2. Mechanism of Angiogenesis
The angiogenesis process embodies forming new blood vessels from existing vessels in response to physiological and pathological mechanisms [
41]. During embryogenesis, the vascular network develops through a combination of vasculogenesis, referred to as the de novo formation of the heart and new blood vessels from stem endothelial cells, namely, angioblasts, and angiogenesis, which expands the initial primitive vascular plexus [
42]. Although most blood vessels remain quiescent under physiological conditions, tissue repair and regeneration through wound healing, ovulation, and endometrial thickening throughout the menstrual cycle are based on angiogenesis for proper functioning [
43,
44,
45]. While vascular growth varies depending on where angiogenesis is initiated and the tissue to which they will provide a new blood supply, several mechanisms are common in forming these vessels [
46]. In a hypoxic state, the recruitment of cells that promote inflammation; angiogenic growth factor production; degradation of the basement membrane; and endothelial cells (ECs) sprouting, migrating, proliferating, differentiating, and modulating vascular support cells are some of the shared characteristics in angiogenesis [
47].
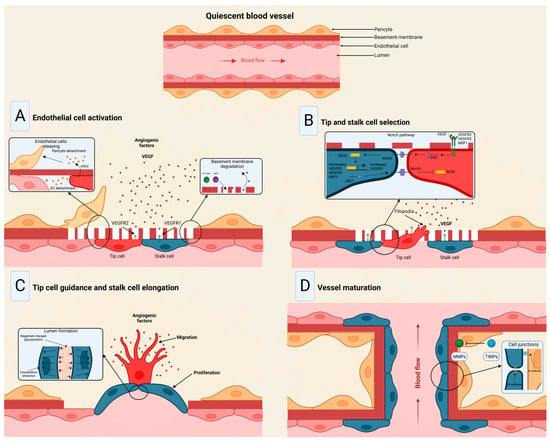
The angiogenic process (
Figure 3) comprises several stages involving the sprouting, migration, and proliferation of ECs guided by the vascular endothelial growth factor (VEGF) [
48]. Following VEGF stimulation, pericytes from the vessel wall detach, and the basal membrane is weakened by proteolytic degradation. At the same time, ECs adopt an invasive and motile phenotype called tip cells that send out filamentous pseudopodia to guide vascular budding [
49]. The cells behind the tip cells are denominated stalk cells, which proliferate to maintain the integrity of the structure and function of the nascent vessels, mainly expanding the vascular lumen [
50]. ECs modify their shape by negatively charging glycoproteins on the apical surface to repel each other and open the lumen while redistributing cell-to-cell adhesion to the periphery [
51]. For maturation to occur, pericytes must be recruited by the platelet-derived growth factor subunit B (PDGF-B) and angiopoietin 1 (Ang1) signaling along with the strengthening and consolidation of the adhesion between ECs with junctional molecules such as VE-cadherin, while a basement membrane is deposited by tissue inhibitors of metalloproteinases (TIMPs) [
52,
53].
Figure 3. Stages of the angiogenic process. (
A) Angiogenic signals, such as VEGF, promote pericyte detachment from the basement membrane and weaken the extracellular matrix. (
B) Endothelial cells display characteristic phenotypes after VEGF stimulation: migratory tip cells or proliferating stalk cells. (
C) Attractive and repulsive forces control endothelial cells, forming a vessel lumen to initiate blood flow. (
D) PDGF-B and Ang1 signaling lead to pericyte recruitment, while junctional molecules consolidate EC–EC adhesion. Modified from Viallard et al. [
47].
Under physiological conditions, angiogenesis is strongly regulated by factors ranging from metabolites to hormones [
54]. Various molecular pathways have been extensively studied that comprise the angiogenesis process, including the VEGF-VEGFR, Angiopoietin-Tie, Delta-Notch, and Ephrin-Eph [
55]. Angiogenesis, vasculogenesis, and lymphangiogenesis are mostly regulated by six VEGF members encoded in the human genome, namely, VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, and the placenta growth factor (P1GF) [
48,
56]. According to their molecular configuration and affinity, the aforementioned factors may bind to different tyrosine kinase receptors VEGFR-1, VEGFR-2, and/or VEGFR-3; VEGF-A binds to VEGFR-2 to contribute to angiogenesis, whereas VEGF-C and VEGF-D enhance lymphatic vessel sprouting by binding to VEGFR-3 [
57]. Another group of angiogenesis inducers are the platelet-derived growth factors (PDGF), which induce ECs proliferation and migration by binding to two tyrosine kinase receptors, PDGFR-α and PDGFR-β [
58]. Another angiogenesis factor is the fibroblast growth factor (FGF) 2, responsible for inducing metalloproteinase (MMP) secretion to degrade the basement membrane and promote vessel sprouting along with VEGF [
59]. Angiopoietin 1, interleukin 8 (IL-8), epidermal growth factor (EGF), and tumor necrosis factor α (TNF-α) also exert a pro-angiogenic effect in ECs through several signaling pathways [
60,
61,
62,
63]. Conversely, angiopoietin 2, angiostatin, endostatin, vasostatin, and TIMPs inhibit angiogenesis and play an important role in achieving vascular homeostasis [
64,
65].
