The main reason for the development of the yield gap in crop production is the inefficient man-agement of nitrogen (N). The nitrogen gap (NG) cannot be ameliorated without an indication and quantification of soil characteristics that limit N uptake by a crop plant. The insufficient supply of N to a plant during its cardinal stages of yield formation is a result of two ma-jor-variabilities. The first is spatial variability in the soil characteristics responsible for water supply to a plant, also serving as a nutrient carrier. The second is a vertical variability in soil factors, decisive for pools of available nutrients, and their in-season accessibility to the grown crop. The long-term strategy for NG cover should focus first on soil characteristics (humus stock, pH, nutrient content) responsible for water storage and its availability to the currently grown plant.
- nitrogen use efficiency
- soil constraints
- subsoil
- spatial variability
1. Food Gap and Sustainable Intensification of Agriculture
The global human population, depending on the scenario, will reach 9–10 billion in 2050. Some prognoses of food requirements in 2050, based on the level of food production in 2005, fluctuate from 50% to 110% [1][2]. The analysis made by Hunter et al. [3] showed a much lower level of food demand in 2050 compared to 2010, ranging from 25% to 70%. The effective management of global food demand in 2050 should be a result of the simultaneous implementation of three complementary strategies, termed ”mega-wedges” [4]. They are as follows: (i) filling the food production gap, (ii) decreasing food losses in the entire food chain, (iii) reduction in the worldwide food demand. The relative contribution of each particular mega-wedge in food security control has been assessed as 46%, 33.6%, and 20.4%, respectively. The second strategy does not include only direct losses of energy and proteins in the food chain, but also reduced productivity of soil and water (10.4%). This means that above 55% of the future food demand directly depends on efforts oriented on covering the food production gap. With respect to food losses, it is necessary to stress that the net excess of the global protein supply is almost equal to its intake by humans (36% vs. 44%) [5]. It can be, therefore, concluded that the management of the food production chain requires significant changes in agriculture. The conceptual (diagnostic, management, techniques, technology) preparation of the agriculture sector for a considerable increase in food production in the coming 30 years is the key challenge for the present generation.
The current level of food production is a function of two main factors, i.e., (i) actual crop yield, (ii) crop yield improvement. The actual yield of a particular crop in a strictly defined locality (field) is a result of the efficiency of production inputs under the course of meteorological conditions during the growing season [6][7]. The realization of the second target depends on progress in (i) breeding of new cultivars, (ii) improvement in water efficiency (WUE), (iii) improvement in nitrogen use efficiency (NUE). It is necessary to remember that the success of the Green Revolution, resulting in the significant increase of yields of major crops (cereals, rice, maize) was a result of the induced synergism between new, high-yielding cultivars, high rates of applied fertilizer nitrogen (Nf), and a high level of crop protection, based on agrochemicals [8]. An insufficient level of the required synergism between these three main factors has resulted in stagnation in the world average yields of main crops during the last two decades [9]. The necessary annual rate of yield increase of four global crops. i.e., maize, rice, wheat, and soybean to cover the food gap in 2050 would have to reach 2.4%. The current yearly rate of yield increase for these four crops is far below the assumed threshold, being at the level of 1.6%, 1.0%, 0.9%, and 1.3%, respectively [10]. For example, the genetic progress in nitrogen use efficiency for wheat in the years 1985–2010 was only in the range of 0.30–0.37% y−1. This was a result of the increased value of the Nitrogen Harvest Index (NHI), i.e., the relative amount of N accumulated in grain [11].
Future progress in food production, in fact, will depend on two key drivers. The first is high-tech intensification, based on highly productive cultivars of main crop plants, and the efficient use of mineral fertilizers and other agro-chemicals. This strategy has been responsible for about ¾ of the global food production increase in the last 85 years. The remaining ¼ was due to the increase in the area of arable land, mainly in less-developed countries. The productivity of the newly cultivated soils is to a great extent driven by their natural fertility. In the future, this option will be strongly limited due to the lack of high or even medium fertile soils. The primary resources of potential arable land are extensive pastures and tropical forests. The key disadvantage of the first is a shortage of water and of the second, low fertility, mainly due to high acidity and a shortage of essential nutrients. In addition, this option requires a considerable financial input to increase soil fertility, as a prerequisite of an economically and environmentally sound level of production [12][13].
