Microalgal biomass is considered as a sustainable and renewable feedstock for biofuel production (biohydrogen, biomethane, biodiesel) characterized by lower emissions of hazardous air pollutants than fossil fuels. Photobioreactors for microalgae growth can be exploited using many industrial and domestic wastes. It allows locating the commercial microalgal systems in areas that cannot be employed for agricultural purposes, i.e., near heating or wastewater treatment plants and other industrial facilities producing carbon dioxide and organic and nutrient compounds.
- microalgal biomass
- microalgae cultivation
- biofuels
1. Introduction
|
Product |
Use |
Ref. |
|
|---|---|---|---|
|
Agar |
Food ingredient, fruit preserves, hydrocolloids, clarifying brewing agent, paper industry, and others |
[13] |
|
|
Alginate |
Food additive, medical, pharmaceutical, paper, cosmetic and fertilizer industries, textile printing |
[1] |
|
|
Antioxidants |
Preservatives in cosmetic, chemical, food, and pharmaceutical industries |
[14] |
|
|
Astaxanthin |
Food supplement as food dye additive and antioxidant |
[15] |
|
|
Beta-carotene and carotenoids |
Precursor for vitamin A and supplement for vitamin C, food additive as coloring agent, and antioxidant |
[16] |
|
|
Bioenergy and biofuels |
Biodiesel, bioethanol, biogas, biohydrogen, biomethane, aviation gas, biobutanol, biosyngas, bio-oil, gasoline, solid fuel, jet fuel |
[15] |
|
|
Biochar |
Agricultural and sorbent uses, combustion |
[13] |
|
|
Biorefinery |
Various chemicals and biofuels |
[14] |
|
|
Biosorbents |
Ion exchange materials that bind strongly heavy metal ions |
[1] |
|
|
Carragen or carrageenan |
Pet food, food additive, gels, toothpaste |
[1] |
|
|
Catalysts |
Catalytic properties |
[14] |
|
|
Chemicals |
Industrial and medicinal uses |
[1] |
|
|
Conditioners |
Chemical, cosmetic, and farming industries |
[15] |
|
|
Digester residue |
Compost or vermicompost |
[13] |
|
|
Extraction of |
hydrocolloids or gums |
Food industry, phytocolloids such as agar, alginate, and carrageenan |
[14] |
|
lipids carbohydrates starch and cellulose |
Biogas, biodiesel, gasoline, jet fuel, alcohols, renewable hydrocarbons |
||
|
minerals and trace elements |
Food supplements, glass production, metallurgy |
||
|
proteins |
Fertilizers, industrial enzymes, animal/fish feeds, surfactants, bioplastics |
||
|
Feed |
Animal food |
[15] |
|
|
Fertilizers |
N-, P-, and K-rich fertilizers |
[1] |
|
|
Phytosterol |
Food supplements |
[15] |
|
|
Pigments |
Natural colorants in paper and textile industries |
[14] |
|
|
Production |
Cosmetic |
Water-binding agents and antioxidants, “skin foods” |
[15] |
|
Food and drink |
Nori, kombu, wakame, cheese, soup, noodles, pasta, wine, tea, others |