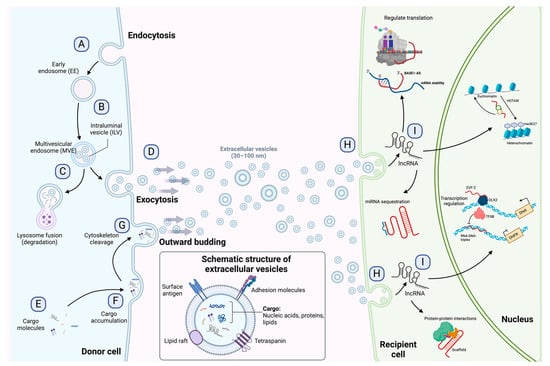
Extracellular vesicles produced by many cellular lineages under specific circumstances can be taken up by ECs to promote and regulate angiogenesis [
66]. For instance, a study in endometrial stromal cells (HESCs) found active secretion of EVs during decidualization in a controlled manner by the hypoxia-inducible factor 2 alpha (HIF2α)–Ras-related protein Rab-27B (RAB27B) cascade, revealing a cargo with a variety of growth regulators, signaling molecules, metabolic modulators, and factors that control the expansion and remodeling of ECs [
67]. In a myocardial infarction animal model, stem cell-derived small extracellular vesicles (sEVs) loaded with miR-486-5p promoted cardiac angiogenesis via fibroblastic MMP19-VEGFA cleavage signaling [
68]. Moreover, Gregorius et al. [
69] evaluated the effects of mesenchymal stromal cell (MSC)-derived sEVs on the proliferation, migration, and tube formation of cerebral microvascular ECs. Interestingly, hypoxic preconditioning enhanced angiogenesis and increased post-ischemic endothelial survival by regulating several miRNAs through the uptake of protein-enriched sEVs cargo. Another study demonstrated that HS-1 protein X-1 (HAX1), a major regulator of myeloid homeostasis, was present in EVs secreted by nasopharyngeal carcinoma (NPC) tumors, promoting an angiogenesis phenotype by activating the focal adhesion kinase (FAK) pathway in ECs by increasing the expression level of integrin subunit beta 6 (ITGB6) [
70]. Conversely, EVs produced by bone marrow MSCs were found to be enriched in the cluster of differentiation 39 (CD39), TIMP-1, and CD73, inhibiting tumor angiogenesis by targeting the extracellular matrix remodeling and the endothelial cell migration [
71].
3. Exosomal Long Non-Coding and Circular RNAs in Cancer Angiogenesis
LncRNAs in EV cargo have been demonstrated lately, mainly in cancer [
72,
73,
74]. A recent study launched an online repository of EV long RNAs (exLRs) in diverse human body fluids, comprising 19,643 mRNAs, 15,645 lncRNAs, and 79,084 circRNAs obtained from human blood, cerebrospinal fluid, bile, and urine samples. The database provides novel exLR signatures to help discover new biomarkers that could aid in diagnosing and treating many diseases [
75]. Based on available recent research, Casado-Díaz et al. [
76] concluded that lncRNAs and other RNAs included in MSC-derived EVs can be applied in chronic skin ulcers to improve accelerated healing and decrease scar formation due to immunosuppressive and immunomodulatory properties. Conversely, in a diabetic wound-healing animal model, upregulated lncRNAs packed in EVs from fibroblasts enhanced keratinocyte MMP-9 expression to induce collagen degradation, delaying wound healing [
77]. Recently, the long non-coding repressor of NFAT (NRON) was detected in BMSC-derived EVs, inhibiting osteoclast differentiation and osteoporotic bone loss in vitro and in vivo [
78].
