Bile acids (BAs) are amphipathic sterols primarily synthesized from cholesterol in the liver and released in the intestinal lumen upon food intake. BAs play important roles in micellination of dietary lipids, stimulating bile flow, promoting biliary phospholipid secretion, and regulating cholesterol synthesis and elimination. Emerging evidence, however, suggests that, aside from their conventional biological function, BAs are also important signaling molecules and therapeutic tools. In the last decade, the therapeutic applications of BAs in the treatment of ocular diseases have gained great interest.
- bile acids
- UDCA
- TUDCA
- ocular diseases
1. Introduction
Bile acids (BAs) are hormones which serve many different roles in the body’s digestive and metabolic systems. Their unique chemical composition contributes to their reported anti-inflammatory and antioxidant effects, explaining their historical use in traditional Chinese medicine (TCM) [1,2,3]. A total of forty-four different animal bile products isolated from both vertebrates and invertebrates have been used to treat ailments of the liver, skin, and biliary system. The earliest records suggest that bile salts from dog and ox were first used as therapeutics; however, the use of bear bile continues to be the most popular and of particular interest in today’s exploration of BA-based therapy [1,2]. Bile and gallbladders collected from black (Selenarctos thibetamus) and brown (Ursus arctos) bears have been used for thousands of years to remove toxins from the body, stop convulsions, and improve vision [1,3]. In TCM, as a medicine “cool” in nature, bear bile is used to treat “hot” diseases like fever, inflammation, swelling, and pain [1,3]. Modern analysis of bear bile shows that it contains relatively high levels of ursodeoxycholic acid (UDCA) and tauroursodeoxycholic acid (TUDCA) and has numerous pharmacological properties, such as: antimicrobial, anti-inflammatory, anti-convulsant, anti-hepatotoxic, and sedative [3,4,5,6]. The hepatoprotective, anti-gallstones and hypolipidemic effects of bear bile have led to its use as a therapeutic for liver disease and biliary cirrhosis in both TCM and modern Western medicine [2,3,7].
Current interest in BAs arose from the discovery of their signaling properties and ability to regulate the activity of many genes through the orphan nuclear receptor farnesoid X receptor (FXR) [8]. FXR agonists have shown potential as treatments for primary biliary cirrhosis and nonalcoholic steatohepatitis [8,9,10,11]. The Discovery of another BA receptor, transmembrane G-protein-coupled receptor 5 (TGR5), and its widespread distribution in the body, has brought on new explorations on BA-based therapies [8] and their extra-hepatic homeostatic functions on lipid, glucose, and energy metabolism [12,13,14,15]. Interestingly, more recent studies have shown that BAs are protective in neurodegenerative diseases, including a number of ocular afflictions.
2. Bile Acid Physiology and Metabolism
BAs eliminate cholesterol, aid in lipid transport, and stimulate bile flow and biliary phospholipid secretion [16]. BAs maintain liver cells viability while regulating the storage and production of hepatic glucose [17]. Additionally, BAs play a critical role in cholesterol catabolism, being responsible for nearly 50% of its daily turnover [18]. Primarily responsible for lipid digestion and absorption, BAs have been known to act in the small intestine as digestive detergents [18]. Conjugated BAs give rise to bile salts, of which amphiphilic nature allows them to form micelles and work in tandem with lipases at the intestine’s brush border to digest fats [19].
BAs are produced in hepatocytes and are transported to the gallbladder through bile canaliculi. There are two major pathways for BA synthesis: the classical and alternative pathways [16,20] (Figure 1). The classical pathway is initiated by cholesterol 7α-hydroxylase (CYP7A1) and regulated by FXR [14,21,22]. CYP7A1 is the only rate-limited enzyme in BA synthesis and leads to the production of the two primary BAs: cholic acid (CA) and chenodeoxycholic acid (CDCA). In the classical pathway, CYP7A1 is converted to 7 α- hydroxy-4-cholesten-3-one (C4), which is the precursor to both CA and CDCA [14,16,22]. C4 can then be converted to CA with 12α-hydroxylase (CYP8B1), and without this conversion, C4 is converted to CDCA [14,16,22]. Mitochondrial sterol 27-hydroxylase (CYP27A1) catalyzes the steroid side-chain oxidation and subsequent proximal β-oxidation of C4 to form 24-carbon compounds such as CA and CDCA [14,16,22].
