Quorum sensing is a type of chemical communication by which bacterial populations control expression of their genes in a coordinated manner. This regulatory mechanism is commonly used by pathogens to control the expression of genes encoding virulence factors and that of genes involved in the bacterial adaptation to variations in environmental conditions. In phytopathogenic bacteria, several mechanisms of quorum sensing have been characterized. In this review, we describe the different quorum sensing systems present in phytopathogenic bacteria, such as those using the signal molecules named N-acyl-homoserine lactone (AHL), diffusible signal factor (DSF), and the unknown signal molecule of the virulence factor modulating (VFM) system. We focus on studies performed on phytopathogenic bacteria of major importance, including Pseudomonas, Ralstonia, Agrobacterium, Xanthomonas, Erwinia, Xylella,Dickeya, and Pectobacterium spp. For each system, we present the mechanism of regulation, the functions targeted by the quorum sensing system, and the mechanisms by which quorum sensing is regulated.
1. Introduction
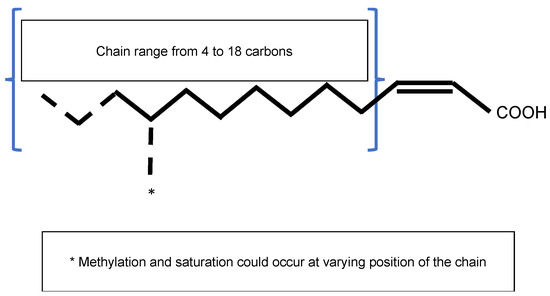
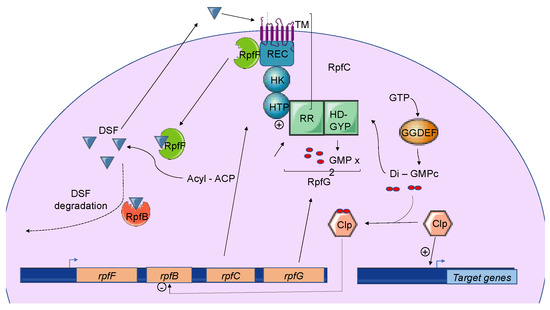
Quorum sensing (QS) is a cell-to-cell communication mechanism used by bacteria for promoting collective behavior within a population. This cooperative behavior relies on the production, detection, and response to signal molecules in a cell-density-dependent manner. At a low cell density, a basal level of the signal molecule is produced by bacteria. Signal molecules can be diffused or exported into the extracellular environment. As bacterial density increases, signal molecules accumulate. After reaching a threshold, signal molecules are perceived by the bacteria, which initiate a set of biological activities in a coordinated fashion. Acyl-homoserine lactone (AHL) was the first signal molecule, identified in the 1980s [
1,
2]. Originally discovered in the bioluminescent marine bacterium
Aliivibrio fischneri, these signal molecules were later characterized in a plethora of bacteria including
Pectobacterium carotovorum (formerly named
Erwinia),
Agrobacterium tumefaciens,
Citrobacter amalonaticus, and
Pseudomonas aeruginosa [
3,
4]. Since then, several other types of QS signals have been identified, and most QS signals are either small organic molecules or peptides with 5 to 20 amino acids. In Gram-positive bacteria, the signal molecules are mainly peptides [
5], while in Gram-negative bacteria, they are organic molecules smaller than 1000 Da. A universal signal described as autoinducer 2 (AI-2) is also produced by some Gram-positive and Gram-negative bacteria. These signal molecules are produced (i) at a specific growth stage, (ii) under particular physiological conditions, or (iii) in response to an environmental change. QS controls the expression of the many genes involved in a variety of functions, such as biofilm formation, toxin production, exopolysaccharide synthesis, extracellular enzyme production, motility, and plasmid conjugation. In pathogenic bacteria and, therefore, in plant pathogenic bacteria, QS plays a major role in the regulation of virulence factor productions and the infectious processes.
