Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is causing the current pandemic of coronavirus disease 2019 (COVID-19), and COVID-19 vaccines focus on its spike protein. However, in addition to facilitating the membrane fusion and viral entry, the SARS-CoV-2 spike protein promotes cell growth signaling in human lung vascular cells, and patients who have died of COVID-19 have thickened pulmonary vascular walls, linking the spike protein to a fatal disease, pulmonary arterial hypertension (PAH).
- cell signaling
- coronavirus
- COVID-19
- lung
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is causing the current pandemic of coronavirus disease 2019 (COVID-19) [1][2]. To date, 100 million people have been infected with this virus, and COVID-19 has killed 2 million patients worldwide. In addition to the serious health problems, the pandemic has affected people economically, as well as sociologically, and the world awaits the means to end this horrible situation.
As a respiratory virus, the SARS-CoV-2 infection promotes severe pneumonia and acute respiratory distress syndrome [3]. One characteristic feature of the SARS-CoV-2 infection is, however, that elderly patients with pre-existing cardiovascular diseases are particularly susceptible to developing severe conditions and, ultimately, suffering death, while younger individuals only exhibit mild symptoms or are even asymptomatic when infected [2][4]. This feature of SARS-CoV-2 distinguishes it from other respiratory viruses and may serve as a basis for its profound effects. Thus, understanding these aspects of the disease could be crucial to developing therapeutic strategies to treat COVID-19.
Viruses use their membrane fusion proteins to bind to host cell receptors to facilitate entry into the cells through the process of membrane fusion and release the viral genomes [5]. The viral membrane fusion protein of SARS-CoV-2 is the spike protein, which is a Class I viral membrane fusion protein [2][6]. The host cell receptor of the SARS-CoV-2 spike protein for viral membrane fusion has been identified as angiotensin-converting enzyme 2 (ACE2) [7]. In humans, ACE2 (EC 3.4.17.23) normally functions as a peptidase enzyme that cleaves angiotensin II into angiotensin (1-7) [8]. Many cell types, including those in the lungs, arteries, heart, kidneys, and intestines, express ACE2 [9].
The SARS-CoV-2 spike protein consists of two subunits: Subunit 1 (S1) that contains the ACE2 receptor-binding domain (RBD) and Subunit 2 (S2) that participates in viral cell membrane fusion [3,6]. Since it is a well-conserved and exposed region of the virus, the spike protein has been used as the molecule to acquire immunity in the COVID-19 vaccines.
In addition to facilitating the viral entry and serving as the antigen for the vaccines, the SARS-CoV-2 spike protein elicits cell signaling in human host cells without the rest of the virus [10]. The SARS-CoV-2 spike protein is potent in affecting human pulmonary artery smooth muscle cells (PASMCs) and human pulmonary artery endothelial cells; pM levels of the recombinant protein were able to activate cell signaling. Thus, the spike protein of this virus can influence the host cells, and some of these actions could cause the pathogenesis of some diseases (Figure 1).
Figure 1. Actions of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike protein. In addition to facilitating SARS-CoV-2 entry into host cells and serving as the basis for the vaccine development, the spike protein of this virus also influences the host cells by activating cell signaling. Spike protein-mediated cell signaling could have adverse consequences.
2. The SARS-CoV-2 Spike Protein Activates Cell Signaling in Lung Vascular Cells
In the studies published by Suzuki et al. [10], cultured human PASMCs were treated with the recombinant full-length S1 subunit (Val16—Gln690) of the SARS-CoV-2 spike protein. As shown in Figure 2, the full-length S1 subunit of SARS-CoV-2 at a concentration as low as 130 pM strongly activated the phosphorylation of a mitogen-activated protein kinase kinase (MEK) at the Ser217 and Ser221 residues [10]. The kinetics of MEK phosphorylation promoted by the SARS-CoV-2 spike protein were consistently found to be transient, with a peak at 10 min. This fast activation suggests that it may be a receptor-mediated cell-signaling event. The recombinant SARS-CoV-2 spike protein also activated MEK in human pulmonary artery endothelial cells [10].