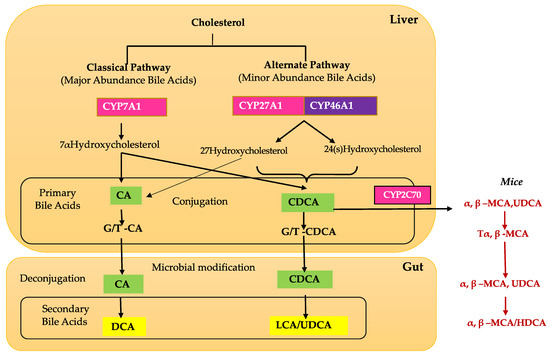
The alternate pathway is initiated by sterol 27-hydroxylase (CYP27A1), a mitochondrial cytochrome P450 enzyme that is widely distributed in macrophages and most tissues [14,23,24]. Studies have shown that the alternative pathway contributes to around 9% of total BA synthesis in human hepatocytes [16]. Initiation by CYP27A1, followed by CYP7B1 (25-hydroxycholesterol 7-alpha-hydroxylase), leads to the synthesis of CA and CDCA in the liver and extrahepatic tissues such as steroidogenic tissues and macrophages [16,24].
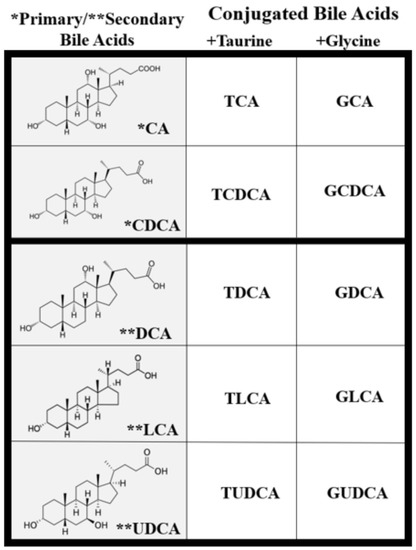
Once in the intestinal lumen, gut microbiota produces secondary bile salts by modifying primary BAs with 7α-hydroxylation, deconjugation, oxidation, and epimerization of hydroxyl groups on carbon-3, carbon-7, and carbon-12 [21,25,26]. Bile salts, conjugated BAs, are deconjugated via hydrolysis of the amide bond between glycine or taurine and the BA, allowing the BAs to be used as substrates for modification by the microbiota. This reaction is catalyzed by bile salt hydrolases (BSH) in bacterial enzymes [27,28,29] (Figure 2). Dehydratases from the anaerobic gut flora turn unconjugated CA and CDCA into deoxycholic acid (DCA) and lithocholic acid (LCA), respectively, and epimerization of hydroxyl groups of CDCA by hydroxysteroid dehydrogenases of intestinal bacteria leads to the formation of UDCA [30].
Unconjugated BAs are reabsorbed into intestinal enterocytes via passive diffusion in the jejunum and colon, whereas conjugated BAs, bile salts, are reabsorbed through a sodium-dependent BA transporter (ASBT) in the jejunum [27,31]. While 95% of BAs are reabsorbed and return to the liver via the portal circulation, the remaining 5% do not get reabsorbed at the intestinal lumen and are excreted in feces [14,32]. This mechanism puts a constant demand on the liver to continue BA synthesis. Though BAs are recycled through enterohepatic circulation, less than 10% of BAs reach systemic circulation [14], where they can be detected in both plasma and cerebrospinal fluid on the scale of ng/mL [33]. However, BA concentration has never been measured in ocular fluids [21].
