Botryosphaeriaceae (Botryosphaeriales) include several species reported as endophytes, latent, and woody plant pathogens on a broad range of host. The most common symptoms observed in association with species of Botryosphaeriaceae are twig, branch and trunk cankers, die-back, collar rot, root cankers, gummosis, decline and, in severe cases, plant death.
- Diplodia
- Dothiorella
- Lasiodiplodia
- Neofusicoccum
- pathogenic fungi
- phylogeny
1. Introduction
Citrus production represents one of the most important fruit industries worldwide in terms of total yield. Greece, Italy, Portugal, and Spain are the most important European producers of citrus fruit [1]. In 2019, nearly 11 million tons of citrus was produced in Europe on approximately 515,000 ha [2]. Most canker diseases of citrus, as well as further fruit-tree crops, are caused by a broad range of fungal species that infect the wood mainly through winter pruning wounds and a subsequent colonization of vascular tissues [3]. Several abiotic and biotic factors are considered responsible for rots and gumming on the trunk and main branches in citrus. Frost damage, sunscald, or water distribution can promote the infection of numerous ascomycetes and basidiomycetes [4]. Several fungal infections involving twigs, branches and trunks of citrus caused by Colletotrichum and Diaporthe species were reported in different continents [5][6][7][8][9]. Guarnaccia and Crous [10] reported serious cankers developing in woody tissues of lemon trees caused by Diaporthe spp., often with a gummose exudate, causing serious blight and dieback. Canker diseases of citrus are also caused by other fungal genera such as Fusarium and Neocosmospora [11], Peroneutypa [12][13], and Phaeoacremonium [14]. Recently, significant attention has been dedicated to revising species and genera of Botryosphaeriaceae, which encompass species with a cosmopolitan distribution that are able to cause diseases of numerous plant species worldwide [15][16].
Botryosphaeriaceae (Botryosphaeriales) include several species reported as endophytes, latent, and woody plant pathogens on a broad range of host [15][16][17]. This family has undergone significant revision after the adoption of molecular tools to resolve its taxonomy [15][16][18][19][20][21][22][23]. Recently, the taxonomy of Botryosphaeriaceae (and other families in Botryosphaeriales) has been reviewed by Phillips et al. [23] based on morphology of the sexual morphs, phylogenetic relationships on internal transcribed spacer (ITS) and 28S large subunit (LSU) of nuclear ribosomal DNA (nrDNA) sequences and evolutionary divergence times. The authors highlighted the main findings made by Yang et al. [16] who included new families, genera, and species in Botryosphaeriales based on morphology and multi-marker phylogenetic analyses of a large collection of isolates. Currently, six families are accepted in Botryosphaeriales and 22 genera have been included in Botryosphaeriaceae [23][24][25].
The most common symptoms observed in association with species of Botryosphaeriaceae are twig, branch and trunk cankers, die-back, collar rot, root cankers, gummosis, decline and, in severe cases, plant death [15][17]. Plant infections mainly occur through natural openings or wounds, but these fungal species could also survive in latency. This ability could lead to their spread worldwide through asymptomatic plant material, seedlings and fruit, frequently circumventing the adopted quarantine measures [22]. Moreover, stress and non-optimal plant growth conditions consistently induce the expression of diseases associated with Botryosphaeriaceae species. Thus, global warming could increase plant stress and induce favourable conditions for the development of Botryosphaeriaceae diseases [17][26][27]. Species within the Botryosphaeriaceae represent a serious threat to different crops including major fruit, berry fruit and nut crops cultivated in sub-tropical, tropical, or temperate areas [22][28][29][30].
Several species of Diplodia (Di.), Dothiorella (Do.), Lasiodiplodia, Neofusicoccum, and Neoscytalidium (Ne.) have been previously reported to affect Citrus species [13][31][32][33]. For example, Ne. dimidiatum has been reported causing citrus branch canker in California [13] and Italy [32]; Do. viticola, L. citricola, L. theobromae, and Ne. dimidiatum have been described in association with branch and trunk dieback of citrus trees in Iran [14][34] and Dothiorella spp. have been detected as causal agents of citrus gummosis in Tunisia [35]. Moreover, Di. seriata, Di. mutila, Do. viticola, L. mediterranea and L. mitidjana, have been recovered from symptomatic citrus trees in Algeria [33].