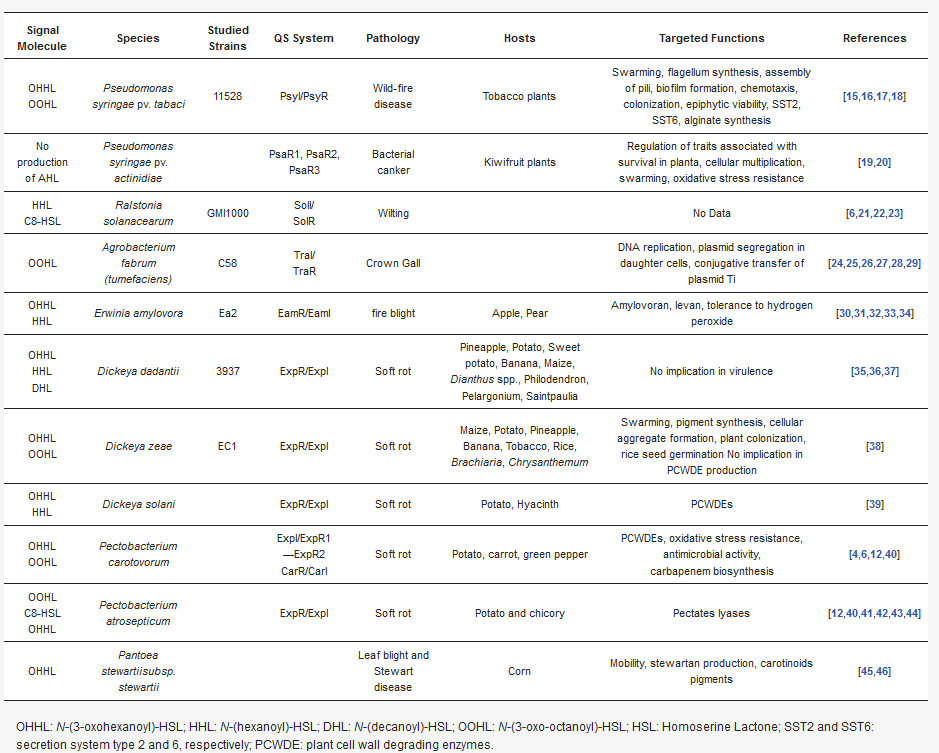
This entry aims to describe how phytopathogenic bacteria incorporate QS mechanisms into the complex regulatory cascades that control genes in pathogenicity and colonization of the host, and thereby update the data reviewed more than 15 years ago in Von Bodman et al. [
6]. We present QS systems harbored by phytopathogenic bacteria, i.e., the ones relying on AHL or diffusible signal factors (DSF), in addition to the virulence factor modulating (VFM) system. For each of these systems, we present the regulatory mechanism, the target genes of QS, and the mechanisms that are involved in the QS process. Mansfield et al. previously listed 10 species of phytopathogenic bacteria of major economic and scientific importance [
7]. Here, we focus on the QS systems present in these species, including
Pseudomonas syringae,
Ralstonia solanacearum,
Agrobacterium tumefaciens spp., bacteria of the genus
Xanthomonas spp.,
Erwinia amylovora,
Xylella fastidiosa,
Dickeya spp., and
Pectobacterium spp. ().
Table 1. Quorum screening (QS) systems present in bacterial plant pathogen species. The table presents a ranked list of the bacteria according to Mansfield et al.
| Top 10 Rank [7] |
Bacterial Pathogen Species |
QS Mechanisms |
Involvement in Virulence |
| 1 |
Pseudomonas syringae |
AHL |
Yes |
| 2 |
Ralstonia solanacearum |
AHL |
No |
| DSF-derived signals |
Yes |
| 3 |
Agrobacterium tumefaciens with pTi |
AHL |
Yes |
| 4 |
Xanthomonas oryzae pv oryzae |
DSF |
Yes |
| 5 |
Xanthomonas campestris pv |
DSF |
Yes |
| 6 |
Xanthomonas axonopodis pv |
DSF |
Yes |
| 7 |
Erwinia amylovora |
AHL |
Yes * |
| 8 |
Xylella fastidiosa |
DSF |
Yes |
| 9 |
Dickeya spp. |
AHL |
No ** |
| Vfm |
Yes |
| 10 |
Pectobacterium carotovorum (and atrosepticum) |
AHL |
Yes |
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms9020239