||
|
Fruit and vegetable preservatives |
Food industry |
||
|
Glass |
Glass industry |
||
|
Paper pulp supplements |
Paper industry |
||
|
Textile |
Textile industry |
||
|
Therapeutic materials |
Pharmaceutical industry |
||
|
Sector |
Use |
Ref. |
|
|---|---|---|---|
|
Wastewater treatment |
Nitrogen and phosphorus removal from municipal wastewater |
[17] |
|
|
Biodegradation of sparingly degradable pollutants |
|||
|
Treatment of organic wastewater |
[20] |
||
|
Treatment of hard-to-manage wastewater produced by |
timber and paper industry |
||
|
textile industry |
[23] |
||
|
phenol industry |
|||
|
Ethanol and citric acid production |
[26] |
||
|
Removal of heavy metals (copper, nickel, lead) from wastewater |
|||
|
Gas treatment |
Reducing emissions of carbon dioxide and other pollutants (nitrogen and sulfur oxides) from waste and exhaust gases |
||
|
Waste management |
Use of waste glycerol as a carbon source in heterotrophic cultivation |
[30] |
|
|
Microalgae cultivation using industrial waste or low-value feedstocks, such as |
breadcrumbs |
[31] |
|
|
brewer’s spent yeast |
[32] |
||
|
coconut water |
[33] |
||
|
empty palm fruit bunches |
[34] |
||
|
Leachate treatment |
Biodegradation of landfill leachates |
[35] |
|
|
Neutralization of degraded effluent from anaerobic fermentation of sewage sludge |
[36] |
||
|
Biogas upgrading |
Biological sequestration of CO2 with photosynthetic microalgae (photosynthesis allows producing biogas with 94% methane content) |
[37] |
|
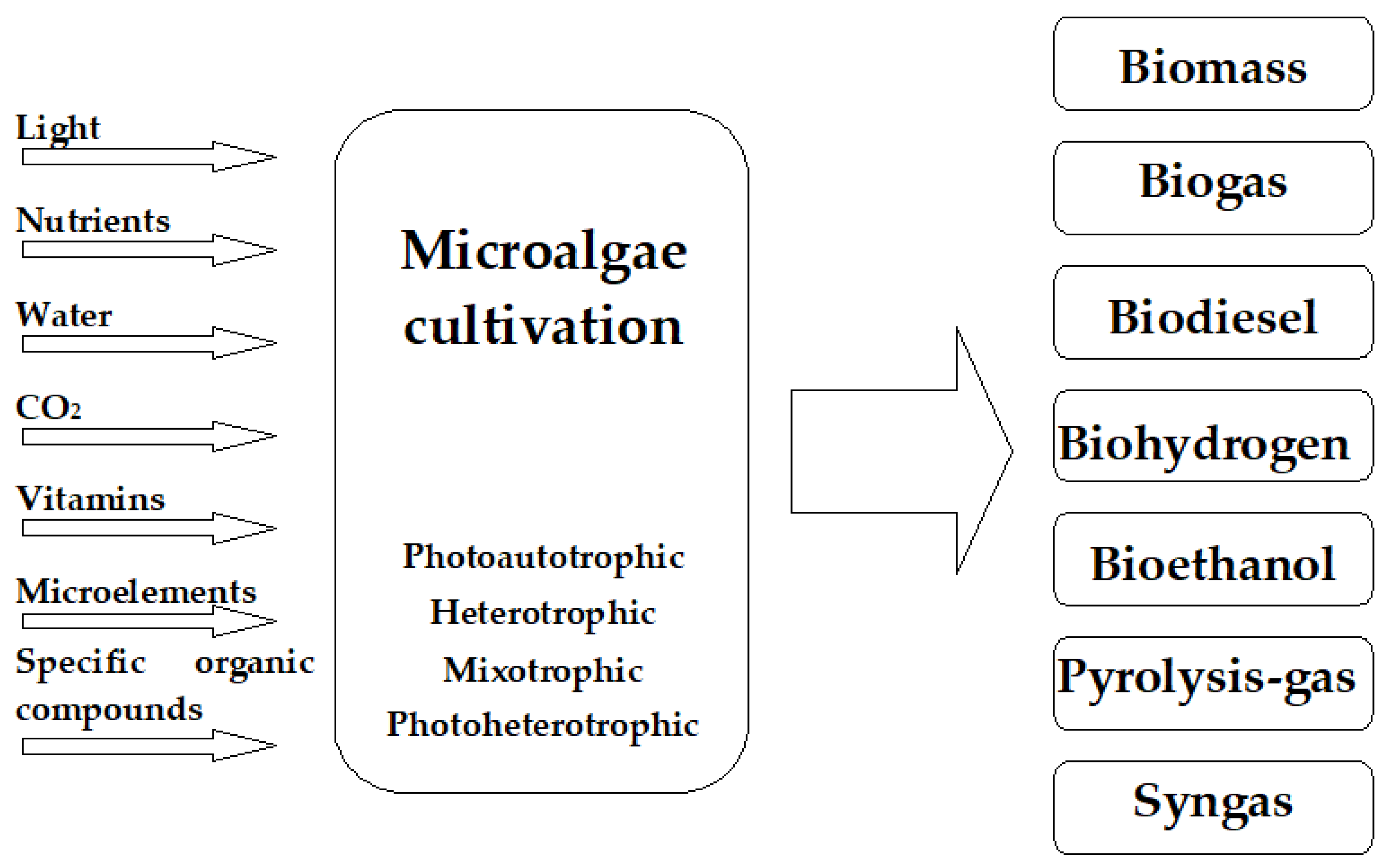
2. Microalgal Biomass as a Source of Biofuels
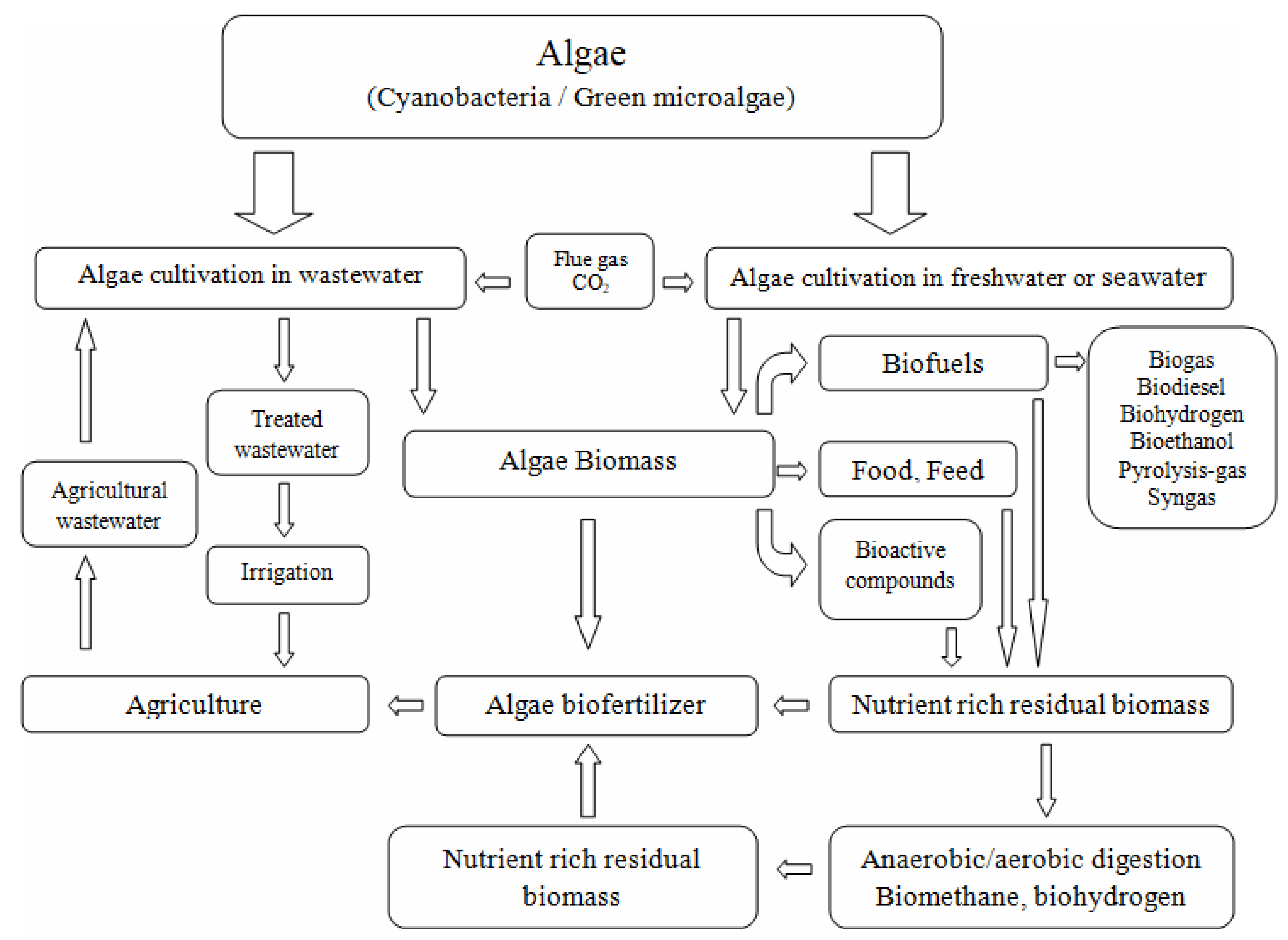
3. Systems of Microalgae Species Cultivation for Biofuel
This entry is adapted from the peer-reviewed paper 10.3390/su12239980
References
- Deviram, G.; Mathimani, T.; Anto, S.; Ahamed, T.S.; Ananth, D.A.; Pugazhendhi, A. Applications of microalgal and cyanobacterial biomass on a way to safe, cleaner and a sustainable environment. J. Clean. Prod. 2020, 253, 119770.
- Kamani, M.H.; Eş, I.; Lorenzo, J.M.; Remize, F.; Roselló-Soto, E.; Barba, F.J.; Clark, J.H.; Khaneghah, A.M. Advances in plant materials, food by-products, and algae conversion into biofuels: Use of environmentally friendly technologies. Green Chem. 2019, 21, 3213–3231.
- Patil, S.; Prakash, G.; Lali, A.M. Reduced chlorophyll antenna mutants of Chlorella saccharophila for higher photosynthetic efficiency and biomass productivity under high light intensities. J. Appl. Phycol. 2020, 32, 1559–1567.
- Santhakumaran, P.; Ayyappan, S.M.; Ray, J.G. Nutraceutical applications of twenty-five species of rapid-growing green-microalgae as indicated by their antibacterial, antioxidant and mineral content. Algal Res. 2020, 47, 101878.