The progressively increasing demand for food, resulting from the permanently growing human population, is in contradiction to the concept of sustainable development of agricultural production. The most advanced scenarios of a sustainable approach to food production assume a massive reduction or even the elimination of N fertilizers and pesticides. The main objective of this restrictive view of agriculture is to arrest the degradation of both local and global environments [14]. In the current and future reality, food demand is too high to completely abandon modern production measures [15][16]. A realistic view of sustainable agricultural development, termed as sustainable intensification of agriculture (SIA), defines this concept as a process or production system where yields are increased without an adverse impact on the environment and without the cultivation of low-quality land [17]. A less sophisticated, but at the same time a more practical definition of SIA, has been proposed by Smith [18]. The core of this definition is to produce more, and high-quality food per unit of used and applied production measures, taking into account both soil fertility and externally applied measures (fertilizers, pesticides, fuel) on the one hand, and protection of the existing ecosystems against damage on the other hand. These opinions are summarized in the concept of twin objectives, which relies on the assumption that agricultural development, including new tools, both implemented as new production technologies, production systems, and management, has to ensure the stability of the global ecosystem [19][20].
The area of arable land and the amount of available water during the growing season of a particular crop plant are key factors that limit food production for a country, region, or field. A rise in food production cannot be based solely on the increased efficiency of water usage. As reported recently by Grafton et al. [21], based on an analysis of numerous methods of crop plant irrigation, no direct substitution between water and nitrogen was observed. The key factor limiting yield, as results from this study, is nitrogen. The authors clearly stated that the greatest challenge to the progress of crop plant productivity depends on the improvement in N management. The required rate of food production increase to cover the food gap by 2050 can be achieved by an adequate supply of fertilizers, both nitrogenous, and those balancing N, i.e., containing P, K, Mg, S, and micronutrients. Scenarios of nitrogen fertilizers (Nf) consumption in agriculture to cover the food production gap vary 3-fold, i.e., from 85 Mt N y−1 to 260 Mt N y−1 [22]. In the model of N flows by Conijn et al. [23], the total consumption of Nf will increase to 181 Mt y−1, i.e., by 76% in 2050 as compared to 2010 (103 Mt N y−1). The increased Nf consumption will lead, however, to a simultaneous increase in losses of its active compounds into the environment. The N loss in 2050 is projected to lie in the range of 102–156% with respect to 2010 [24].
Any increase in the production efficiency of both key agronomic factors, i.e., water and nitrogen, depends on the soil status of all the other production factors decisive for their efficiency. The production of phosphorus fertilizers to cover crop plant requirements for this nutrient is set to increase in the period 2010–2050 by 32% (from 17.9 to 23.7 Mt P y−1) [23]. An analysis made by Pradhan et al. [25] showed that to fulfill food production goals in 2050 the consumption of N, P, and K fertilizers will increase in the range of 45–73% for N, 22–46% for P2O5, and 2–3-fold for K2O, respectively, compared to 2010. The sustainable intensification of agriculture cannot be realized without taking into account other nutritional factors that limit the productivity of water and nitrogen, such as magnesium, sulfur, and micronutrients [26][27].
2. The In-Season Management of Nitrogen—Cardinal Growth Stages
The first task in the reorientation of the crop production system is to calculate the total requirement of a currently grown plant for nitrogen. This can be calculated for an average yield harvested on a particular field or based on the potential yield of a given cultivar in the same climatic region. Two additional components have to be taken into account to make a reliable estimation of the total N requirement by the currently grown cultivar. The first is nitrogen concentration in the main product, for example, seeds or grain. There is still ongoing scientific discussion with respect to the extent to which N concentration in seeds or grain is a conservative, i.e., genetically, or environmentally governed trait [28][29][30]. The second component refers to the partitioning of N taken up by the crop between the main yield, and its by-product, for example, between grain and straw. At harvest, this process refers to the value of the nitrogen harvest index (NHI), which is a conservative trait of seed crops [31].