3. Cytotoxic and Cytoprotective Effects of Bile Acids
Though BAs are an integral part of numerous biological processes, they can also exert toxic effects and cause inflammation, apoptosis, and cell death [16,34,35]. BA concentration and hydrophobicity are important in determining their toxic or protective effects, with a more hydrophobic BA being more toxic. Increased retention of hydrophobic BAs promotes hepatocyte apoptosis or necrosis that may lead to liver injury [34,36,37]. BA hydrophobicity is determined by the number, position, and orientation of its hydroxyl groups along with amination at its C-24 position and, based on this, the BA sequence from least to most hydrophobic is: UDCA, CA, CDCA, DCA, and LCA [34,38] (Figure 2). Once a BA is conjugated, however, it becomes more hydrophilic and, therefore, less cytotoxic [34,38]. Defective transport, faulty biosynthesis, or excess bacterial degradation may lead to an excess of toxic BAs. Specific conditions associated with high intracellular and extracellular concentrations of BAs include cholestasis and BA malabsorption, respectively [27]. Disorders associated with dysregulated BA metabolism include cholestatic liver disease, dyslipidemia, fatty liver diseases, cardiovascular diseases, and diabetes, among others [16,39,40].
Studies have shown that the accumulation of hydrophobic BAs inside hepatocytes leads to their injury and death. In one study, hepatocytes expressed dramatic levels of subcellular damage when exposed to concentrated CA and CDCA [34]. The canalicular membranes were particularly altered in the presence of concentrated LCA and tauro-LCA [36], the most hydrophobic of the common BAs. Moreover, biliary epithelium cells were very sensitive to the cytotoxicity of hydrophobic unconjugated bile salts while they remained largely undamaged by taurine- or glycine-conjugated bile salts [36].
Aside from their hydrophobicity, abnormal amounts of BAs can lead to toxic effects on hepatic cells as well. Decreased BA concentration will stimulate hepatic cell apoptosis, while increased BA concentration will stimulate necrosis [34,41]. Cytotoxic BAs were shown to alter membrane cholesterol levels, whereas cytoprotective BAs had no effect on the membrane. This evidence suggests that BA cytotoxicity could depend on their effects on plasma membrane integrity or on mitochondrial membrane affinity for apoptotic proteins [42]. BAs have also been shown to induce injury on several cell types, including hepatic, biliary, gastrointestinal, renal, and lung cells [34]. Interestingly, patients experiencing the cytotoxic effects of hydrophobic BAs might undergo hydrophilic BA-based therapy, which entails displacing cytotoxic BAs with the non-toxic UDCA or TUDCA.
UDCA and TUDCA, although present at very low circulating levels in humans, have been studied for their cytoprotective effects due to their hydrophilic nature and low toxicity [43]. Their mechanism of action is not fully understood; however, stabilization of cell membranes (structure and fluidity), protection against oxidative injury, and prevention of apoptosis have all been reported [34,43].
BA pharmacological use has been applied to pathologies such as cholelithiasis, primary biliary cirrhosis, primary sclerosing cholangitis, and cystic fibrosis and is being suggested for neurodegenerative disorders associated with increased apoptosis such as Huntington’s Disease [34,44]. The most significant effects of UDCA and TUDCA as therapeutic agents have specifically been studied in patients with either liver or digestive disease [2]. For example, UDCA is used in patients with cholestatic liver disease to slow disease progression by decreasing hepatocyte injury. Moreover, it can be used in patients with BA malabsorption to sequester BAs and provide symptomatic relief from diarrhea [45].
Some studies demonstrate that the mechanism by which UDCA protects membranes is dependent on membrane cholesterol. In one study, cholesterol was found to influence the toxicity of both UDCA and the highly toxic sodium deoxycholate (SDC). UDCA was able to mitigate the cytotoxic effects of SDC only in the presence of cholesterol, whereas, enhancing its cytotoxic effects in the absence of cholesterol. This relationship of events suggests that the extent of UDCA cytoprotective effects is dependent on the level of cholesterol in membranes. In the context of treating cholestasis patients with UDCA, one must consider changes in membrane cholesterol composition that might affect the efficacy of this drug [37].