- Tolboom, S.N.; Carrillo-Nieves, D.; de Jesús Rostro-Alanis, M.; de la Cruz Quiroz, R.; Barceló, D.; Iqbal, H.M.N.; Parra-Saldivar, R. Algal-based removal strategies for hazardous contaminants from the environment—A review. Sci. Total. Environ. 2019, 665, 358–366.
- Ziolkowska, J.R. Chapter 1—Biofuels technologies: An overview of feedstocks, processes, and technologies. In Biofuels for a More Sustainable Future; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2020; pp. 1–19.
- Stiles, W.A.V.; Styles, D.; Chapman, S.P.; Esteves, S.; Bywater, A.; Melville, L.; Silkina, A.; Lupatsch, I.; Fuentes, C.; Lovitt, R.; et al. Using microalgae in the circular economy to valorise anaerobic digestate: Challenges and opportunities. Bioresour. Technol. 2018, 267, 732–742.
- SundarRajan, P.; Gopinath, K.P.; Greetham, D.; Antonysamy, A.J. A review on cleaner production of biofuel feedstock from integrated CO2 sequestration and wastewater treatment system. J. Clean. Prod. 2019, 210, 445–458.
- Nawaz, T.; Rahman, A.; Pan, S.; Dixon, K.; Petri, B.; Selvaratnam, T. A review of landfill leachate treatment by microalgae: Current status and future directions. Processes 2020, 8, 384.
- Nagarajan, D.; Lee, D.J.; Chang, J.S. Biogas Upgrading by Microalgae: Strategies and Future Perspectives. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Alam, M., Wang, Z., Eds.; Springer: Singapore, 2019.
- Rahman, A.; Agrawal, S.; Nawaz, T.; Pan, S.; Selvaratnam, T. A review of algae-based produced water treatment for biomass and biofuel production. Water 2020, 12. 2351.
- Sahu, S.K.; Mantri, V.A.; Zheng, P.; Yao, N. Chapter 1 Algae Biotechnology. Current Status, Potential and Impediments. In Encyclopedia of Marine Biotechnology; John Wiley & Sons Ltd: Hoboken, NJ, USA, 2020.
- Vassilev, S.V.; Vassileva, C.G. Composition, properties and challenges of algae biomass for biofuel application: An overview. Fuel 2016, 181, 1–33.
- Barsanti, L.; Gualtieri, P. Is exploitation of microalgae economically and energetically sustainable? Algal Res. 2018, 31, 107–115.
- Bhalamurugan, G.L.; Valerie, O.; Mark, L. Valuable bioproducts obtained from microalgal biomass and their commercial applications: A review. Environ. Eng. Res. 2018, 23, 229–241.
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal Biomass Util. 2012, 3, 89–100.
- Gomez Villa, H.; Voltolina, D.; Nieves, M.; Pina, P. Biomass production and nutrient budget in outdoor cultures of Scenedesmus obliquus (chlorophyceae) in artificial wastewater, under the winter and summer conditions of Mazatla´ n, Sinaloa, Mexico. Vie Et Milieu 2005, 55, 121–126.
- Mùnoz, R.; Guieysse, B. Algal-bacterial processes for the treatment of hazardous contaminants: A review. Water Res. 2006, 40, 2799–2815.
- Chojnacka, K.; Chojnacki, A.; Gorecka, H. Biosorption of Cr3+, Cd2+ and Cu2+ ions by blue–green algae Spirulina sp.: Kinetics, equilibrium and the mechanism of the process. Chemosphere 2005, 59, 75–84.
- Mùnoz, R.; Köllner, C.; Guieysse, B. Biofilm photobioreactors for the treatment of industrial wastewaters. J. Hazard. Mater. 2009, 161, 29–34.
- Yewalkar, S.N.; Dhumal, K.N.; Sainis, J.K. Chromium (VI)-reducing Chlorella spp. isolated from disposal sites of paper-pulp and electroplating industry. J. Appl. Phycol. 2007, 19, 459–465.
- Tarlan, E.; Dilek, F.B.; Yetis, U. Effectiveness of algae in the treatment of a wood-based pulp and paper industry wastewater. Bioresour. Technol. 2002, 84, 1–5.
- Acuner, E.; Dilek, F.B. Treatment of tectilon yellow 2G by Chlorella vulgaris. Process. Biochem. 2004, 39, 623–631.
- Essam, T.; Magdy, A.A.; El Tayeb, O.; Mattiasson, B.; Guieysse, B. Solar-based detoxification of phenol and p-nitrophenol by sequential TiO2 photocatalysis and photosynthetically aerated biological treatment. Water Res. 2007, 41, 1697–1704.
- Lima, S.A.C.; Raposo, M.F.J.; Castro, P.M.L.; Morais, R.M. Biodegradation of p-chlorophenol by a microalgae consortium. Water Res. 2004, 38, 97–102.
- Valderramaa, L.T.; Del Campoa, C.M.; Rodrigueza, C.M.; de-Bashan, L.E.; Bashan, Y. Treatment of recalcitrant wastewater from ethanol and citric acid production using the microalga Chlorella vulgaris and the macrophyte Lemna minuscule. Water Res. 2002, 36, 4185–4192.
- Tien, C.J. Biosorption of metal ions by freshwater algae with different surface characteristics. Process. Biochem. 2002, 38, 605–613.
- Jacob-Lopes, E.; Scoparo, C.H.G.; Queiroz, M.I.; Franco, T.T. Biotransformations of carbon dioxide in photobioreactors. Energy Convers. Manag. 2010, 51, 894–900.
- De Morais, M.G.; Costa, J.A.V. Isolation and selection of microalgae from coal fired thermoelectric power plant for biofixation of carbon dioxide. Energy Convers. Manag. 2007, 48, 2169–2173.
- Lam, M.K.; Lee, K.T.; Rahman, M.A. Current status and challenges on microalgae-based carbon capture. Int. J. Greenh. Gas Control. 2012, 10, 456–469.
- Thyagarajan, T.; Puri, M.; Vongsvivut, J.; Barrow, C.J. Evaluation of Bread Crumbs as a Potential Carbon Source for the Growth of Thraustochytrid Species for Oil and Omega-3 Production. Nutrients 2014, 6, 2104–2114.
- Ryu, B.; Kim, K.; Kim, J.; Han, J.; Yang, J. Use of organic waste from brewery industry for high-density cultivation of docosahexaenoic acid-rich microalga Aurantochytrium sp. KRS101. Bioresour. Technol. 2012, 129, 351–359.
- Unagul, P.; Assantachai, C.; Phadungruengluij, S.; Suphantharika, M.; Tanticharoen, M.; Verduyn, C. Coconut water as a medium additive for the production of docosahexaenoic acid (C22:6 n3) by Schizochytrium mangrovei Sk-02. Bioresour. Technol. 2007, 98, 281–287.
- Hong, W.; Yu, A.; Heo, S.; Oh, B.; Kim, C.; Sohn, J.; Yang, J.W.; Kondo, A.; Seo, J.W. Production of lipids containing high levels of docosahexaenoic acid from empty palm fruit bunches by Aurantiochytrium sp. KRS101. Bioprocess. Biosyst. Eng. 2013, 36, 959–963.
