Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Endocrinology & Metabolism
Plant-beneficial Pseudomonas spp. aggressively colonize the rhizosphere and produce numerous secondary metabolites, such as 2,4-diacetylphloroglucinol (DAPG). DAPG is a phloroglucinol derivative that contributes to disease suppression, thanks to its broad-spectrum antimicrobial activity. A famous example of this biocontrol activity has been previously described in the context of wheat monoculture where a decline in take-all disease (caused by the ascomycete Gaeumannomyces tritici) has been shown to be associated with rhizosphere colonization by DAPG-producing Pseudomonas spp.
- 2
- 4-diacetylphloroglucinol
- DAPG
- Pseudomonas
- biocontrol
- antibiotic
1. Introduction
Phloroglucinol derivatives are a large class of secondary metabolites widely distributed in plants and brown algae. Over a thousand phloroglucinol derivatives have been characterized to date. As an example, 429 phloroglucinol derivatives have been isolated from the genus Hypericum alone [1]. Phloroglucinol derivatives found in plants and brown algae have extremely diverse structures, ranging from the simple grandinol, an acylphloroglucinol produced by several Eucalyptus species, to the more complex phlorotannins found in several families of brown algae [2,3]. These compounds often exhibit antiviral, antibacterial and antifungal activity [2]. Phloroglucinol derivatives are also produced by some microorganisms [2,4]. By contrast with the phloroglucinol derivatives found in plants and brown algae, phloroglucinol derivatives of microbial origin are rather simple. Some Pseudomonas strains produce 2,4-diacetylphlorglucinol (DAPG) alongside its biosynthetic intermediates monoacetylphloroglucinol (MAPG) and phloroglucinol.
DAPG-producing Pseudomonas spp. have received particular attention due to their ability to control numerous soil-borne plant diseases, including take-all of wheat, tobacco black root rot and sugar beet damping-off [4,5]. These bacteria also play an important role in natural disease suppressiveness found in several soils across the world. Besides their presence in the rhizosphere, DAPG-producing Pseudomonas spp. are also known to colonize various environment, including the phyllosphere [6], the skin surface of certain amphibians [7] and the surface of marine algae [8]. This review specifically covers rhizosphere-inhabiting DAPG-producing Pseudomonas spp.
Several reviews have been previously published on rhizosphere-inhabiting DAPG-producing Pseudomonas spp. and their role in take-all decline [4,9].
2. Genetics, Biochemistry, and Evolution of DAPG Biosynthesis
2.1. The Phl Biosynthetic Gene Cluster
Genes involved in DAPG biosynthesis were cloned several times from three different DAPG-producing Pseudomonas strains: Pseudomonas sp. Q2-87 [10,11], P. kilonensis F113 [12] and P. protegens CHA0 [13]. Further characterization of the genomic fragment isolated from Pseudomonas sp. Q2-87 led to the description of the so-called phl biosynthetic gene cluster (BCG) [14]. Six genes were originally described: four were found to be directly involved in DAPG biosynthesis (phlABCD) and the two others were shown to encode a putative permease (phlE) and a TetR regulatory protein (phlF). Three other genes were later discovered and associated with the BCG: phlG, which encodes a hydrolase involved in DAPG degradation [15], phlH, which encodes another TetR regulatory protein [15] and phlI, which encodes an uncharacterized protein [16,17]. The organization of the phl BCG is conserved in all DAPG-producing Pseudomonas spp. sequenced to date [18]. The biosynthetic cluster and the current understanding of the DAPG biosynthesis pathway are presented in Figure 1.