2. Field Sampling and Fungal Isolation
In this study, the sampling focused on symptomatic plants of Citrus limon, C. reticulata, C. sinensis, C. sinensis × Poncirus trifoliata, and Microcitrus australasica. Samples were collected in 19 orchards. Citrus trees showed various external disease symptoms, including partial or complete yellowing, wilting leaves and twigs, and dieback of branch tips, but also defoliation and branch decline. Canker and cracking of the bark associated with gummose exudate occurred on trunks and branches. Internal observation of infected branches revealed black to brown wood discoloration in cross-sections, wedge-shaped necrosis or irregular wood discoloration. Twigs were wilted and occasionally presenting sporocarps (Figure 1). Symptoms were detected in all the orchards and regions investigated. A total of 63 fungal isolates were collected and were found to be characterized by dark green to grey, fast-growing mycelium on MEA. Moreover, the isolates produced pycnidia on pine needles within 40 days, containing pigmented or hyaline conidia. According to these characteristics, the fungal isolates were classified as Botryosphaeriaceae spp. based on comparison with the previous generic descriptions [15]. Among the collected isolates, 18 were obtained from trunk cankers, 10 were associated with branch infections, and 35 from twig dieback.

Figure 1. Symptoms on citrus tissues with associated Botryosphaeriacae species. (A) Branch decline in commercial lemon orchard. (B) Trunk canker and bark cracking of C. sinensis. (C,D) Trunk and branch canker with gummosis of C. sinensis plants. (E,F) External cracking with gummosis and internal wood discoloration of the same affected branch of C. reticulata plant. (G,H) Internal wood discoloration and branch blight of C. limon. (I) Twig dieback of young C. sinensis × P. trifoliata and M. australasica (J) plants.
3. Phylogenetic Analyses
A combined multi-marker (ITS, TEF1, and TUB2) phylogenetic tree was inferred for each genus (Diplodia, Dothiorella, Lasiodiplodia, and Neofusicoccum) obtained in this study (Figure 2, Figure 3, Figure 4 and Figure 5). The best nucleotide models for the Bayesian Inference analysis of each dataset were as follows: SYM (symmetrical model) + I (proportion of invariable sites) + G (gamma distribution) (Diplodia, Dothiorella, Lasiodiplodia, and Neofusicoccum) for ITS; GTR (generalized time-reversible model) + G (Diplodia, Dothiorella and Neofusicoccum) and HKY (Hasegawa–Kishino–Yano) + I + G (Lasiodiplodia) for TEF1 and GTR + G (Diplodia, Lasiodiplodia and Neofusicoccum) and GTR + I + G (Dothiorella) for TUB2. The Diplodia phylogenetic analysis revealed the isolates as belonging to Di. pseudoseriata (15 isolates, BPP = 1 and ML-BS = 100), Di. seriata (9 isolates, BPP = 1 and ML-BS = 95), Di. olivarum (2 isolates, Bayesian posterior probabilities (BPP) = 1 and maximum likelihood bootstrapped (ML-BS) = 99), and Di. mutila (1 isolate, BPP = 0.99 and ML-BS = 87) (Figure 2). The Dothiorella phylogeny (Figure 3) grouped the isolates together within Do. viticola (9 isolates, BPP = 1 and ML-BS = 99). The Lasiodiplodia phylogenetic analysis placed five isolates as L. theobromae (BPP = 1 and ML-BS = 98) (Figure 4). The Neofusicoccum phylogeny (Figure 5) grouped sequences from our isolates as belonging to N. luteum (2 isolates, BPP = 1 and ML-BS = 94), N. parvum (16 isolates) and N. mediterraneum (4 isolates, BPP = 1 and ML-BS = 98).