- Lin, L.; Chan, G.Y.S.; Jiang, B.L.; Lan, C.Y. Use of ammoniacal nitrogen tolerant microalgae in landfill leachate treatment. Waste Manag. 2007, 27, 1376–1382.
- Jedynak, P.; Burczyk, J.; Borowski, S.; Kaszycki, P.; Hałat-Łaś, M.; Kędra, M.; Mungunkhuyag, K.; Malec, P. Use of microalgae for treatment of post-fermentation effluent from biogas production. Przemysł Chem. 2018, 97, 2106–2109.
- Yan, C.; Zheng, Z. Performance of photoperiod and light intensity on biogas upgrade and biogas effluent nutrient reduction by the microalgae Chlorella sp. Bioresour. Technol. 2013, 139, 292–299.
- Kumar, M.; Sun, Y.; Rathour, R.; Pandey, A.; Thakur, I.S.; Tsang, D.C.W. Algae as potential feedstock for the production of biofuels and value-added products: Opportunities and challenges. Sci. Total. Environ. 2020, 716, 137116.
- Callegari, A.; Bolognesi, S.; Cecconet, D.; Capodaglio, A.G. Production technologies, current role, and future prospects of biofuels feedstocks: A state-of-the-art review. J. Crit. Rev. Environ. Sci. Technol. 2020, 50, 384–436.
- Walsh, M.J.; van Gerber Doren, L.; Sills, D.L.; Archibald, I.; Beal, C.M.; Lei, X.G.; Greene, C.H. Algal food and fuel coproduction can mitigate greenhouse gas emissions while improving land and water-use efficiency. Environ. Res. Lett. 2016, 11, 114006.
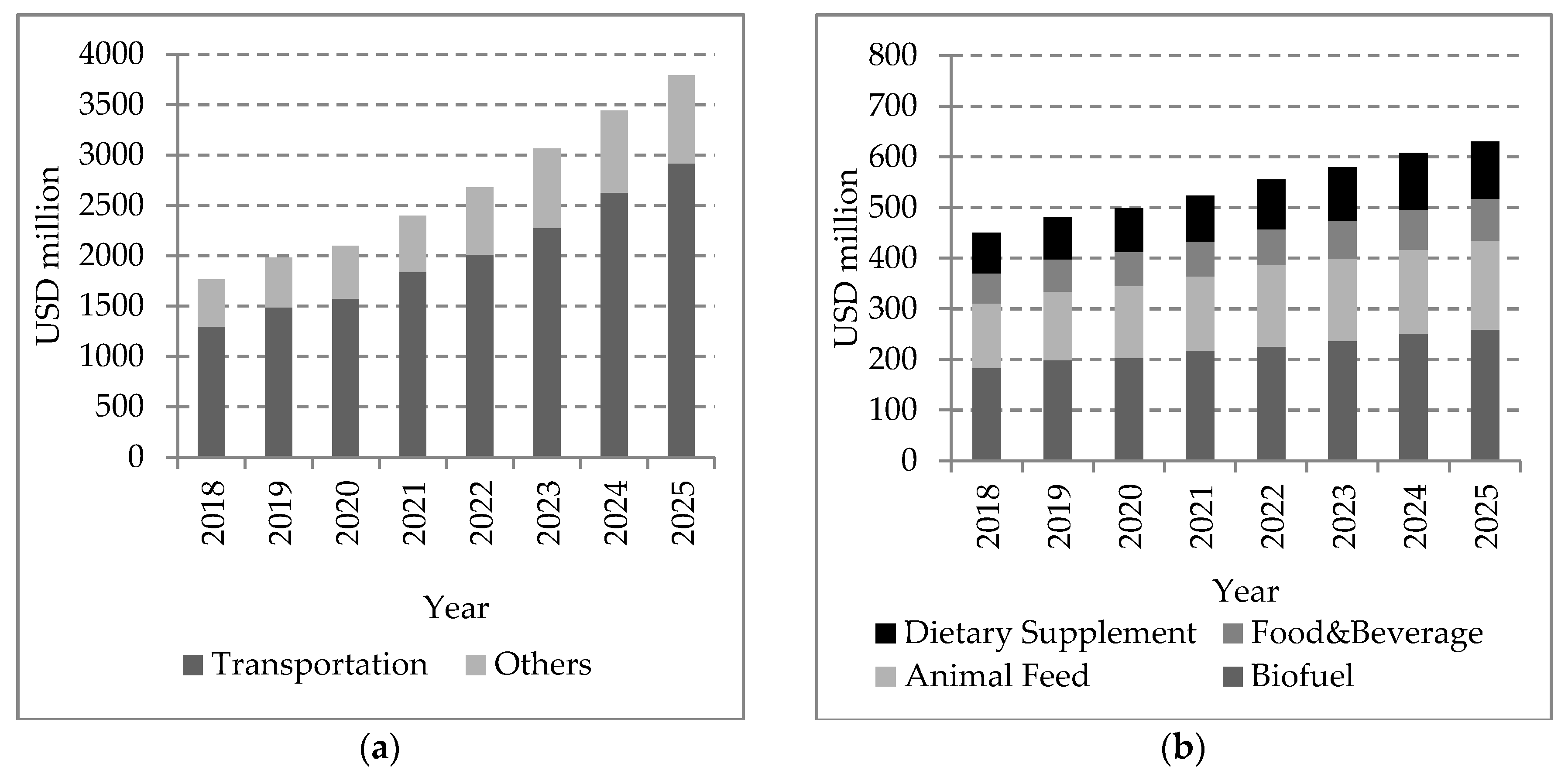
- Market Analysis Report. Algae Biofuel Market Size, Share & Trend Analysis By Application (Transportation, Others), By Region (North America, Europe, Asia Pacific, ROW), By Country, And Segment Forecasts, 2018–2025. Grand View Research. 2017. Available online: https://www.grandviewresearch.com/industry-analysis/algae-biofuel-market (accessed on 4 August 2020).
- Market Analysis Report. Biodiesel Market Analysis by Feedstock , By Application (Fuel, Power Generation), And Segment Forecasts, 2018–2025. Grand View Research. 2017. Available online: https://www.grandviewresearch.com/industry-analysis/biodiesel-market (accessed on 5 August 2020).
- Fu, W.; Nelson, D.R.; Mystikou, A.; Daakour, S.; Salehi-Ashtiani, K. Advances in microalgal research and engineering development. Curr. Opin. Biotechnol. 2019, 59, 157–164.
- Yadav, G.; Sen, R. Sustainability of Microalgal Biorefinery: Scope, Challenges, and Opportunities. In Sustainable Energy Technology and Policies; Green Energy and, Technology, De, S., Bandyopadhyay, S., Assadi, M., Mukherjee, D., Eds.; Springer: Singapore, 2018.