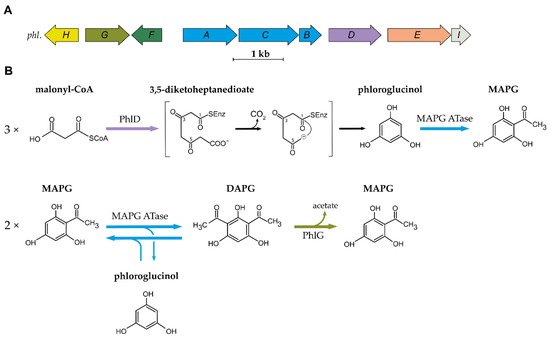
Figure 1. Organization of the 2,4-DAPG biosynthetic cluster and current understanding of the biosynthesis scheme. (A) Organization of the phl biosynthetic gene cluster found in DAPG-producing Pseudomonas spp. (B) Current understanding of the biosynthesis and degradation of DAPG in the genus Pseudomonas. MAPG ATase is an enzyme multiplex composed of PhlA, PhlB and PhlC units. Abbreviations are as follows: MAPG, monoacetylphloroglucinol; DAPG, 2,4-diacetylphloroglucinol.
2.2. Biosynthesis and Degradation of DAPG
The first step in the biosynthesis of DAPG is catalysed by the type III polyketide synthase (PKS) PhlD [14,19]. Type III PKSs are homodimeric enzymes that catalyse the iterative condensation of a starter substrate (usually an acyl-CoA) with several extender substrate units (usually malonyl-CoA) to generate a linear polyketide, which is subsequently cyclized [20]. Bangera and Thomashow [14] proposed that PhlD uses acetoacetyl-CoA as the starter substrate to produce monoacetylphloroglucinol (MAPG), but it was later showed that PhlD produces phloroglucinol from malonyl-CoA instead [19]. While PhlD uses malonyl-CoA as a preferred substrate, it also accepts other starter substrates with an aliphatic chain of C4-C12 in vitro [21]. The proposed mechanism leading to the formation of phloroglucinol is that PhlD catalyses the iterative condensation of three molecules of malonyl-CoA into 3,5-diketoheptanedioate [19]. This polyketide intermediate undergoes decarboxylation and is subsequently cyclized via a Claisen condensation, leading to the formation of phloroglucinol [19,21].
In the following steps, acetylation of phloroglucinol leads to the formation of MAPG, which is subsequently acetylated into DAPG. MAPG was identified as a putative intermediate in DAPG biosynthesis by Shanahan and colleagues [22], who also found experimental evidence for an enzymatic acetyltransferase activity in cell-free extracts of P. kilonensis F113. Bangera and Thomashow [14] found that phlACB was required for the biosynthesis of MAPG and DAPG, suggesting a role for PhlACB in the production and the acetylation of MAPG. Achkar and colleagues [19] confirmed that the product of phlACB catalyses the acetylation of phloroglucinol and MAPG: The addition of phloroglucinol to the culture medium of an E. coli strain carrying plasmid-localized phlACB led to the formation of MAPG and DAPG. Later, Hayashi and colleagues [17] characterized the multimeric enzyme composed of PhlA, PhlC and PhlB units, which was named MAPG acetyltransferase (MAPG ATase). This enzyme, unlike most acetyltransferases described so far, was shown to catalyse C-C bond formation without the use of CoA-activated substrates [17]. The MAPG ATase catalyses the disproportionation of two molecules of MAPG, resulting in the formation of DAPG and the production of phloroglucinol.
The MAPG ATase also catalyses the reverse reaction, yielding two molecules of MAPG from a molecule of DAPG and a molecule of phloroglucinol [17]. This results in an equilibrium where DAPG, MAPG and phloroglucinol are present at quasi-equimolar concentration. Recent studies have provided new insight into the catalytic properties, the mechanism, and the structure of the MAPG ATase [23,24,25,26,27]. This enzyme was shown to use various non-natural substrates as acyl-donor in in vitro experiments [26,27,28]. This is particularly interesting given the fact that the acyl donor involved in MAPG biosynthesis from phloroglucinol remains to be characterized. The crystal structure revealed that PhlACB subunits are arranged in a Phl(A2C2)2B4 composition where four PhlB units mediate the binding of two PhlA and two PhlC dimers [23]. Crystal soaking and site-directed mutagenesis experiments suggest that only PhlC units are involved in the acyl transfer reaction [23].