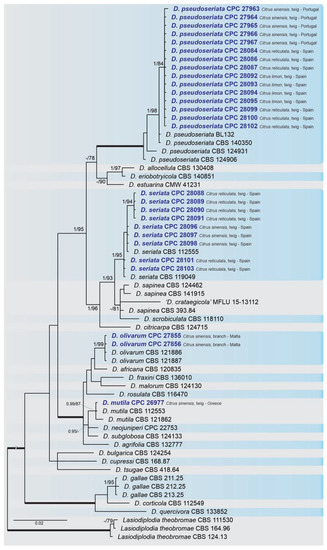
Figure 2. Bayesian inference analysis of Diplodia species using ITS rDNA, TEF1 and TUB2 sequences. Isolates obtained in this study are in bold and blue. Bayesian posterior probability (BPP) and maximum likelihood-bootstrap (ML-BS) values equal or greater than 0.95 and 70%, respectively, are shown near nodes. Thickened branches represent clades with ML-BS = 100% and a BPP = 1.0. The tree was rooted to L. theobormae (CBS 111530, CBS 164.96 and CBS 124.13).
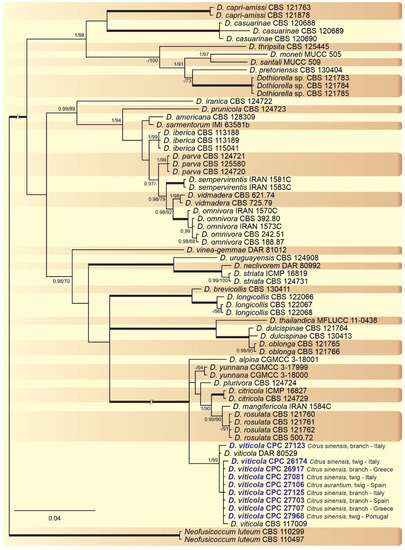
Figure 3. Bayesian inference analysis of Dothiorella species using ITS rDNA, TEF1, and TUB2 sequences. Isolates obtained in this study are in bold and blue. Bayesian posterior probability (BPP) and ML bootstrap (ML-BS) values equal or greater than 0.95 and 70%, respectively, are shown near nodes. Thickened branches represent clades with ML-BS = 100% and a BPP = 1.0. The tree was rooted to N. luteum (CBS 110299 and CBS 110497).
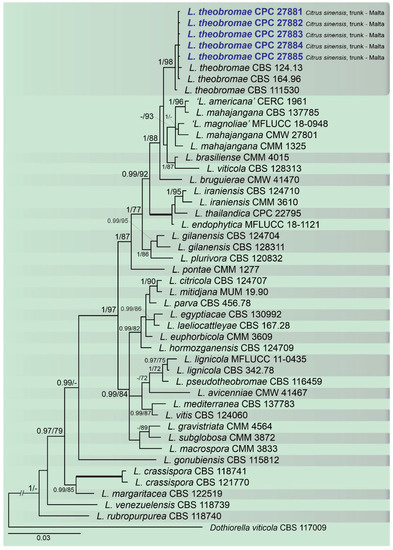
Figure 4. Bayesian inference analysis of Lasiodiplodia species using ITS rDNA, TEF1, and TUB2 sequences. Isolates obtained in this study are in bold and blue. Bayesian posterior probability (BPP) and ML bootstrap (ML-BS) values equal or greater than 0.95 and 70%, respectively, are shown near nodes. Thickened branches represent clades with ML-BS = 100% and a BPP = 1.0. The tree was rooted to Do. viticola (CBS 117009).
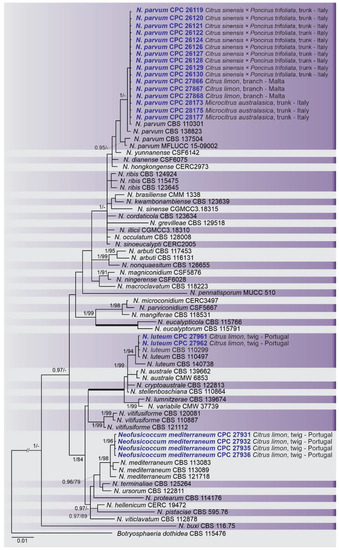
Figure 5. Bayesian inference analysis of species Neofusicoccum using ITS rDNA, TEF1, and TUB2 sequences. Isolates obtained in this study are in bold and blue. Bayesian posterior probability (BPP) and ML bootstrap (ML-BS) values equal or greater than 0.95 and 70%, respectively, are shown near nodes. Thickened branches represent clades with ML-BS = 100% and a BPP = 1.0. The tree was rooted to B. dothidea (CBS 115476).