- Anto, S.; Mukherjee, S.S.; Muthappa, R.; Mathimani, T.; Deviram, G.; Kumar, S.S.; Verma, T.N.; Pugazhendhi, A. Algae as green energy reserve: Technological outlook on biofuel production. Chemosphere 2020, 242, 125079.
- Borowitzka, M.A.; Vonshak, A. Scaling up microalgal cultures to commercial scale. Eur. J. Phycol 2017, 52, 407–418.
- Webster, A.; Gardner, J. Aligning technology and institutional readiness: The adoption of innovation. Technology Anal. Strateg. Manag. 2019, 31, 1229–1241.
- Ozdemir, H.I.; Pinto, C.A.; Unal, R.; Keating, C.B.; Britcher, C. Supporting technology selection via portfolio readiness level and technology forecasting. In Proceedings of the International Annual Conference of the American Society for Engineering Management; Huntsville, Philadelphia, PA, USA, 24–26 October 2019.
- Bates, C.A.; Clausen, C. Engineering Readiness: How the TRL Figure of Merit Coordinates Technology Development. Eng. Stud. 2020, 12, 9–38.
- Okoro, V.; Azimov, U.; Munoz, J.; Hernandez, H.H.; Phan, A.N. Microalgae cultivation and harvesting: Growth performance and use of flocculants—A review. Renew. Sustain. Energy Rev. 2019, 115, 109364.
- Lardon, L.; He´lias, A.; Sialve, B.; Steyer, J.; Bernard, O. Life-cycle assessment of biodiesel production from microalgae. Environ. Sci. Technol. 2009, 43, 6475–6481.
- Clarens, A.F.; Nassau, H.; Resurreccion, E.P.; White, M.A.; Colosi, L.M. Environmental Impacts of Algae-Derived Biodiesel and Bioelectricity for Transportation. Environ. Sci. Technol. 2011, 45, 7554–7560.
- Frank, E.D.; Han, J.; Palou-Rivera, I.; Elgowainy, A.; Wang, M.Q. User Manual for algae life-cycle analysis with GREET: Version 0.0; ANL/ESD/11-7; Argonne National Laboratory: Lemont, IL, USA, 2011.
- Ganesan, R.; Manigandan, S.; Samuel, M.S.; Shanmuganathan, R.; Brindhadevi, K.; Chi, N.T.L.; Duc, P.A.; Pugazhendhi, A. A review on prospective production of biofuel from microalgae. Biotechnol. Rep. 2020, 27, e00509.
- Peng, L.; Fu, D.; Chu, H.; Wang, Z.; Qi, H. Biofuel production from microalgae: A review. Environ. Chem. Lett. 2020, 18, 285–297.
- Coimbra, R.N.; Escapa, C.; Otero, M. Comparative Thermogravimetric Assessment on the Combustion of Coal, Microalgae Biomass and Their Blend. Energies 2019, 12, 2962.
- Panahi, H.K.S.; Tabatabaei, M.; Aghbashlo, M.; Dehhaghi, M.; Rehan, M.; Nizami, A.-S. Recent updates on the production and upgrading of bio-crude oil from microalgae. Bioresour. Technol. Rep. 2019, 7, 100216.
- Feng, R.; Zaidi, A.A.; Zhang, K.; Shi, Y. Optimization of microwave pretreatment for biogas enhancement through anaerobic digestion of microalgal biomass. Period. Polytech. Chem. Eng. 2018, 63, 65–72.
- Córdova, O.; Chamy, R. Chapter 15—Microalgae to Biogas: Microbiological Communities Involved. In Microalgae Cultivation for Biofuels Production; Elsevier: London, UK; San Diego, CA, USA; Oxford, UK, 2020; pp. 227–249.
- Li, F.; Hülsey, M.J.; Yan, N.; Dai, Y.; Wang, C.-H. Co-transesterification of waste cooking oil, algal oil and dimethyl carbonate over sustainable nanoparticle catalysts. Chem. Eng. J. 2020, 405, 127036.
- Wang, Y.; Yang, H.; Zhang, X.; Han, F.; Tu, W.; Yang, W. Microalgal Hydrogen Production. Small Methods 2020, 4.
- Özçimen, D.; Koçer, A.T.; İnan, B.; Özer, T. Chapter 14—Bioethanol production from microalgae. In Handbook of Microalgae-Based Processes and Products; Elsevier: London, UK; San Diego, CA, USA; Cambridge, MA, USA; Oxford, UK, 2020; pp. 373–389.
- Chernova, N.I.; Kiseleva, S.V.; Larina, O.M.; Sytchev, G.A. Manufacturing gaseous products by pyrolysis of microalgal biomass. Int. J. Hydrog. Energy 2019, 45, 1569–1577.
- Lee, X.J.; Ong, H.C.; Gan, Y.Y.; Chen, W.-H.; Mahlia, T.M.I. State of art review on conventional and advanced pyrolysis of macroalgae and microalgae for biochar, bio-oil and bio-syngas production. Energy Convers. Manag. 2020, 210.
- Patil, R.A.; Kausley, S.B.; Joshi, S.M.; Pandit, A.B. Chapter 27—Process intensification applied to microalgae-based processes and products. In Handbook of Microalgae-Based Processes and Products; Elsevier: London, UK; San Diego, CA, USA; Cambridge, MA, USA; Oxford, UK, 2020; pp. 737–769.
- Nagappan, S.; Devendran, S.; Tsai, P.-C.; Dahms, H.-U.; Ponnusamy, V.K. Potential of two-stage cultivation in microalgae biofuel production. Fuel 2019, 252, 339–349.
- Fazal, T.; Mushtaq, A.; Rehman, F.; Khan, A.U.; Rashid, N.; Farooq, W.; Rehman, M.S.U.; Xu, J. Bioremediation of textile wastewater and successive biodiesel production using microalgae. Renew. Sustain. Energy Rev. 2018, 82, 3107–3126.
- Zhang, L.; Cheng, J.; Pei, H.; Pan, J.; Jiang, L.; Hou, Q. Cultivation of microalgae using anaerobically digested ef fl uent from kitchen waste as a nutrient source for biodiesel production. Renew. Energy 2018, 115, 276–287.
- Sati, H.; Mitra, M.; Mishra, S.; Baredar, P. Microalgal lipid extraction strategies for biodiesel production: A review. Algal Res. 2019, 38.
- Menegazzo, M.L.; Fonseca, G.G. Biomass recovery and lipid extraction processes for microalgae biofuels production: A review. Renew. Sustain. Energy Rev. 2019, 107, 87–107.
- Lee, S.Y.; Khoiroh, I.; Vo, D.N.; Kumar, S.; Loke, P. Show Techniques of lipid extraction from microalgae for biofuel production: A review. Environ. Chem. Lett. 2020, 1–21.
- Ranjith Kumar, R.; Hanumantha Rao, P.; Arumugam, M. Lipid extraction methods from microalgae: A comprehensive review. Front. Energy Res. 2015, 2, 61.