4. Retinal Bile Acid Metabolism
Although the liver is the primary site of BA production, several recent studies have identified extrahepatic production sites, including the brain and retina [14,21]. The BA pathway plays an important role in cholesterol metabolism in the retina. Emerging evidence and our unpublished data show that unlike the liver, the brain and retina mainly rely on the alternate BA biosynthetic pathway [14]. The first step in the alternate pathway is initiated by the conversion of cholesterol into 27-hydroxycholesterol via CYP27A1. Subsequently, oxysterol 7α-hydroxylase (CYP7B1), through subsequent hydroxylation, leads to the formation of primary BAs, CA, and CDCA (Figure 1). Simultaneously, the formation of 24(S)-hydroxycholesterol from cholesterol is carried out by CYP46A1 (cholesterol 24-hydroxylase). 24(S)-hydroxycholesterol is then metabolized via oxysterol 7α-hydroxylase II (CYP39A1) to generate CDCA (Figure 1). Cholesterol involvement is implicated in several retinal pathologies, such as in age-related macular degeneration, where large cholesterol-rich deposits are a hallmark feature [46]. CYP27A1 and CYP46A1 are two enzymes that are important to ocular function in humans. In the retina, CYP27A1 is the principal hydroxylase that catalyzes the hydroxylation of cholesterol for BA synthesis to account for the majority of enzymatic cholesterol elimination in the retina [46]. Deficiency in CYP27A1 is seen in cerebrotendinous xanthomatosis, a rare congenital disease characterized by several neurological and ocular symptoms such as cataracts, neurological dysfunction, and premature retinal degeneration [46]. One study found an increase in focal cholesterol deposits along Bruch’s membrane in CYP27A-/- mice [46]. Combined deficiency in CYP27A1 and CYP46A1 results in neovascularization and lipid accumulation in the mouse retina [47]. CYP46A1 is expressed in both the brain and the retina, though mainly the neural retina, and has a role in maintaining retinal cholesterol homeostasis and retinal immune response [48]. A single nucleotide polymorphism in the CYP46A1 gene, rs754203, is a risk factor for glaucoma and has also been identified as a predictive marker for the risk of cataracts [49]. Based on the current literature, it is evident that the retinal BA pathway is understudied, and more studies are required to fully appreciate and understand BA retinal metabolism and functions.
5. Bile Acid Transporter and Receptor in the Retina
BAs require essential transporters to maintain their enterohepatic circulation and facilitate their active transport through membrane barriers in the liver and intestine. The sodium BA cotransporter family (SLC10) is a solute carrier family of genes that has been found to contain several BA transporters localized in many different tissues in the body. Most studied are the Na+/taurocholate co-transporting polypeptide (NTCP; SLC10A1) and the apical sodium-dependent BA transporter (ASBT; SCL10A2), which serve their primary transport functions in the liver and intestine, respectively. The gene SLC10A4 has been localized in the human eye, and it is associated with cholinergic synapses throughout the central nervous system. While SLC10A4 is also responsible for encoding protein P4, little data supporting the functional properties of the P4 gene is currently available [50,51].
Emerging evidence suggests that aside from their conventional biological function in the enterohepatic system, BAs are also important signaling molecules [16] by activating specific receptors, such as FXR and TGR5, and their downstream signaling cascades. In the retina, the FXR and TGR5 receptors are currently the most studied, however, reports on FXR retinal expression remain inconsistent, and TGR5 expression has been reported only in the adult retina [21,52,53,54]. Other BA receptors have been identified, such as vitamin D receptor (VDR), pregnane X receptor (PXR), glucocorticoid receptor (GR) and mineralocorticoid receptor (MR), constitutive androstane receptor (CAR), α5 β1 integrin, and sphingosine-1-phosphate receptor 2 (s1PR2) [55,56]. The biological functions of these receptors are much less characterized and have not been previously studied in the retina. Furthermore, the literature describing the importance of BA receptors in a wide range of diseases of the liver, biliary system, and gastrointestinal tract is vast [57], whether their extrahepatic functions are underinvestigated. No current information is available on BA receptors expression pattern in the developing retina.