In tumors, the high rate of cell proliferation forces the formation of new blood vessels [
79]. However, in most cases, these blood vessels are dilated, tortuous, and immature, leading to excessive permeability and increased hypoxia [
80]. In addition, vascular disorganization causes heterogeneity in the tumor blood vessel network, creating highly vascularized tumor areas and other hypoxic areas with low vascular density [
47]. Thus, hypoxia becomes a major driver of tumor angiogenesis, along with other mechanisms promoted by activated oncogenes or loss of tumor suppressor genes, in which lncRNAs play an important role, mainly through acting as competing endogenous RNAs for miRNAs [
81]. Similarly, circRNAs have been extensively studied in cancer, elucidating important roles in tumor development, growth, and angiogenesis [
82]. For instance, VEGFR-related pathways have been linked to circRNAs by affecting tumor angiogenesis by sponging miRNAs [
83,
84]. The landscape of exosomal lnc- and circRNAs in angiogenesis in cancer is summarized in
Table 1 and
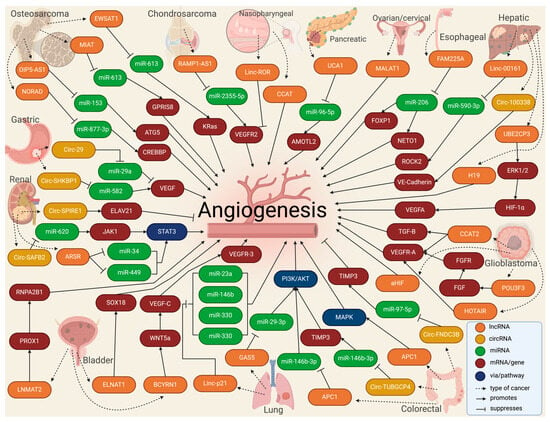
Figure 4.
Figure 4. Molecular landscape of exosomal lnc- and circRNAs in angiogenesis in cancer.
Table 1. The landscape of exosomal lnc- and circRNAs in angiogenesis in cancer.
3.1. Bone Malignancies
Some studies have been dedicated to studying exosomal lncRNAs in bone malignancies. LncRNA Opa-interacting protein 5-antisense 1 (OIP5-AS1) was found to be overexpressed in exosomes secreted by osteosarcoma cells, increasing angiogenesis in tubule formation assays by mechanistically sponging miR-153 and increasing the autophagy-related 5 protein (ATG5) [
85]. Interestingly, serum samples from osteosarcoma patients could transfer via EVs the myocardial infarction-associated transcript (MIAT), promoting the proliferation of osteosarcoma cell lines and angiogenesis in HUVECs by sponging miR-613 and upregulating G protein-coupled receptor 158 (GPR158) [
86]. In an in vitro and animal model, BMSC-EVs carried the non-coding RNA activated by DNA damage (NORAD) into osteosarcoma cells and upregulated CREB-binding protein (CREBBP) by sponging miR-877-3p to promote proliferation, invasion, migration, and angiogenesis [
87]. Another lncRNA called Ewing sarcoma-associated transcript 1 (EWSAT1) was found to regulate osteosarcoma-induced angiogenesis via two mechanisms: (1) by increasing in sensitivity/reactivity of vascular endothelial cells triggered by exosomes carrying EWSAT1, and (2) by increasing angiogenic factors secretion [
88]. Moreover, exosomes secreted by chondrosarcoma cells were loaded with the receptor activity-modifying protein 2 antisense 1 (RAMP2-AS1). They could enhance HUVECs proliferation, migration, and tube formation by acting as a ceRNA for miR2355-5p to regulate VEGFR2 expression. In addition, the overexpression of RAMP2-AS1 in the serum of chondrosarcoma patients was demonstrated to be closely related to local invasiveness, distant metastasis, and poor prognosis [
89].
3.2. Esophageal, Gastric, and Colorectal Cancers
Some of the most prevalent tumors of the gastrointestinal (GI) tract have been explored regarding the role played in angiogenesis by exosomes loaded with different lncRNAs and circRNAs. For instance, the exosomal lncRNA family with sequence similarity 225 member A (FAM225A) was highly expressed in esophageal squamous cell carcinoma (ESCC), upregulating neuropilin and tolloid-like 2 (NETO2) and forkhead box P1 (FOXP1) expression by sponging miR-206 to accelerate tumor progression and angiogenesis [
93].