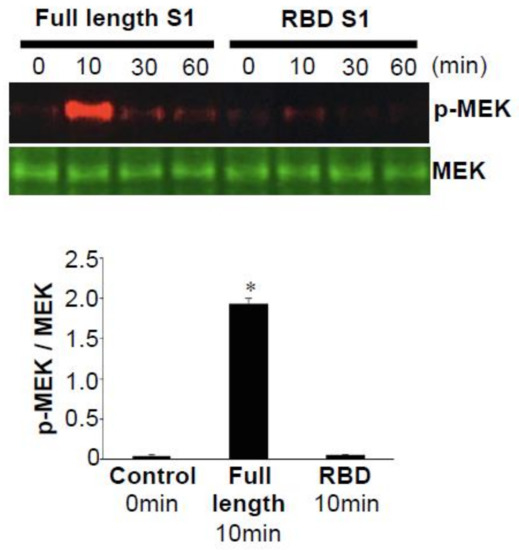
Figure 2. Full-length SARS-CoV-2 spike protein subunit 1 (S1), but not the receptor-binding domain (RBD)-only containing S1 protein, activates mitogen-activated protein kinase kinase (MEK) phosphorylation. Human pulmonary artery smooth muscle cells (PASMCs) were treated with the recombinant full-length S1 subunit (Val16—Gln690) or the RBD region of the S1 subunit (Arg319—Phe541). Cell lysates were prepared and subjected to Western blotting using antibodies against phosphorylated MEK (p-MEK) and the MEK protein. The bar graph represents means ± SEM. * Significantly different from the 0-min control at p < 0.05 (Taken from Suzuki et al. [10]).
These results demonstrate that the SARS-CoV-2 spike protein without the rest of the virus can elicit cell signaling—specifically, the activation of the MEK/extracellular signal-regulated kinase (ERK) pathway—in human lung vascular smooth muscle and endothelial cells. The MEK/ERK pathway is a well-known cell growth mechanism [11]. Thus, this SARS-CoV-2 spike protein-mediated cell growth signaling may promote the thickening of pulmonary vessels in COVID-19. Consistently, we found that the pulmonary vessel walls of postmortem lung tissues from COVID-19 patients who died of acute respiratory distress syndrome were significantly thickened compared with those of H1N1 influenza patients who died of the same syndrome [10]. These results provide a possible link between the SARS-CoV-2 spike protein and PAH.
3. Pathology of PAH
PAH is a fatal disease without a cure that can affect both males and females of any age, including children [12][13]. It is a progressive disease, and by the time patients are diagnosed, the thickening of the pulmonary vascular walls has often already occurred. Increased resistance in the pulmonary circulation places strain on the right ventricle, which leads to right heart failure and death [12][13]. The median overall survival for patients with PAH is 2.8 years from the time of diagnosis (three-year survival: 48%) without treatment [14][15]. Even with currently available therapies, the prognosis remains poor, with a three-year survival of PAH patients reported to be only 58–75% [16][17][18][19].
4. COVID-19 Vaccines and PAH
COVID-19 vaccines currently under consideration, including RNA vaccines (BNT162b2 and mRNA-1273) [20][21][22], viral vector-based vaccines (AZD1222 and Ad26.COV2.S) [23][24], and recombinant protein (NVX-CoV2373) [25], all introduce the SARS-CoV-2 spike protein into the human body. Whether the spike protein elicits cell signaling in host cells and exerts adverse events such as promoting PAH is a question raised in response to the experimental results in cultured cells [10]. RNA and viral vector-based vaccines use human host cells to produce the spike protein; thus, the intracellular spike protein will be produced. The intracellular effects of this foreign molecule on human cells have not been defined.
BNT162b2 encodes the full-length spike protein of SARS-CoV-2 with two proline mutations [20][21], while its sister vaccine BNT162b1 encodes only the RBD of the SARS-CoV-2 spike protein, trimerized by the addition of a T4 fibritin foldon domain [26][27]. Our experiments also revealed that, in contrast to the full-length S1 subunit (Val16—Gln690) of the SARS-CoV-2 spike protein that activated cell signaling, the S1 RBD (Arg319—Phe541) did not elicit such events [10]. Figure 2 shows that, while the full-length S1 subunit strongly activated MEK, the S1 RBD did not activate MEK in human PASMCs [10]. Similarly, the full-length S1 subunit, but not the RBD-only containing S1 subunit, of the SARS-CoV-2 spike protein promoted the phosphorylation of MEK in human pulmonary artery endothelial cells [10]. Thus, the RBD-only protein such as the one encoded by the BNT162b1 vaccine may not impose the risk of developing PAH. Further work is needed to understand the effects of various SARS-CoV-2 spike protein segments on human host cells in order to develop the most efficacious and safe vaccines without long-term adverse consequences.
This entry is adapted from the peer-reviewed paper 10.3390/jor1010004
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269.
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506.
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422.