4. Occurrence of Botryosphaeriaceae among Countries and Citrus Species
Among countries, Do. viticola was found in Greece, Italy, Portugal, and Spain; N. parvum in Italy and Malta, and Di. pseudoseriata in Portugal and Spain. In addition, Di. mutila and Di. seriata were exclusively isolated in Greece and Spain, respectively; L. theobromae and Di. olivarum were only found in Malta, and N. luteum and N. mediterraneum were exclusively found in Portugal. Based on the citrus species, N. parvum (25.4%) and Di. pseudoseriata (23.8%) were the most frequently detected Botryosphaeriaceae spp. on C. sinensis × P. trifoliata, C. limon, C. reticulata, C. sinensis, and/or M. australasica; Di. seriata (on C. reticulata and C. sinensis); and Do. viticola (on C. aurantium and C. sinensis) had an equal percentage of frequency (14.3%); Di. mutila (exclusively found on C. sinensis), N. luteum and N. mediterraneum (only found on C. limon) and Di. olivarum and L. theobromae (exclusively found on C. sinensis) had low frequency values varying from 1.6% to 7.9%.
5. Discussion
Several Botryosphaeriaceae spp. have been detected in association with citrus cankers worldwide. Diplodia seriata, Di. mutila, Do. iberica, Do. viticola, L. parva, N. australe, N. luteum, N. mediterraneum, N. parvum, and Ne. dimidiatum have been recovered from necrotic tissues of branch canker and rootstock citrus samples in California [13][31][36]. Recently, Di. citricarpa was described for a fungus on twigs of Citrus sp. in Iran [16] and L. mitidjana was introduced for a fungus causing branch canker and dieback of C. sinensis in Algeria [33]. Botryosphaeriaceae spp. causing disease on citrus are known in European countries, where N. parvum and Ne. dimidiatum were reported on C. reticulata in Greece and on C. sinensis in Italy, respectively [32][37].
This study represents the first large survey aimed at studying the occurrence, genetic diversity, and pathogenicity of Botryosphaeriaceae species associated with symptomatic citrus species of citrus-producing areas in Greece, Italy, Portugal, Malta, and Spain [10][38]. Results obtained during our study have added new information about the pathogenicity of Botryosphaeriaceae spp. in citrus-producing areas of these European countries. Symptomatic plants were observed during fieldwork in all the citrus orchards and regions investigated and all isolates used in the pathogenicity test caused lesions on wood of inoculated citrus plants. Phylogenetic multi-marker analyses recognized botryosphaeriaceous isolates in four Diplodia species, with Di. pseudoseriata (15 isolates) being the most common; followed by three Neofusicoccum species, with N. parvum (16 isolates) as dominant species, Do. viticola (9 isolates), and L. theobromae (5 isolates). All species found in this study, except Di. pseudoseriata and Di. olivarum, which are reported for the first time on Citrus spp., have been found in citrus-producing areas of California (USA) [13][31][36].
Diplodia and Neofusicoccum species were dominant in this study. Different species of Neofusicoccum and Diplodia were the most frequently detected pathogens causing gummosis on citrus in California [36] and Di. citricarpa was a new species isolated from Citrus sp. in Iran [16]. Species of Diplodia, Dothiorella, Lasiodiplodia, and Neofusicoccum detected in our study are widely reported as pathogens of other host plants in Algeria and Tunisia [39][40], Australia [41], Brazil [42], China [43][44], Chile [45], Italy, Portugal [39][46][47][48], South Africa [49], and the USA [13][31][36]. The results obtained in our study provide valuable information related to the richness, occurrence, and pathogenicity of Botryosphaeriaceae species in association with citrus species. This study is also the first major survey for Botryosphaeriaceae species associated with symptomatic citrus species in citrus-producing areas of five European countries, providing essential information for future monitoring. Moreover, while previous reports of canker diseases of citrus were based exclusively on morphological observations, the current study aimed to investigate the fungi affecting the major citrus production areas in Europe by large-scale sampling, morphology, and DNA phylogeny. The information achieved with this study about Botryosphaeriaceae population and citrus canker etiology provide fundamental knowledge to start further studies aimed to improve the disease management.