In gastric cancer patients, exosomal circ-SHKBP1 was overexpressed in tumor and blood samples. When the exosomes were isolated and exposed to different cell lines, cells showed a promoted proliferation, invasion, migration, and angiogenesis rate by mechanistically regulating the miR-582-3p/HUR/VEGF axis and suppressing heat shock protein 90 (HSP90) degradation [
116]. Similarly, 30 blood samples and tissues from gastric cancer patients were taken to analyze circ-FCH and mu domain-containing endocytic adaptor 2 (FCHO2). It was found that circ-FCHO2 up-modulation led to a poor outcome, while circ-FCHO2 silencing weakened the proliferation, invasion, angiogenesis, and stem cell characteristics, presumably by activating the Janus kinase 1 (JAK1)/signal transducer and activator of transcription 2 (STAT2) pathway via sponging miR-194-5p [
117]. Conversely, by acting as a miR-587 sponge to adjust the expression of the sclerostin domain-containing 1 (SOSTDC1), circ-0001190 overexpression inhibited cell viability, proliferation, angiogenesis, migration, and invasion of gastric cancer cell lines [
114]. Moreover, circ-0044366 was highly expressed in gastric cancer and impaired the proliferation, migration, and tube formation of HUVECs by exosomal communication by acting as miR-29a ceRNA and regulating the VEGF pathway [
115].
In colorectal cancer (CRC), tumor growth, angiogenesis, and liver metastasis were suppressed by exosomal circ-fibronectin type III domain-containing 3B (FNDC3B) overexpression by acting via the miR-97-5p/TIMP3 pathway [
98]. Similarly, exosomes derived from lncRNA adenomatous polyposis coli (APC1)-silenced CRC cells promoted angiogenesis by activating the mitogen-activated protein kinase 1 (MAPK) pathway in endothelial cells, while enforced APC1 was sufficient to inhibit CRC growth, metastasis, and tumor angiogenesis by suppressing exosome production [
99]. Interestingly, exosomes loaded with circ-tubulin gamma complex component 4 (TUBGCP4) derived from CRC cells enhanced vascular endothelial cell migration and tube formation via inducing filopodia formation and endothelial cell tipping by upregulating the pyruvate dehydrogenase kinase 2 (PDK2) to activate the AKT serine/threonine kinase 1 (AKT) signaling pathway and by sponging miR-146b-3p [
100]. A very interesting study by Zhi et al. [
125] compared EVs derived from the b-Raf proto-oncogene (BRAF) wild-type CRC and the BRAF
V600E mutant patients to find the overexpression of 13 lncRNAs and downregulation of 22 lncRNAs in exosomes from the BRAF
V600E mutation type. This difference showed a higher microvascular and micro-lymphatic vessel density of the BRAF
V600E mutant CRC tissues.
3.3. Liver and Pancreatic Cancers
LncRNA-loaded exosomes from tumors from other organs related to the GI tract have also shown some relationship with tumor angiogenesis. You et al. [
101] reported high levels of Linc-00161 in serum-derived exosomes from hepatocellular cancer (HCC) patients and the supernatants of HCC cell lines, which are associated with poor survival. Mechanistically, Linc-00161 promoted angiogenesis in HUVECs by inhibiting miR-590-3p and activating the Rho-associated coiled-coil-containing protein kinase 2 (ROCK2) axis. In an in vitro study, exosomes with lncRNA H19 were released by CD90+ HCC cells and modulated endothelial cells, promoting an angiogenic phenotype and cell-to-cell adhesion [
103]. Similarly, lncRNA ubiquitin-conjugating enzyme E2 C pseudogene 3 (UBE2CP3) was overexpressed in HCC EVs. It promoted HUVEC proliferation, migration, and tube formation via the activation of the ERK/HIF-1α/p70S6K/VEGFA signaling cascade, promoting HCC tumorigenicity [
102]. In another study, exosomal circ-100388 affected the cell proliferation, angiogenesis, permeability, and vasculogenic mimicry formation ability of HUVECs and HCC tumor metastasis [
104].
In cholangiocarcinoma (CCA), the cholangiocarcinoma-associated circular RNA 1 (circ-CCAC1) from CCA-derived EVs was transferred to endothelial monolayer cells, disrupting endothelial barrier integrity and inducing angiogenesis. Interestingly, circ-CCAC1 increased cell leakiness by sequestering the enhancer of zeste 2 polycomb repressive complex 2 subunit (EZH2) gene, thus elevating the SH3 domain-containing GRB2 like 2, endophilin A1 (SH3GL2) expression to reduce levels of intercellular junction proteins [
124].