6. Pharmacological Effects of Bile Acids in Ocular Disease
In the last decade, studies on the usefulness of BAs in the treatment of ocular diseases have gained great interest in the scientific community. Particularly, UDCA and TUDCA are reported to be beneficial against photoreceptor degeneration, ganglion cell death, cataract, diabetic retinopathy (DR), etc. [21]. Previously, the neuroprotective properties of BAs in retinal diseases have been reviewed by multiple authors [58,59,60]. Herein we summarize recent advances in using BAs to treat ocular diseases and highlight significant lacunae and future studies. Additionally, our current review also discusses the recent studies, including ours, on the effects of BAs and their receptor agonists in the retinal vasculature in various forms of retinal diseases, including DR and retinopathy of prematurity (ROP). Of interest, our review also focuses on the role of the BA-gut microbiome axis in various retinal diseases, a topic that was not discussed in previously published review articles.
Although the mechanism of action of UDCA and TUDCA remains unclear, it is thought to involve suppression of both caspase-dependent and caspase-independent apoptosis, likely by inhibiting the release of the apoptosis-inducing factor (AIF) from the retinal pigment epithelium (RPE), photoreceptors, and retinal ganglion cells (RGC) [21,61,62].
Retinal detachment, oxidative stress, and inflammation are also a key contributor to photoreceptor degeneration following their detachment from the RPE [63]. Photoreceptor cell death and retinal detachment are associated with numerous ocular diseases such as ROP, retinitis pigmentosa, age-related macular degeneration, and DR [63,64,65,66,67]. A study by Mantopoulos et al. showed that TUDCA preserved photoreceptor survival after retinal detachment by inhibiting caspase activity and reducing ER and oxidative stress [68].
In galactosemic rats, TUDCA treatment alleviated cataract formation by inhibiting the unfolded protein response (UPR)-dependent pathway [69]. Cataract formation results from stressors leading to ER and oxidative stresses of lens epithelial cells (LECs), resulting in a UPR-dependent death pathway. TUDCA rescued cultured human LECs that were treated with ER stressors. The same group also reported that galactosemic rats treated with TUDCA had significantly reduced LEC death and partially delayed hyper mature cataract formation [69]. However, in selenite-induced nuclear cataract in rats, a control demonstrating the UPR-independent death pathway, TUDCA did not prevent LEC death and did not alleviate cataract formation. Overall, TUDCA showed potential as a prophylactic drug for some types of cataracts in which ER stress and LEC death play a central role [69]. When the ER is subjected to multiple pressures, unfolded proteins accumulate within the organelle, stimulating the UPR. The consequences of this response are ultimately excessive ER stress and cell death [70,71].
One study has explored the significance of TUDCA structural resemblance to compounds that bind to a light-activated form of rhodopsin to inhibit the activation of transducin. This study used computer modeling systems to show that TUDCA docks to light-activated rhodopsin using a similar mode of binding to the C-terminal domain of the transducin alpha subunit. These results suggest that TUDCA specific interaction with rhodopsin may contribute to its effect as a potential therapeutic compound for degenerative retinal diseases [53].
Retinal degenerative diseases are important causes of blindness with little or no present effective cures. BA therapeutic potential has been tested in several models of retinal degeneration. Leber congenital amaurosis (LCA) is a form of retinal dystrophy that often leads to severe vision impairment and blindness; it has been shown that ER stress induced by S-opsin aggregation leads to rapid cone degeneration [72,73,74]. TUDCA has been shown to reduce ER stress and apoptosis in mice models of LCA and significantly preserve photoreceptors survival [62,73].
In one model of light-induced retinal damage, TUDCA treatment was shown to reduce superoxide radical levels [75]. TUDCA treated mice were found to have significantly higher a- and b-wave amplitudes on ERG readings and presented with milder retinal functional damage seven days post-light exposure when compared to mice given other treatments [75]. Similar findings were also shown in a model of light-induced retinal degeneration (LIRD). Mice treated with TUDCA showed greater ERG a-wave and b-wave amplitudes compared to control mice. Moreover, TUDCA-treated mice were found to have thicker outer nuclear layers, more photoreceptor cells, and more fully developed photoreceptor outer segments [76]. Collectively, these results indicate that TUDCA slows retinal degeneration.