This entry is adapted from the peer-reviewed paper 10.3390/plants10030492
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2019. Available online: (accessed on 26 February 2020).
- Eurostat. Citrus Fruit Statistics; Eurostat: Luxembourg, 2020.
- Crous, P.W.; Wingfield, M.J. Fungi infecting woody plants: Emerging frontiers. Persoonia 2018, 40, i–iii.
- Fawcett, H.S. Citrus Diseases and Their Control; McGraw-Hill: New York, NY, USA, 1936.
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74.
- Huang, F.; Hou, X.; Dewdney, M.M.; Fu, Y.S.; Chen, G.Q.; Hyde, K.D.; Li, H. Diaporthe species occurring on citrus in China. Fungal Divers. 2013, 61, 237–250.
- Mahadevakumar, S.; Yadav, V.; Tejaswini, G.S.; Sandeep, S.N.; Janardhana, G.R. First report of Phomopsis citri associated with dieback of Citrus lemon in India. Plant Dis. 2014, 98, 1281.
- Guarnaccia, V.; Crous, P.W. Species of Diaporthe on Camellia and Citrus in the Azores Islands. Phytopathol. Mediterr. 2018, 57, 307–319.
- Mayorquin, J.S.; Nouri, M.T.; Peacock, B.B.; Trouillas, F.P.; Douhan, G.W.; Kallsen, C.; Eskalen, A. Identification, Pathogenicity, and Spore Trapping of Colletotrichum karstii Associated with Twig and Shoot Dieback in California. Plant Dis. 2019, 103, 1464–1473.
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334.
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P.W. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia 2018, 40, 1–25.
- Timmer, L.W.; Garnsey, S.M.; Graham, J.H. Compendium of Citrus Diseases, 2nd ed.; American Phytopathological Society: Saint Paul, MN, USA, 2000.
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with Citrus branch canker in the southern California desert. Plant Dis. 2016, 100, 2402–2413.
- Espargham, N.; Mohammadi, H.; Gramaje, D.A. Survey of Trunk Disease Pathogens within Citrus Trees in Iran. Plants 2020, 9, 754.
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167.
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346.
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106.
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253.
- Crous, P.W.; Giraldo, A.; Hawksworth, D.L.; Robert, V.; Kirk, P.M.; Guarro, J.; Robbertse, B.; Schoch, C.L.; Damm, U.; Trakunyingcharoen, T.; et al. The genera of fungi: Fixing the application of type species of generic names. IMA Fungus 2014, 5, 141–160.
- Crous, P.W.; Muller, M.M.; Sanchez, R.M.; Giordano, L.; Bianchinotti, M.V.; Anderson, F.E.; Groenewald, J.Z. Resolving Tiarosporella spp. allied to Botryosphaeriaceae and Phacidiaceae. Phytotaxa 2015, 202, 73–93.
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052.
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49.
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22.
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456.
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115.
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Cankers and other diseases caused by the Botryosphaeriaceae. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CAB International: Boston, MA, USA, 2013; pp. 298–317.
- Pour, F.N.; Ferreira, V.; Félix, C.; Serôdio, J.; Alves, A.; Duarte, A.S.; Esteves, A.C. Effect of temperature on the phytotoxicity and cytotoxicity of Botryosphaeriaceae fungi. Fungal Biol. 2020, 124, 571–578.
- Guarnaccia, V.; Martino, I.; Tabone, G.; Brondino, L.; Gullino, M.L. Fungal pathogens associated with stem blight and dieback of blueberry in northern Italy. Phytopathol. Mediterr. 2020, 59, 229–245.
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488.
- Aiello, D.; Gusella, G.; Fiorenza, A.; Guarnaccia, V.; Polizzi, G. Identification of Neofusicoccum parvum causing canker and twig blight on Ficus carica in Italy. Phytopathol. Mediterr. 2020, 59, 213–218.
