures of more than 20 OBPs or PBPs from species belonging to different insect orders have been solved and are available in Entrez’s 3-D structure database at NCBI, ac- companied by more than 20 detailed papers [94,98,99,100,101,102,103,104,105]. OBP structures have been solved in ligand-free (apo) or ligand-bound states, allowing researchers to study the interaction of the binding cavity with pheromones or with general odorants (Figure 4).
Figure 4. Three-dimensional structure of D. melanogaster OBP28a. (a) Apo- and (b) bound-three-dimensional structures of the fruit fly (Drosophila melanogaster) OBP28a (PDB: 6QQ4). α1 to α6 indicate the six α-helices found in the structure of classical insect OBPs. Drosophila OBP28a is in complex with penta-ethylene glycol [94].
The potential ligands of insect OBPs have been identified using different techniques. These studies revealed that OBPs can bind different families of chemical compounds. The physiological role of OBPs in the perception of the identified ligands was investigated in further studies using behavioural assays and electrophysiology.
4. Diverse Chemosensory Functions of OBPs
Since the discovery of OBPs, several hypotheses and models have been proposed concerning their roles in chemoreception [125]. Later, studies using structural analyses, in vitro binding, behavioural assays and electrophysiological recordings revealed unsuspected roles of insect OBPs (see Table 2).
|
OBP
|
Role
|
Publication
|
|
OBP76a (LUSH)
|
Solubilization, transport and interaction with SNMP1
|
[111,122,134]
|
|
OBP69a
|
Implication in cVA response, role remains unclear
|
[140]
|
|
OBP28a
|
Modulation of olfactory sensitivity
|
[34,94]
|
|
OBP59a
|
Humidity detection
|
[57]
|
|
OBP57d
and OBP57e
|
Modulation of oviposition site preference to C6-C9 acids in D. melanogaster and D. sechellia
Specialization of D. sechellia to its host plant (Tahitian Noni)
|
[151]
[152]
|
|
OBP49a
|
Suppression of the appetence for sweet compounds through the perception of bitter chemicals
|
[36]
|
|
OBP56h
|
Modulation of mating behaviour by alteration of cuticular hydrocarbon profiles in males
|
[142]
|
|
OBP19b
|
Detection of peculiar amino acids
|
[37]
|
Conclusions
(1) Studies conducted in the last 40 years have provided information regarding the different roles of insect OBPs. OBPs were believed to be only expressed in olfactory organs and to be strictly involved in chemoreception mechanisms. However, an increasing number of reports has revealed that OBPs are expressed in most organs of the insect body and have non-conventional roles, including in taste, immunity response and humidity detection.
(2) We present the latest discoveries made on the structural and binding properties of insect OBPs. We focus on the properties of insect OBPs and, more specifically, on their tissue and cellular expression. We also present the varied functional roles, both classical and some non-conventional, of currently known OBPs.
(3) Yet there is massive amount of information on OBP functions that we ignore and needs to be investigated. These studies will pave the way to different technological applications in environmental, food quality and medical fields.