Total N input (Ni) in the soil/crop system for an assumed yield of grain/seeds is calculated based on the algorithm:
|
Ni = (YCPY ∙ GNc)/NHI |
(1) |
where:
Ni—nitrogen input, kg ha−1;
YCPY—climatic potential yield of the grown cultivar, kg ha−1;
GNc—grain/seed nitrogen concentration, kg t−1;
NHI—nitrogen harvest index, a value in the range of 0.6–0.8.
The key objective of nitrogen fertilizer (Nf) application is to synchronize its application time with the crop plant requirement. The dominant factor is the stage of plant growth and the required content of N, which progressively increases with the crop growth. The right determination of the Nf dose in the critical stage of yield formation is, therefore, the decisive factor in the exploitation of the yielding potential of the currently cultivated crop. The crop demand for N in a particular growth stage depends on the rate of plant biomass growth, primarily driven by temperature and water supply [32]. The sum of physiologically active temperatures (GDD), and water and nitrogen supply are major factors for the quantity of biomass produced by the crop during the respective phase [33]. The rate of seed crop growth throughout the vegetative season is not constant. Based on this criterion, three mega-phases can be distinguished, named as canopy foundation (CF), yield component construction (YCC), and yield realization (YR) [34]. The first two periods describe the vegetative part of the plant life cycle and the third one its reproductive phases (Figure 1). The shape of the dry matter accumulation curve can be described mathematically using very sophisticated models [35]. In cereals, the CF period, extending from sowing up to the end of tillering, is responsible for the number of tillers per plant. The course of dry matter accumulation is best described by the exponential regression model. The YCC period, extending from the beginning of stem elongation up to flowering, is responsible for the set of flowers. The rate of dry matter accumulation during this period is best described by the linear regression model. The dry matter yield of wheat at the end of the CK2 can be used to make a prognosis of grain yield [36].
The borderline stages of plant growth, which occur between the mega-phases of CF and CYC, and between the CYC and YR periods, can be named Cardinal Knots (CKs), i.e., CK1 and CK2, for the first and second borderline of consecutive mega-periods, respectively. These two CKs are decisive stages for yield component formation. For example, as shown in Figure 2, the content of soil nitrate-nitrogen (NN) during the YCC period of oilseed rape undergoes a strong depletion (=N uptake by WOSR during the YCC period). The recorded NN depletion significantly affected the seed density, which was the key yield component, determining seed yield (R2 = 0.89, p ≤ 0.001). What is most important, however, is the fact, that the dependence obtained clearly defines the rosette stage (BBCH 30) as the decisive stage for the yield prognosis (NNop = 163.3 kg ha−1 for Yamax = 77.6 · 1000 seeds m−2). The result obtained indicates that in farming practice, the time of Nf application has to precede the BBCH 30. In maize, the second rate of Nf is applied, based on Nmin determination, at the 5th leaf stage [37][38].
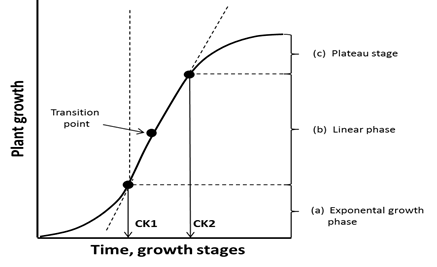
Figure 1. A conceptual pattern of dry matter accumulation by a typical seed/grain crop. Legend: CK1, CK2—cardinal stage 1 and 2, respectively (author’s own concept).
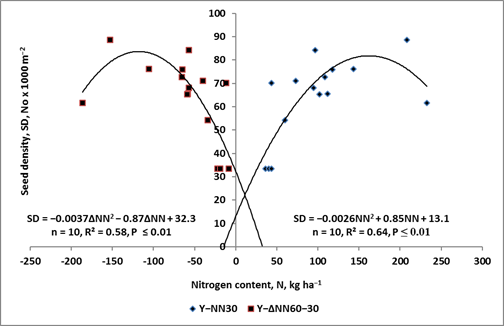
Figure 2. The response of seed density of winter oilseed rape to the content of nitrate nitrogen at the rosette stage and its uptake by the crop up to flowering. (based on [39]. Legend: NN- the content of nitrate nitrogen at BBCH 30; ΔNN—the change of the NN content during the period extending from the rosette up the flowering stage of WOSR growth.