- Kumar, V.; Arora, N.; Nanda, M.; Pruthi, V. Different Cell Disruption and Lipid Extraction Methods from Microalgae for Biodiesel Production. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Alam, M., Wang, Z., Eds.; Springer: Singapore, 2019.
- Choudhary, P.; Assemany, P.P.; Naaz, F.; Bhattacharya, A.; Castro, J.S.; Couto, E.A.; Calijuri, M.L.; Pant, K.K.; Malik, A. A review of biochemical and thermochemical energy conversion routes of wastewater grown algal biomass. Sci. Total. Environ. 2020, 726, 137961.
- Ren, J.; Liu, Y.-L.; Zhao, X.-Y.; Cao, J.-P. Biomass thermochemical conversion: A review on tar elimination from biomass catalytic gasification. J. Energy Inst. 2020, 93, 1083–1098.
- Clark, J.; Deswarte, F. Introduction to chemicals from biomass. In Wiley Series in Renewable Resources; Stevens, C.V., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008.
- Radenahmad, N.; Azad, A.T.; Saghir, M.; Taweekun, J.; Bakar, M.S.A.; Reza, M.S.; Azad, A.K. A review on biomass derived syngas for SOFC based combined heat and power application. Renew. Sustain. Energy Rev. 2020, 119, 109560.
- Kousheshi, N.; Yari, M.; Paykani, A.; Saberi Mehr, A.; de la Fuente, G.F. Effect of Syngas Composition on the Combustion and Emissions Characteristics of a Syngas/Diesel RCCI Engine. Energies 2020, 13, 212.
- Hirano, A.; Hon-Nami, K.; Kunito, S.; Hada, M.; Ogushi, Y. Temperature effect on continuous gasification of microalgal biomass: Theoretical yield of methanol production and its energy balance. Catal. Today 1998, 45, 399–404.
- Minowa, T.; Sawayama, S. A novel microalgal system for energy production with nitrogen cycling. Fuel 1999, 78, 1213–1215.
- Patil, V.; Tran, K.-Q.; Giselrød, H.R. Towards sustainable production of biofuels from microalgae. Int. J. Mol. Sci. 2008, 9, 1188–1195.
- Ong, H.C.; Chen, W.-H.; Farooq, A.; Gan, Y.Y.; Lee, K.T.; Ashokkumar, V. Catalytic thermochemical conversion of biomass for biofuel production: A comprehensive review. Renew. Sustain. Energy Rev. 2019, 113, 109266.
- McKendry, P. Energy production from biomass (part 2): Conversion technologies. Bioresour. Technol. 2002, 83, 47–54.
- Dote, Y.; Sawayama, S.; Inoue, S.; Minowa, T.; Yokoyama, S.-Y. Recovery of liquid fuel from hydrocarbon-rich microalgae by thermochemical liquefaction. Fuel 1994, 73, 1855–1857.
- Minowa, T.; Yokoyama, S.-Y.; Kishimoto, M.; Okakura, T. Oil production from algal cells of Dunaliella tertiolecta by direct thermochemical liquefaction. Fuel 1995, 74, 1735–1738.
- Aravind, S.; Kumar, P.S.; Kumar, N.S.; Siddarth, N. Conversion of green algal biomass into bioenergy by pyrolysis. A review. Environ. Chem. Lett. 2020, 18, 829–849.
- Miao, X.L.; Wu, Q.Y. High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. J. Biotechnol. 2004, 110, 85–93.
- Miao, X.; Wu, Q.; Yang, C. Fast pyrolysis of microalgae to produce renewable fuels. J. Anal. Appl. Pyrolysis 2004, 71, 855–863.
- Demirbas, A. Oily products from mosses and algae via pyrolysis. Energy Sources Part A—Recover. Utilization Environ. Effects 2006, 28, 933–940.
- Li, F.; Srivatsa, S.C.; Bhattacharya, S. A review on catalytic pyrolysis of microalgae to high-quality bio-oil with low oxygeneous and nitrogenous compounds. Renew. Sustain. Energy Rev. 2019, 108, 481–497.
- Hirano, A.; Ueda, R.; Hirayama, S.; Ogushi, Y. CO2 fixation and ethanol production with microalgal photosynthesis and intracellular anaerobic fermentation. Energy 1997, 22, 137–142.
- Ueno, Y.; Kurano, N.; Miyachi, S. Ethanol production by dark fermentation in the marine green alga, Chlorococcum littorale. J. Ferment. Bioeng. 1998, 86, 38–43.
- Lee, S.Y.; Sankaran, R.; Chew, K.W.; Tan, C.H.; Krishnamoorthy, R.; Chu, D.-T.; Show, P.-L. Waste to bioenergy: A review on the recent conversion technologies. BMC Energy 2019, 1, 4.
- Ghirardi, M.L.; Zhang, L.; Lee, J.W.; Flynn, T.; Seibert, M.; Greenbaum, E.; Melis, A. Microalgae: A green source of renewable H2. Trends Biotechnol. 2000, 18, 506–511.
- Saratale, G.D.; Saratale, R.G.; Banu, J.R.; Chang, J.-S. Biohydrogen Production From Renewable Biomass Resources. In Biohydrogen; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–277.
- Veras, T.S.; Mozer, T.S.; César, A.S. Hydrogen: Trends, production and characterization of the main process worldwide. Int. J. Hydrogen Energy 2017, 42, 2018–2033.
- Razu, M.H.; Hossain, F.; Khan, M. Advancement of Bio-hydrogen Production from Microalgae. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Springer: Gateway East, Singapore, 2019; pp. 423–462.
- Grechanik, V.; Romanova, A.; Naydov, I.; Tsygankov, A. Photoautotrophic cultures of Chlamydomonas reinhardtii: Sulfur deficiency, anoxia, and hydrogen production. Photosynth. Res. 2020, 143, 275–286.
- Fakhimi, N.; Tavakoli, O. Improving hydrogen production using co-cultivation of bacteria with Chlamydomonas reinhardtii microalga. Mater. Sci. Energy Technol. 2019, 2, 1–7.
- Skjanes, K.; Knutsen, G.; Kӓllqvist, T.; Lindblad, P. H2 production from marine and freshwater species of green algae during sulfur deprivation and considerations for bioreactor design. Int. J. Hydrog. Energy 2008, 33, 511–521.
- Guan, Y.; Deng, M.; Yu, X.; Zhang, W. Two-stage photo-biological production of hydrogen by marine green alga Platymonas subcordiformis. Biochem. Eng. J. 2004, 19, 69–73.
- Guo, Z.; Chen, Z.; Lu, H.; Fu, Y.; Yu, X.; Zhang, W. Sustained hydrogen photoproduction by marine green algae platymonas subcordiformis integrated with in situ hydrogen consumption by an alkaline fuel cell system. J. Biotechnol. 2008, 136, 558–576.
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62.
- Sialve, B.; Bernet, N.; Bernard, O. Anaerobic digestion of microalgae as a necessary step to make microalgal biodiesel. Sustainable. Biotechnol. Adv. 2009, 27, 409–416.