- Adesemoye, A.O.; Eskalen, A. First Report of Spencermartinsia viticola, Neofusicoccum australe, and N. parvum causing branch canker of citrus in California. Plant Dis. 2011, 95, 770.
- Polizzi, G.; Aiello, D.; Vitale, A.; Giuffrida, F.; Groenewald, J.; Crous, P.W. First report of shoot blight, canker, and gummosis caused by Neoscytalidium dimidiatum on citrus in Italy. Plant Dis. 2009, 93, 1215.
- Berraf-Tebbal, A.; Mahamedi, A.E.; Aigoun-Mouhous, W.; Špetík, M.; Čechová, J.; Pokluda, R.; Baránek, M.; Eichmeier, A.; Alves, A. Lasiodiplodia mitidjana sp. nov. and other Botryosphaeriaceae species causing branch canker and dieback of Citrus sinensis in Algeria. PLoS ONE 2020, 15, e0232448.
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1–10.
- Hamrouni, N.; Nouri, M.; Trouillas, F.; Said, A.; Sadfi-Zouaoui, N.; Hajlaoui, M. Dothiorella gummosis caused by Dothiorella viticola, first record from citrus in Tunisia. New Dis. Rep. 2018, 38, 10.
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of species of Botryosphaeriaceae causing bot gummosis in citrus in California. Plant Dis. 2014, 98, 55–61.
- Vakalounakis, D.J.; Ntougias, S.; Kavroulakis, N.; Protopapadakis, E. Neofusicoccum parvum and Diaporthe foeniculina associated with twig and shoot blight and branch canker of citrus in Greece. J. Phytopathol. 2019, 167, 527–537.
- Guarnaccia, V.; Groenewald, J.Z.; Li, H.; Glienke, C.; Carstens, E.; Hattingh, V.; Crous, P.W. First report of Phyllosticta citricarpa and description of two new species, P. paracapitalensis and P. paracitricarpa, from citrus in Europe. Stud. Mycol. 2017, 87, 161–185.
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Boutiti, M.Z.; Jamâa, M.L.B.; Phillips, A.J.L. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214.
- Mahamedi, A.E.; Phillips, A.J.L.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, distribution and host association of Botryosphaeriaceae species causing oak decline across different forest ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765.
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q. Current status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2018, 48, 35–44.
- Machado, A.R.; Custódio, F.A.; Cabral, P.G.C.; Capucho, A.S.; Pereira, O.L. Botryosphaeriaceae species causing dieback on Annonaceae in Brazil. Plant Pathol. 2019, 68, 1394–1406.
- Xu, C.; Zhang, H.; Zhou, Z.; Hu, T.; Wang, S.; Wang, Y.; Cao, K. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752.
- Li, G.; Slippers, B.; Wingfield, M.J.; Chen, S. Variation in Botryosphaeriaceae from Eucalyptus plantations in YunNan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 22.
- Valencia, A.L.; Pilar, M.; Gil, B.A.; Latorre, I.; Rosales, M. Characterization and Pathogenicity of Botryosphaeriaceae Species Obtained from Avocado Trees with Branch Canker and Dieback and from Avocado Fruit with Stem End Rot in Chile. Plant Dis. 2019, 103, 996–1005.
- Gusella, G.; Aiello, D.; Polizzi, G. First report of leaf and twig blight of Indian hawthorn (Rhaphiolepis indica) caused by Neofusicoccum parvum in Italy. J. Plant Pathol. 2020, 102, 275.
- Alves, A.; Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Phillips, A.J.L. The complex of Diplodia species associated with Fraxinus and some other woody hosts in Italy and Portugal. Fungal Divers. 2014, 67, 143–156.
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Yan, J.Y.; Li, X.H. Saprobic Botryosphaeriaceae, including Dothiorella italica sp nov., associated with urban and forest trees in Italy. Mycosphere 2017, 8, 1157–1176.
- Jami, F.; Slippers, B.; Wingfield, M.J.; Loots, M.T.; Gryzenhout, M. Temporal and spatial variation of Botryosphaeriaceae associated with Acacia karroo in South Africa. Fungal Ecol. 2015, 15, 51–62.