The yield realization (YR) period commences from the onset of flowering and persists up to the physiological maturity of the plant. It can, however, be divided into two parts. The first part extends from flowering up to the watery stage of the seed/grain plant growth, i.e., to BBCH 71 (15% of seed/grain DW). It has well been documented that nitrogen supply to a seed crop, like cereals and oilseed rape, significantly affects the degree of yield component expression. The N pool accumulated by the seed plant during its vegetative growth, i.e., before flowering, considerably impacts the number of seed/grain per field unit area (physiological sink capacity) [40][41][42]. The second part of the YR period, which begins from the early milk-stage (BBCH 72) and finishes at physiological plant maturity (BBCH 89/90), is termed as the grain/seed filling period (GFP, SFP). The course of dry matter accumulation can be described by different mathematical models, but the linear or quadratic dominate. During this particular period, grain/seed reaches its final individual weight [43].
Efficient in-season N management, including knowledge of Nmin resources, should be oriented towards covering the plant N requirements during the period extending from the onset of flowering to the physiological plant maturity. For the seed crops, 75–85% of the N finally accumulated in seeds/grains originates from the pre-flowering resources, i.e., present in vegetative plant parts [44]. Post-flowering N management by the crop canopy can be described by three algorithms:
|
N remobilization quota: NRQ = Nhv Nfl, kg ha−1 |
(2) |
|
N remobilization efficiency: NRE = NRQ/Nfl · 100, % |
(3) |
|
Post-flowering N uptake: PNUP = Nt − Nfl, kg ha−1 |
(4) |
where:
Nhv—N amount in vegetative organs of crop canopy at harvest, kg ha−1;
Nfl—N amount in vegetative organs of crop canopy at the onset of flowering, kg ha−1;
Nt—total N amount in crop canopy at harvest, kg ha−1.
The patterns of N accumulation by the plant between particular CKs during the growth of a crop are the basis for a build-up of an efficient strategy of the in-season N status correction. Any free choice of Nf timing and its dose, as frequently observed in classic or even modern fertilization programs, does not fit the crop requirements for N, being the main reason for its inefficiency. A sound strategy of N management in seed crops, despite an almost similar accumulation pattern throughout the life cycle, is crop-specific. For example, in bread wheat, the key period of yield component formation extends from CK1 to CK2. During this period, the requirement for N results from both (i) the number of grain per unit area, and (ii) the required protein content in grain [45]. Consequently, a nitrogen fertilization strategy based on the correction of the plant N status in both CK1 to CK2 should also take into account protein concentration in grain. As a rule, any increase in the number of grains per unit field area results in a protein concentration decrease [46]. Therefore, any fertilization strategy, oriented to the increase in the crude protein concentration, requires an extra Nf dose, which should be applied at the end of the CK2. In maize, its nutritional status at the 5th leaf stage, which slightly precedes CK1, is decisive for the degree of yield component expression [47][48]. Nitrogen fertilizer should be applied just at such a time preceding this cardinal stage of maize growth because it affects the potential number of leaves and cobs. Nitrogen status in maize at the CK2, before the beginning of flowering, is important for the yield development, but it has only a predictive value [49]. In farming practice, it makes no sense to apply Nf at this particular stage because the yield was already fixed in much earlier stages of maize growth.
Nitrogen efficiency depends on the supply of other nutrients needed for its uptake and utilization [50][51][52]. It is necessary to pay attention to the fact that the accumulation of K by major crop plants, like wheat, oilseed rape, sugar beets, or potato, is as a rule higher as compared to N [53][54]. The maximum K uptake by high-yielding WOSR reaches its maximum during the phase of the main stem elongation. It can, therefore, be concluded that an efficient uptake of K from the subsoil by some crops, in comparison to N, is a necessary condition for the effective uptake of nitrogen. As recently reported by Grzebisz et al. , the efficient uptake of NO3- ions by WOSR depends on the adequate concentration of K and other nutrients, such as Ca and Mg in the sub-soil. All these nutrients are responsible for the development of yield components by crop plants.