In models of retinitis pigmentosa (RP), TUDCA has also been studied as a potential therapeutic agent. The common outcome of the RP disease family includes apoptosis of rods followed by degeneration of cones [77,78,79]. One of the main causes of RP is a mutation in the GTPase regulator (RPGR) gene, which codes for a scaffold protein responsible for ciliary protein trafficking processes [80]. In one study, TUDCA was found to suppress microglial activation preceding retinal degeneration in an RPGR conditional knockout mouse model [80]. Additionally, TUDCA treatment inhibited inflammation and prevented photoreceptor degeneration. Together, these findings support TUDCA as a potential therapeutic tool for RPGR-associated RP.
Boatright et al. utilized rd10 mice to test the efficacy of TUDCA in RP [76]. Rd10 mice have a missense mutation in phosphodiesterase 6B, leading to rod degeneration around postnatal days 14-16 (P14-P16). Their results showed that systemic injection of TUDCA prevented apoptosis during the early stage of the disease and preserved function and morphology of photoreceptor cells in these mice by inhibiting caspase-mediated photoreceptor apoptosis. [76]. In another study by the same group, TUDCA ability to preserve photoreceptors during retinal degeneration in rd10 mice was further analyzed and confirmed at later stages (P30) when photoreceptor cell loss peaks [81]. At P30 in the rd10 model, the majority of rods have degenerated, and only some cones remain [82] resembling the end-stage RP for most patients [83]. TUDCA treatment between P6 and P30 preserved overall photoreceptors number and function [81] and significantly reduced caspase 3 activation and apoptosis. Systemic TUDCA treatment in these mice was able to delay retinal damage to at least P30. In this case, TUDCA treatments were also able to slow the loss of both photoreceptors and retinal function. Additionally, TUDCA treatment was found to slow the loss of RGC and prevent epithelial cell death [84].
In another study of retinal degeneration, photoreceptor degeneration in mice retinas was chemically (N-methyl-N-nitrosourea) induced [85]. To explore TUDCA as a potential therapeutic agent, TUDCA was subcutaneously injected. Retinal flat-mounts of TUDCA treated mice showed efficient cone photoreceptor preservation. Additionally, TUDCA therapy rectified abnormalities in the visual signal shown to inhibit photoreceptor degeneration and decrease visual impairments in mice that received transmission. A multi-electrode array was used to detect the restrictive effect of TUDCA therapy on spontaneous firing response as well as the supportive role TUDCA therapy plays in enhancing the light-induced firing response. In degenerative retinas, TUDCA treatment was noted to preserve the basic configurations of the ON–OFF signal pathway. The results of this study suggest subcutaneous delivery of TUDCA has therapeutic potential in retinopathies with progressive photoreceptor degeneration [85].
BAs were also tested in models of optic nerve injury. Optic nerve transection was performed on C57BL/6 mice following systemic TUDCA treatment for 10 days. Immunohistochemistry staining on retinal flat mounts indicated a loss of 60% of RGC in control mice, which was completely prevented in mice treated with TUDCA [84].
Diabetic retinopathy, a retinal neurovascular disorder, is the leading cause of blindness in adults of working age groups in the Western world [86,87]. UDCA has also been found to reverse the breakdown of the blood-retinal barrier (BRB) and reduce retinal inflammation in models of DR [88]. Reduced retinal expression of interleukin-1β and interleukin-6 in response to UDCA treatment suggests that UDCA inhibits retinal inflammation and attenuates BRB breakdown. Moreover, UDCA treatment increased expression of the tight junction proteins claudin-1 and claudin-19 and led to a reversal of the reduced thickness of both the inner and outer nuclear layer of the retina [88]. UDCA’s suggested ability to reverse damage to retinal layers typically compromised in ocular disease continues to drive research toward its use as a therapeutic agent for DR.