- Olguin, E.J. The cleaner production strategy applied to animal production. In Environmental Biotechnology and Cleaner Bioprocesses; Olguín, E.J., Sánchez, G., Hemández, E., Eds.; Taylor and Francis: London, UK, 2000; pp. 107–121.
- Yen, H.-W.; Brune, D.E. Anaerobic co-digestion of algal sludge and waste paper to produce methane. Bioresour. Technol. 2007, 98, 130–134.
- Klocke, M.; Mahnert, P.; Mundt, K.; Souidi, K.; Linke, B. Microbial community analysis of a biogas-producing completely stirred tank reactor fed continuously with fodder beet silage as mono-substrate. Syst. Appl. Microbiol. 2007, 30, 139–151.
- Schlüter, A.; Bekel, T.; Diaz, N.N.; Dondrup, M.; Eichenlaub, R.; Gartemann, K.H.; Krahn, I.; Krause, L.; Kromeke, H.; Kruse, O.; et al. The metagenome of a biogas-producing microbial community of a production-scale biogas plant fermenter analysed by the 454-pyrosequencing technology. J. Biotechnol. 2008, 136, 77–90.
- Campbell, J.; Lobell, D.; Field, C. Greater transportation energy and GHG offsets from bioelectricity than ethanol. Science 2009, 324, 1055–1057.
- Börjesson, P.; Berglund, M. Environmental systems analysis of biogas systems -part I: Fuel-cycle emissions. Biomass Bioenergy 2006, 30, 469–485.
- Ubando, A.T.; Rivera, D.R.T.; Chen, W.H.; Culaba, A.B. A comprehensive review of life cycle assessment (LCA) of microalgal and lignocellulosic bioenergy products from thermochemical processes. Bioresour. Technol. 2019, 291, 121837.
- Ubando, A.T.; Felix, C.B.; Chen, W.-H. Biorefineries in circular bioeconomy: A comprehensive review. Bioresour. Technol. 2020, 299, 122585.
- Chandra, R.; Iqbal, H.M.N.; Vishal, G.; Lee, H.S.; Nagra, S. Algal biorefinery: A sustainable approach to valorize algal-based biomass towards multiple product recovery. Bioresour. Technol. 2019, 278, 346–359.
- Kisielewska, M.; Zieliński, M.; Dębowski, M.; Kazimierowicz, J.; Romanowska-Duda, Z.; Dudek, M. Effectiveness of Scenedesmus sp. Biomass Grow and Nutrients Removal from Liquid Phase of Digestates. Energies 2020, 13, 1432.
- Mohan, S.V.; Hemalatha, M.; Chakraborty, D.; Chatterjee, S.; Ranadheer, P.; Kona, R. Algal biorefinery models with self-sustainable closed loop approach: Trends and prospective for blue-bioeconomy. Bioresour. Technol. 2019, 295, 122128.
- Mishra, S.; Roy, M.; Mohanty, K. Microalgal bioenergy production under zero-waste biorefinery approach: Recent advances and future perspectives. Bioresour. Technol. 2019, 292, 122008.
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Comparison of Growth Rate and Nutrient Content of Five Microalgae Species Cultivated in Greenhouses. Plants 2019, 8, 279.
- Aziz, M.M.A.; Kassim, A.K.; Shokravi, Z.; Jakarni, F.M.; Liu, H.Y.; Zaini, N. Two-stage cultivation strategy for simultaneous increases in growth rate and lipid content of microalgae: A review. Renew. Sustain. Energy Rev. 2020, 119, 109621.
- Sánchez-Bayo, A.; Morales, V.; Rodríguez, R.; Vicente, G.; Bautista, L.F. Cultivation of microalgae and cyanobacteria: Effect of operating conditions on growth and biomass composition. Molecules 2020, 25, 2834.
- Neofotis, P.; Huang, A.; Sury, K.; Chang, W.; Joseph, F.; Gabr, A.; Twary, S.; Qiu, W.; Holguine, O.; Polle, J.E.W. Characterization and classification of highly productive microalgae strains discovered for biofuel and bioproduct generation. Algal Res. 2016, 15, 164–178.
- Li, P.; Sakuragi, K.; Makino, H. Extraction Techniques in Sustainable Biofuel Production: A Concise Review. Fuel Process. Technol. 2019, 193, 295–303.
- Piasecka, A.; Nawrocka, A.; Wiącek, D.; Krzemińska, J. Agro-industrial by-product in photoheterotrophic and mixotrophic culture of Tetradesmus obliquus: Production of ω3 and ω6 essential fatty acids with biotechnological importance. Sci Rep. 2020, 10, 1–11.
- Huang, G.H.; Chen, F.; Wei, D.; Zhang, X.W.; Chen, G. Biodiesel production by microalgal biotechnology. Appl. Energy 2010, 87, 38–46.
- Yoo, C.; Jun, S.Y.; Lee, J.Y.; Ahn, C.Y.; Oh, H.M. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour. Technol. 2010, 101, 71–74.
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112.
- Chiu, S.-Y.; Kaom, C.-J.; Chen, C.-H.; Kuan, T.-C.; Ong, S.-C.; Lin, C.-S. Reduction of CO2 by a high-density culture of Chlorella sp. in a semicontinuous photobioreactor. Bioresour. Technol. 2008, 99, 3389–3396.
- Chojnacka, K.; Marquez-Rocha, F.J. Kinetic and stoichiometric relationships of the energy and carbon metabolism in the culture of microalgae. Biotechnology 2004, 3, 21–34.
- Morales-Sánchez, D.; Martinez-Rodriguez, O.A.; Martinez, A. Heterotrophic cultivation of microalgae: Production of metabolites of commercial interest. J. Chem. Technol. Biotechnol. 2016, 92, 925–936.
- De Swaaf, M.; Sijtsma, L.; Pronk, J. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing microalga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672.
- Xu, H.; Miao, X.L.; Wu, Q.Y. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507.
- Kuei-Ling, Y.; Jo-Shu, C. Effects of cultivation conditions and media composition on cell growth and lipid productivity of indigenous microalga Chlorella vulgaris ESP-31. Bioresour. Technol. 2012, 105, 120–127.
- Caporgno, M.P.; Haberkorn, I.; Böcker, L.; Mathys, A. Cultivation of Chlorella protothecoides under different growth modes and its utilisation in oil/water emulsions. Bioresour. Technol. 2019, 288, 121476.
- Sim, S.J.; Joun, J.; Hong, M.E.; Patel, A.K. Split mixotrophy: A novel cultivation strategy to enhance the mixotrophic biomass and lipid yields of Chlorella protothecoides. Bioresour. Technol. 2019, 291, 121820.
- Shen, X.; Qin, Q.; Yan, S.; Huang, J.L.; Liu, K.; Zhou, S.-B. Biodiesel production from Chlorella vulgaris under nitrogen starvation in autotrophic, heterotrophic, and mixotrophic cultures. J. Appl. Phycol. 2019, 31, 1589–1596.