One of the most important priorities in the breeding of crop plants is to increase the uptake efficiency of nutrients, especially of N and P from soil. Efficient acquisition of water and nutrients is required for the realization of both production and environmental goals. The efforts of key breeders have recently focused on the improvement of root system traits through [55][56][57][58][59]:
- Accelerating the early rate of root growth of short-season crops, for example, vegetables;
- Increasing root density in the topsoil to increase uptake of freshly applied fertilizers;
- Deeper rooting of crops grown in areas vulnerable to nitrate-nitrogen leaching.
- Stronger root branching to increase exploration of soil zones rich in mineral nitrogen;
- Increasing the density and length of root hairs for effective acquisition of phosphorus and potassium by a currently grown crop;
- Improvement in mycorrhiza association for effective acquisition of phosphorus and micronutrients.
This entry is adapted from the peer-reviewed paper 10.3390/agronomy11030419
References
- Tilman, D.; Balzer, Ch.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. PNAS 2011, 108, 50.
- Le Mouël, C.; Forslund, A. How can we feed the world in 2050? A review of the responses from global scenario studies. Eur. Rev. Agric. Econ. 2017, 44, 541–591.
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. BioScience 217, 67, 386–391.
- Keating, B.A.; Herrero, M.; Carberry, P.S., Gardner, J.; Cole, N.B. Food wedges: Framing the global food demand and supply challenge towards 2050. Glob. Food Sec. 2014, 3, 125–132.
- Berners-Lee, M.; Kennelly, C.; Watson, R.; Hewitt, C.N. Current global food production is sufficient to meet human nutri-tional needs in 2050 provided there is radical societal adaptation. Elem. Sci. Anth. 2018, 5, 52.
- Brevik, E.C. The potential impact of climate change on soil properties and processes and corresponding influence on food security. Agriculture 2013, 3, 398–417.
- Iizumi, T.; Ramankutty, N. How do weather and climate influence cropping area and intensity? Glob. Food Sec. 2015, 4, 46–50.
- Taiz, L. Agriculture, plant physiology, and human population growth: Past, present, and future. Theor. Exp. Plant Physiol. 2013, 25, 167–181.
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://faostat.fao.org/site/567/default.aspx#ancor (accessed 25 October 2020).
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428.
- Cormier, F.; Foulkes, J.; Hirel, B.; Gouche, D.; Moënne-Loccoz, Y.; Le Gouis, J. Breeding for increased nitrogen-use efficiency: A review for wheat. Plant Breed. 2016, 135, 255–278.
- Smith. P.; Gregory, P.J.; van Vuuren, D. Obersteiner, M.; Havlik, P.; Rounsevell, M.; Woods, J.; Stehfest, E.; Bellarby, J. Com-petition for land. Phil. Trans. R. Soc. B 2010, 365, 2941–2957.
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture (SOLAW)—Managing Systems at Risk; Food and Agriculture Organization of the United Nations and Earthscan: Rome, Italy, 2011.
- Conway, G. The Doubly Green Revolution: Food for All in the Twenty-First Century; Penguin Books: London, UK, 1997.
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. A review. Agron. Sustain. Dev. 2010, 30, 43–55.
- Pretty, J.; Bharucha, Z.P. Sustainable intensification in agricultural systems. Ann. Bot. 2014, 114, 1571–1596.
- The Royal Society. Reaping the Benefits: Science and the Sustainable Intensification of Global Agriculture; RS Policy document; The Royal Society: London, UK, 2009; p.86.
- Smith. P. Delivering food security without increasing pressure on land. Global Food Sec. 2013, 2, 18–23.
- Godfray, H.C.; Garnett, T. Food security and sustainable intensification. Phil. Trans. R. Soc. B 2014, 369, 20120273.
- Rockström, J.; Williams, J.; Daily, G.; Noble, A.; Matthews, N.; Gordon, L.; Wetterstrand, H.; DeClerck F.; Shah, M.; Steduto, P.; de Fraiture, Ch,; Hatibu, N.; Unver, O.; Bird J.; Sibanda, L.; Smith, J. Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio 2017, 46, 4–17.
- Grafton, R.Q., Williams, J.; Jiang, Q. Food and water gaps to 2050: Preliminary results from the global food and water systems (GFWS) platform. Food Sec. 2015, 7, 209–220.