DAPG is degraded by the zinc-dependent hydrolase PhlG [29,30]. PhlG degrades DAPG into MAPG and acetate by cleaving one of the C-C bonds linking the acetyl groups to the phenolic ring [29,30]. This enzyme is highly specific for its substrate DAPG, as it is unable to degrade structurally similar compounds, such as MAPG or triacetylphloroglucinol [29]. The crystal structure of PhlG revealed that it cleaves C-C bonds using a Bet v1-like fold domain, contrary to the alpha/beta fold classically used by hydrolases [30].
2.3. Distribution and Evolution of the Phl Biosynthetic Gene Cluster
The phl BCG is mainly found in the P. corrugata and P. protegens subgroups of the P. fluorescens species complex [18], as shown in Figure 2. The phl BCG is not present in all the strains belonging to these two subgroups, and its distribution in these two subgroups is patchy [18,31,32].
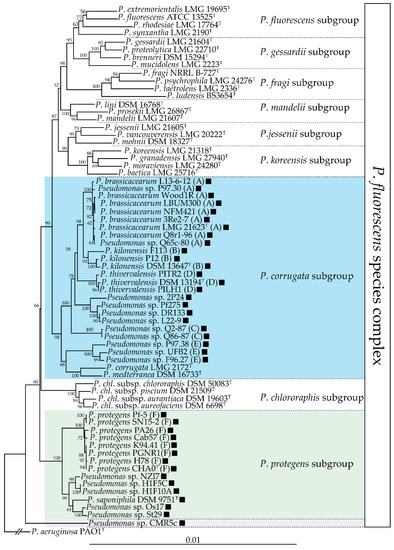
Figure 2. Distribution of the phl biosynthetic gene cluster in the P. fluorescens species complex. This neighbour-joining phylogeny is based on an alignment of the concatenated partial sequences of four housekeeping genes (16s rDNA, gyrB, rpoB, rpoD; 2945 nucleotides total) generated using MUSCLE [33]. The phylogenetic tree was generated using PhyML [34] and the distance matrices were calculated by the Jukes-Cantor method. Bootstrap values over 50% (out of 1000 replicates) are indicated at the nodes. The presence of a black square indicates that that strain harbours the phl biosynthetic gene cluster. The three subgroups encompassing DAPG-producing strains are highlighted in color. Letters following the strain names correspond to multilocus phylogenetic groups described by Frapolli and colleagues [35].
Almario and colleagues [18] proposed that the phl cluster was acquired independently in these two groups and that this cluster was subsequently lost in some lineages of the P. corrugata subgroup. A recent study reported that the phl BCG is present in about half of the genomes sequenced from the P. corrugata subgroup [32]. Interestingly, this subgroup includes numerous phytopathogenic strains, prominently strains belonging to the species P. corrugata and P. mediterranea [36,37]. These phytopathogenic strains do not harbour the phl BCG, suggesting that this cluster could have been lost during the transition between commensal and pathogenic lifestyles. The fact that DAPG can trigger induced systemic resistance in some plant species [38,39,40] is an undesirable trait for a plant pathogen, which means that it could have been counter-selected in these lineages. Outside of these two subgroups, the phl BCG is also present in several other strains, both inside and outside of the P. fluorescens species complex [18,41]. The phl BCG has also been found outside of the Pseudomonas genus: the presence of the phl BCG has been reported in three non-pathogenic Betaproteobacteria, namely Pseudogulbenkiania ferrooxidans EGD-HP2, Chromobacterium vaccinii MWU328 and Chromobacterium piscinae ND17 [18].
The origin of this cluster remains unclear. Most authors agreed upon the fact that the acquisition of the phl BCG in the P. fluorescens species complex is an ancestral event [16,18,42]. The phl BCG might have been acquired separately by the different groups of DAPG-producing Pseudomonas spp. [18]. In Pseudomonas sp. OT69, the phl BCG is embedded in a putative genomic island, suggesting a more recent acquisition by this strain [18]. Kidarsa and colleagues [43] proposed that the phlACB genes might have been acquired from an Archaea. Homologs of these three genes are present in a contiguous gene cluster and in the same order in multiple groups of Archaea, where they may play a role in fatty acid metabolism [43].
This entry is adapted from the peer-reviewed paper 10.3390/metabo11030182
This entry is offline, you can click here to edit this entry!