- Li, T.; Kirchhoff, H.; Gargouri, M.; Feng, J.; Cousins, A.B.; Pienkos, P.T.; Gang, D.R.; Chen, S. Assessment of photosynthesis regulation in mixotrophically cultured microalga Chlorella sorokiniana. Algal Res. 2016, 19, 30–38.
- Zheng, Y.; Chi, Z.; Lucker, B.; Chen, S. Two-stage heterotrophic and phototrophic culture strategy for algal biomass and lipid production. Bioresour. Technol. 2012, 103, 484–488.
- Li, T.; Zheng, Y.; Yu, L.; Chen, S. Mixotrophic cultivation of a Chlorella sorokiniana strain for enhanced biomass and lipid production. Biomass Bioenergy 2014, 66, 204–213.
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049.
- Pang, N.; Gu, X.; Chen, S.; Kirchhoff, H.; Lei, H.; Rojec, S. Exploiting mixotrophy for improving productivities of biomass and co-products of microalgae. Renew. Sustain. Energy Rev. 2019, 112, 450–460.
- Chen, Y.H.; Walker, T.H. Biomass and lipid production of heterotrophic microalgae Chlorella protothecoides by using biodiesel-derived crude glycerol. Biotechnol. Lett. 2011, 33, 1973.
- Zuccaro, G.; Yousuf, A.; Pollio, A.; Steyer, J.-P. Microalgae Cultivation Systems. In Microalgae Cultivation for Biofuels Production; Elsevier: London, UK; San Diego, CA, USA; Cambridge, MA, USA; Oxford, UK, 2020; pp. 11–29.
- Cezare-Gomes, E.A.; del Mejia-da-Silva, L.C.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, A.K.; de Carvalho, J.C.M. Potential of Microalgae Carotenoids for Industrial Application. Appl. Biochem. Biotechnol. 2019, 188, 602–634.
- Zhang, Y.; Su, H.; Zhong, Y.; Zhang, C.; Shen, Z.; Sang, W.; Yan, G.; Zhou, X. The effect of bacterial contamination on the heterotrophic cultivation of Chlorella pyrenoidosa in wastewater from the production of soybean products. Water Res. 2012, 46, 5509–5516.
- Marudhupandia, T.; Sathishkumara, R.; Kumara, T.T.A. Heterotrophic cultivation of Nannochloropsis salina for enhancing biomass and lipid production. Biotechnol. Rep. 2016, 10, 8–16.
- Chandra, R.; Arora, S.; Rohit, M.V.; Mohan, S.V. Lipid metabolism in response to individual short chain fatty acids during mixotrophic mode of microalgal cultivation: Influence on biodiesel saturation and protein profile. Bioresour. Technol. 2015, 188, 169–176.
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232.
- Bhatnagar, A.; Chinnasamy, S.; Singh, M.; Das, K.C. Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl. Energy 2011, 88, 3425–3431.
- Yu, H.F.; Jia, S.R.; Dai, Y.J. Growth characteristics of the cyanobacterium Nostoc flagelliformein photoautotrophic, mixotrophic and heterotrophic cultivation. J. Appl. Phycol. 2009, 21, 127–133.
- Bailey, R.; DiMasi, D.; Hansen, J.; Mirrasoul, P.; Ruecker, C.; Kaneko, T.; Barclay, W. Enhanced Production of Lipids Containing Polyenoic Fatty Acid by Very High Density Cultures of Eukaryotic Microbes in. Fermentors Patent No.: US 6,607,900 B2, 19 August 2003.
- Anand, P.; Saxena, R.K. A comparative study of solvent-assisted pretreatment of biodiesel derived crude glycerol on growth and 1,3-propanediol production from Citrobacter freundii. New Biotechnol. 2011, 29, 199–205.
- Jiang, Y.; Yoshida, T.; Quigg, A. Photosynthetic performance, lipid production and biomass composition in response to nitrogen limitation in marine microalgae. Plant Physiol. Biochem. 2012, 54, 70–77.
- Ogbonna, J.C.; Ichige, E.; Tanaka, H. Regulating the ratio of photoautotrophic to heterotrophic metabolic activities in photoheterotrophic culture of Euglena gracilis and its application to alpha-tocopherol production. Biotechnol. Lett. 2002, 24, 953–958.
- Reverdatto, S.; Beilinson, V.; Nielsen, N.C. A multisubunit acetyl coenzyme A carboxylase fromsoybean. Plant Physiol. 1999, 119, 961–978.
- Chen, J.W.; Liu, W.J.; Hu, D.X.; Wang, X.; Balamurugan, S.; Alimujiang, A.; Yang, W.D.; Liu, J.S.; Li, H.Y. Identification of a malonyl CoA-acyl carrier protein transacylase and its regulatory role infatty acid biosynthesis in oleaginous microalga Nannochloropsis oceanic. Biotechnol. Appl. Biochem. 2016, 64, 620–626.
- Fan, J.; Andre, C.; Xu, C. A chloroplast pathway for the de novo biosynthesis of triacylglycerol inChlamydomonas reinhardtii. FEBS Lett. 2011, 585, 1985–1991.
- Wan, L.; Han, J.; Sang, M.; Li, A.; Wu, H.; Yin, S.; Zhang, C. De novo transcriptomic analysis of anoleaginous microalga: Pathway description and gene discovery for production of next-generationbiofuels. PLoS ONE 2012, 7, e35142.
- Lenka, S.K.; Carbonaro, N.; Park, R.; Miller, S.M.; Thorpe, I.; Li, Y. Current advances in molecular, biochemical, and computational modeling analysis of microalgal triacylglycerol biosynthesis. Biotechnol. Adv. 2016, 34, 1046–1063.
- Show, K.-Y.; Yan, Y.-G.; Lee, D.-J. Biohydrogen production from algae: Perspectives, challenges, and prospects. In Biofuels from Algae, 2nd ed.; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2019; pp. 325–343.
- Dasgupta, C.N.; Gilbert, J.J.; Lindblad, P.; Heidorn, T.; Borgvang, S.A.; Skjanes, K.; Das, D. Recent trends on the development of photobiological processes and photobioreactors for the improvement of hydrogen production. Int. J. Hydrog. Energy 2010, 35, 10218–10238.
- Miyake, J.; Miyake, M.; Asada, Y. Biotechnological hydrogen production: Research for efficient light energy conversion. J. Biotechnol. 1999, 70, 89–101.
- Ni, F.M.; Leung, D.Y.C.; Leung, M.K.H.; Sumathy, K. An overview of hydrogen production from biomass. Fuel Process. Technol. 2006, 87, 461–472.
- Akhlaghi, N.; Najafpour-Darzi, G. A comprehensive review on biological hydrogen production. Int. J. Hydrog. Energy 2020, 45, 22492–22512.
- Das, D.; Veziroglu, T.N. Hydrogen production by biological processes: A survey of literature. Int. J. Hydrog. Energy 2001, 26, 13–28.
- Kim, D.H.; Kim, M.S. Hydrogenases for biological hydrogen production. Bioresour. Technol. 2011, 102, 8423–8431.