- Mohanty, S.K.; Satapathy, A.; Naidu, M.M.; Mukhopadhyay, S.; Sharma, S.; Barton, L.M.; Stroberg, E.; Duval, E.J.; Pradhan, D.; Tzankov, A.; et al. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) and coronavirus disease 19 (COVID-19)—Anatomic pathology perspective on current knowledge. Diagn. Pathol. 2020, 15, 103.
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and Mechanisms of Viral Membrane Fusion Proteins: Multiple Variations on a Common Theme. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 189–219.
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292.
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448.
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: Celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 2020, 126, 1456–1474.
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; Van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637.
- Suzuki, Y.J.; Nikolaienko, S.I.; Dibrova, V.A.; Dibrova, Y.V.; Vasylyk, V.M.; Novikov, M.Y.; Shults, N.V.; Gychka, S.G. SARS-CoV-2 spike protein-mediated cell signaling in lung vascular cells. Vasc. Pharmacol. 2020, 106823.
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18.
- Delcroix, M.; Naeije, R. Optimising the management of pulmonary arterial hypertension patients: Emergency treatments. Eur. Respir. Rev. 2010, 19, 204–211.
- McLaughlin, V.V.; Shah, S.J.; Souza, R.; Humbert, M. Management of Pulmonary Arterial Hypertension. J. Am. Coll. Cardiol. 2015, 65, 1976–1997.
- D’Alonzo, G.E.; Barst, R.J.; Ayres, S.M.; Bergofsky, E.H.; Brundage, B.H.; Detre, K.M.; Fishman, A.P.; Goldring, R.M.; Groves, B.M.; Kernis, J.T.; et al. Survival in Patients with Primary Pulmonary Hypertension. Results from a national prospective registry. Ann. Intern. Med. 1991, 115, 343–349.
- Runo, J.R.; Loyd, J.E. Primary pulmonary hypertension. Lancet 2003, 361, 1533–1544.
- Benza, R.L.; Miller, D.P.; Frost, A.; Barst, R.J.; Krichman, A.M.; McGoon, M.D. Analysis of the Lung Allocation Score Estimation of Risk of Death in Patients with Pulmonary Arterial Hypertension Using Data From the REVEAL Registry. Transplantation 2010, 90, 298–305.
- Humbert, M.; Sitbon, O.; Yaici, A.; Montani, D.; O’Callaghan, D.S.; Jais, X.; Parent, F.; Savale, L.; Natali, D.; Gunther, S.; et al. Survival in incident and prevalent cohorts of patients with pulmonary arterial hypertension. Eur. Respir. J. 2010, 36, 549–555.
- Thenappan, T.; Shah, S.J.; Rich, S.S.; Tian, L.; Archer, S.L.; Gombergmaitland, M. Survival in pulmonary arterial hypertension: A reappraisal of the NIH risk stratification equation. Eur. Respir. J. 2009, 35, 1079–1087.
- Olsson, K.M.; Delcroix, M.; Ghofrani, H.A.; Tiede, H.; Huscher, D.; Speich, R.; Grünig, E.; Staehler, G.; Rosenkranz, S.; Halank, M.; et al. Anticoagulation and Survival in Pulmonary Arterial Hypertension: Results From the Comparative, Prospective Registry of Newly Initiated Therapies for Pulmonary Hypertension (COMPERA). Circulation 2014, 129, 57–65.
- Walsh, E.E.; Frenck, R.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. RNA-Based COVID-19 Vaccine BNT162b2 Selected for a Pivotal Efficacy Study. medRxiv 2020. Available online: https://www.medrxiv.org/content/10.1101/2020.08.17.20176651v2 (accessed on 28 August 2020).
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and immunogenicity of two RNA-based Covid-19 vaccine candidates. N. Engl. J. Med. 2020, 383, 2439–2450.
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA vaccine against SARS-CoV-2—Preliminary report. N. Engl. J. Med. 2020, 383, 1920–1931.
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 2020, 396, 467–478.
- Mercado, N.B.; Zahn, R.; Wegmann, F.; Loos, C.; Chandrashekar, A.; Yu, J.; Liu, J.; Peter, L.; McMahan, K.; Tostanoski, L.H.; et al. Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques. Nature 2020, 586, 583–588.
- Guebre-Xabier, M.; Patel, N.; Tian, J.-H.; Zhou, B.; Maciejewski, S.; Lam, K.; Portnoff, A.D.; Massare, M.J.; Frieman, M.B.; Piedra, P.A.; et al. NVX-CoV2373 vaccine protects cynomolgus macaque upper and lower airways against SARS-CoV-2 challenge. Vaccine 2020, 38, 7892–7896.
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593.
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and T H 1 T cell responses. Nature 2020, 586, 594–599.