Pericytes play an important role in maintaining vascular integrity, and it is thought that retinal pericytes loss, an early hallmark of DR, leads to the BRB breakdown [89,90,91,92]. Endoplasmic reticulum stress may be involved in this process [93]. UDCA has also been reported to reduce pericyte depletion in models of DR [93]. UDCA clearly attenuates the increase in ER stress and prevents loss of pericytes and vascular integrity in DR [93]. Another recent study investigated the effects of UDCA in pericyte depletion mice by injection of an antibody against platelet-derived growth factor reception-β (PDGFR-β clone APB5) [94]. UDCA reduced the expression of F4/80+ macrophages in the APB5-induced retina according to immunofluorescent labeling. UDCA also reduced the increased expression of angiogenic factors and inflammatory mediators (vascular endothelial growth factor; VEGF, intercellular adhesion molecule-1, and monocyte chemotactic protein-1). Along with microvascular changes, neural components of the retina are also affected in DR [95,96]. A study by Gaspar et al., 2013, evaluated the neuroprotective effects of TUDCA in high glucose-treated rat retinal neurons. TUDCA was able to prevent cell death induced by elevated glucose through its anti-apoptotic and antioxidant properties in cultured retinal neural cells. Together, these results suggest that UDCA and TUDCA attenuate retinal vascular and neural abnormalities associated with DR [93,94].
TGR5 receptor plays an important role in regulating metabolic homeostasis [15]. TGR5 activation has also been linked to improving retinal function, and TGR5 agonists have been reported to improve biliary epithelial barrier function [97]. One study demonstrated that treatment of Streptozotocin (STZ)-induced diabetic rats with TGR5 agonist INT-777 was protective against hyperglycemia-induced vascular permeability [97]. Another study by Beli et al., 2018 showed that TGR5 activation using INT-767 significantly reduced acellular capillaries and inflammation in the db/db mice. Particularly, INT-767 treatment to diabetic mice significantly reduced the number of acellular capillaries and numbers of macrophages, leukocytes, and activated microglia in the db/db mice retina [52]. Together, these results emphasize that TGR5 activation could be a viable therapeutic strategy to ameliorate DR.
Age-related macular degeneration is known to result in severe visual loss due to the development of choroidal neovascularization (CNV) along with related manifestations, such as subretinal hemorrhage, RPE detachment, and fibrovascular disciform scarring, also known as wet or exudative AMD [98,99,100]. Woo, et al. demonstrated that UDCA and its derivative, TUDCA, when intraperitoneally injected to the laser-treated rat model, could effectively suppress early VEGF elevation in the retina which inhibits the upregulation of VEGF and eventually results in the reduction in CNV size and vascularity, suggesting the possibility of treating CNV [101]. In a recent study, a mouse model of laser-induced CNV was used to test the efficacy of YSB201 (an aqueous solution of UDCA) in the treatment of AMD [102]. Results indicated that the YSB201 treatment inhibited the return of angiogenesis to the normal choroidal tissue condition. Additionally, while YSB201 treatment at 125 mg/kg/day led to the recovery of retinal function post-injury, the higher dose of YSB201 treatment at 250 mg/kg/day did not yield significantly higher recovery effects. The results of the study specifically describe the inhibitory effects of UDCA formulation on CNV with functional recovery in mice retinas [102]. Taken together, these results suggest that UDCA could be used as a potent supplement for the cure of AMD and related retinal complications.
Further evidence of UDCA therapeutic potential was seen in our recent study of ROP, a leading cause of preventable blindness in children [103,104]. We investigated the efficacy of the secondary bile acid, UDCA and its taurine and glycine conjugated derivatives TUDCA and glycoursodeoxycholic acid (GUDCA) in preventing retinal neovascularization (RNV) in an experimental model of ROP. While all three secondary BAs have pharmacological effects, this ROP model found that UDCA was most effective in demonstrating anti-angiogenic effects, preventing loss of neuronal cells, and protecting against reactive gliosis and BRB breakdown while maintaining normal revascularization of the central retina [105]. UDCA was also able to decrease VEGF and inflammatory cytokine expression [105]. Collectively, our results suggest that UDCA could represent a new effective therapy for ROP.
Together, these findings support the idea that BAs have the potential to show their effects at nearly every structural layer of the eye. Evidence continues to support their therapeutic potential in neurodegenerative and ocular disorders.
This entry is adapted from the peer-reviewed paper 10.3390/biom11020292