- Mogollón, J.M.; Lasaletta, L.; Beusen, A.H.W.; van Grinsven, H.J.M.; Westhoek, H.; Bouwman, A.F.. Assessing future reactive nitrogen inputs into global croplands based on the shared socioeconomic pathways. Environ. Res. Lett. 2018, 13, 044008.
- Conijn, J.G.; Bindraban, P.S.; Schröder, J.J.; Jongschaap, R.E.E. Can our global food system meet food demand within planetary boundries? Agric. Ecosys. Environ. 2018, 251, 244–256.
- Bodirsky, B.L.; Popp, A.; Lotze-Campen, H.; Dietrich, J.Ph.; Rolinski, S,; Weindl, I.; Smitz, Ch.; Müller, Ch.; Bonsch, M.; Humpenöder, F.; Biewald, A.; Stevanovic, M. Reactive nitrogen requirements to feed the world in 2050 and potential to mitigate nitrogen pollution. Nat. Commun. 2014, 5, 3858.
- Pradhan, P.; Fischer, G.; van Velthuizen, H.; Reusser, D.E.; Kropp, J.P. Closing yield gaps: How sustainable can we be? PLoS ONE 2015, 10, e0129487.
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of K fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374.
- Wang, Z.; Hassan, M.U.; Nadeem, F.; Wu., L.; Zhang, F.; Li, X. Magnesium Fertilization improves crop yield in most production systems: A meta-analysis. Front. Plant Sci. 2019, 10, 1727.
- Gaju, O.; Allard, V.; Martre, P; Le Gouis, J.; Moreau, D.; Bogard, M.; Hubbart, S.; Foulkes, M.J. Nitrogen partitioning and remobilization to leaf senescence, grain yield and grain nitrogen concentration in wheat cultivars. Field Crops Res. 2014, 155, 213–223.
- Hawkesford, M.J. Reducing the reliance on nitrogen fertilization for wheat production. J. Cereal Sci. 2014, 59, 276–283.
- Maeoka, R.E.; Sadras, V.O.; Ciampitti, I.A; Diaz, D.R.; Fritz, A.K.; Lollato, R.P. Changes in the phenotype of winter wheat varieties released between 1920 and 2016 in response to in-furrow fertilizer: Biomass allocation, yield, and grain protein con-centration. Front. Plant Sci. 2019, 10, 1786.
- Hawkesford, M.J.; Riche, A.B. Impacts of G x E x M on nitrogen use efficiency in wheat and future prospects. Front. Plant Sci. 2020, 11, 1157.
- Malhi, S.S.; Johnston, A.M.; Schoenaou, J.J.; Wang, Z.H.; Vera, C.L. Seasonal biomass accumulation and nutrient uptake of wheat, barley, and oak on a Black Chernozem soil in Saskatchewan. Can. J. Plant Sci. 2006, 86, 1005–1014.
- Zhou, G.; Wang, Q. A new nonlinear method for calculating growing degree days. Sci. Rep. 2018, 8, 10149.
- Sylvester-Bradley, R.; Lunn, G.; Foulkes, J.; Shearman, V.; Spink, J.; Ingram, J. Management Strategies for Yield of Cereals and Oilseed Rape. HGCA Conference: Agronomic Intelligence: The Basis for Profitable Production. HGCA, 18. Available online: www.hgca.com/publications (accessed on 12 January2021).
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H. A flexible sigmoid function of determinate growth. Ann. Bot. 2003, 91, 361–371.
- Zerche, S.; Hecht, R. Nitrogen uptake of winter wheat during shoot elongation phase in relation to canopy high and shoot density. Agribiol. Res. 1999, 52, 231–250.
- Kahabka, J.E.; Van Es, H.M.; McClenahan, E.J.; Cox, W.J. Spatial analysis of maize response to nitrogen fertilizer in Central New York. Precision Agric. 2004, 5, 463–476.
- Luce, M.St.; Whalen, J.K.; Ziadi, N.; Zebarth, B.J. Nitrogen dynamics and indices to predict soil nitrogen supply in humid temperate soils. Adv. Agron. 2011, 112, 55–102.
- Grzebisz, W.; Łukowiak, R.; Kotnis, K. Evaluation of nitrogen fertilization systems based on the in-season variability of the nitrogenous growth factors and soil fertility factors—a case of winter oilseed rape (Brassica napus L.). Agronomy 2020, 10, 1701.
- Richards, R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 337–458.
- Szczepaniak, W. A mineral profile of oilseed rape in critical stages of growth—nitrogen. J. Elem. 2014, 19, 759–778.
- Grzebisz, W.; Szczepaniak, W.; Grześ, S. Sources of nutrients for high-yielding winter oilseed rape (Brassica napus L.) during post-flowering growth. Agronomy 2020, 10, 626.
- Spiertz, J.; Vos, J. Grain Growth of Wheat and Its Limitation by Carbohydrate and Nitrogen Supply. In Wheat Growth and Model-ling; Day, W., Atkin, R., Eds.; Plenum Press, New York, NY, USA, 1985.
- Barraclough, P.B.; Lopez-Bellido, R.; Hawkesford, M. Genotypic variation in the uptake, partitioning and remobilization of nitrogen during grain-filling in wheat. Field Crops Res. 2014, 156, 242–248.
- Taulemesse, F.; Le Gouis, J.; Gouache, D.; Gibon, Y.; Allard, V. Bread wheat (Triticum aestivum L.) grain protein concentration is related to early post-flowering nitrate uptake under putative control of plant satiety level. PLoS ONE 2016, 11, e0149668.
- Bogard, M.; Allard, V.; Brancourt-Hulmel, M; Heumez, E.; Machet, J-M.; Jeufforoy, M-H.; Gate, Ph.; Le Gouis, J. Deviation from the grain protein concentration—grain yield negative relationships is highly correlated to post-anthesis N uptake by winter wheat. J. Exp. Bot. 2010, 15, 4303–4312.
- Subedi, K.; Ma, B. Nitrogen uptake and partitioning in stay-green and leafy maize hybrids. Crops Sci. 2005, 45, 740–747.
- Grzebisz, W.; Wrońska, M.; Diatta, J.B.; Szczepaniak, W. Effect of zinc application at early stages of maize growth on the patterns of nutrients and dry matter accumulation by canopy. Part, I. Nitrogen uptake and dry matter accumulation patterns. J. Elem. 2008, 13, 17–28.
- Kovacs, P.; Vyn, T.J. Relationships between earl-leaf nutrient concentration at silking and corn biomass and grain yield at maturity. Agron. J. 2017, 109, 2898–2906.
- Szczepaniak, W.; Barłóg, P.; Łukowiak, R.; Przygocka-Cyna, K. Effect of balanced nitrogen fertilization in four-year rotation on plant productivity. J. Central Europ. Agroc. 2013, 14, 64–77.
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J.; Łukowiak, R.; Przygocka-Cyna, K. Nutritional status of winter oilseed rape in cardinal stages of growth as yield indicator. Plant Soil Environ. 2015, 61, 291–296.
- Weih, M.; Hamnér, K.; Pourazari, F. Analyzing plant nutrient uptake and utilization efficiencies: Comparison between crops and approaches. Plant Soil 2018, 430, 7–21.
- White, P.J.; Brown. P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080.
- Tan, Z.X.; Lal, R.; Wiebe, K.D., Global soil nutrient depletion and yield reduction, J. Sustain. Develop. 2015, 26(1), 123-146.
- Mi, G.-H.; Chen, F.-J.; Wu, Q-P.; Lai N-W.; Yuan L-X. Ideotype root architecture for effcient nitrogen acqusition by maize in intensive cropping systems. Sci. China Life Sci. 2010, 553, 1369–1373.
- Ruiz Herrera, L.F.; Shane, M.W.; López-Bucio, J. Nutritional regulation of root development. WIREs Dev. Biol. 2015, 4, 431–443.
- Lynch, J.P. Root phenothypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564.
- Duque, L.O.; Villordon, A. Root branching and nutrient efficiency: Status and way forward in root and tuber crops. Front. Plant Sci. 2019. 10:237.
- Fageria, N.K.; Moreira, A. The role of mineral nutrition on root growth of crop plants. Adv. Agron. 2011, 110, 3–83.