In pancreatic cancer, the expression levels of the lncRNA urothelial cancer-associated 1 (UCA1) in exosomes derived from the serum of patients were associated with poor survival, promoting angiogenesis and tumor growth by acting as a ceRNA of miR-96-5p, relieving the repressive effects on the expression of its target gene angiomotin like 2 (AMOTL2) [
90]. Moreover, the exosomal small nucleolar RNA host gene 11 (SNHG11) promoted cell proliferation, migration, and angiogenesis in pancreatic cancer cell lines but impeded cell apoptosis via sponging miR-324-3p to upregulate VEGFA expression [
126].
3.4. Renal and Bladder Cancers
Some urinary system tumors have observed a relationship between lncRNA-loaded EVs and angiogenesis. In renal cell carcinoma (RCC), RCC-derived exosomes had an lncRNA Ars operon (ARSR) that promoted macrophage polarization, cytokine release, phagocytosis, angiogenesis, and tumor development by sponging miR34/miR-449 and upregulating the signal transducer and activator of transcription 3 (STAT3) pathway [
108]. Similarly, RCC-derived exosomal circular scaffold attachment factor B2 (circ-SAFB2) facilitated the progression, invasion, angiogenesis, and metastasis of RCC by inducing the polarization of M2 macrophages through the miR-620/JAK1/STAT3 axis [
109]. Conversely, exosomal circular spire type actin nucleation factor 1 (circ-SPIRE1) suppressed angiogenesis and vessel permeability through regulating ELAV-like RNA-binding protein 1-mRNA, binding and upregulating polypeptide N-Acetylgalactosaminyltransferase 3 (GALNT3) and KH domain RNA-binding protein (QKI) expression [
110].
In bladder carcinoma (BCa), exosomal brain cytoplasmic RNA 1 (BCYRN1) promoted the tube formation and migration of human lymphatic endothelial cells (HLECs), upregulating the Wnt family member 5A (WNT5A) gene expression by inducing hnRNPA1-associated H2K4 trimethylation in WNAT5a promoter, which activated Wnt/β-catenin signaling to facilitate the secretion of VEGF-C in BCa [
111]. Moreover, lymph node metastasis-associated transcript 2 (LNMAT2)-loaded exosomes from BCa tissues and blood samples stimulated the tube formation and migration of HLECs and enhanced tumor lymphangiogenesis and lymph node metastasis by upregulation of prospero homeobox 1 (PROX1) gene expression by recruitment of hnRNPA2B1 and increasing H3K4 trimethylation [
112]. Comparably, BCa cell-secreted EVs mediated intercellular communication with HLECs through the transmission of the small nucleolar RNA host gene 16 (ELNAT1) and promoted lymphangiogenesis by inducing the ubiquitin-conjugating enzyme E2 (UBC9) gene overexpression to catalyze the small ubiquitin-like modifier (SUMO) binding of hnRNPA1 at the lysine 113 residue [
113].
3.5. Nasopharyngeal and Lung Cancers
LncRNAs in exosomes derived from nasopharyngeal squamous cell carcinoma (NPSCC) and their relationship with tumor angiogenesis have been mildly explored. In serum samples from newly diagnosed NPSCC patients, the long intergenic non-protein-coding RNA, regulator of reprogramming (linc-ROR), was substantially expressed in exosomes that could be taken up by HUVECs, increasing proliferation, migration, and angiogenesis in vitro by mechanistically upregulating the p-AKT/p-VEGFR2 pathway [
91]. Similarly, lncRNA colon cancer-associated transcript 2 (CCAT2) was found in EVs derived from NPSCC patients, promoting HUVEC proliferation and angiogenesis promotion [
92].
In non-small cell lung carcinoma (NSCLC), NSCLC cells secreted exosomes with melanotransferrin antisense 1 (MFI2-AS1) to induce tube formation by HUVECs, promoting angiogenesis and metastasis by sponging miR-107, which in turn activated the PI3K/AKT pathway [
105]. Similarly, high EV Linc-p21 was found in NSCLC blood samples from tumor-draining pulmonary veins before tumor surgical resection. EVs with Linc-p21 were taken up by HUVECs and promoted tube formation and enhanced tumor cell adhesion to endothelial cells by sponging miR-23a, miR-146bv, miR-330, and miR-494 [
106]. In contrast, GAS5 was lowly expressed in human lung cancer tissues, lung cancer cells, and cell culture supernatant exosomes. The exosomes of lung cancer cells containing high GAS5 levels inhibited HUVECs proliferation and tube formation, increasing their apoptosis by sponging miR-29-3p and upregulating phosphatase and tensin homolog (PTEN) and inhibiting PI3K/AKT phosphorylation [
107].
3.6. Glioma and Gliobastoma
A few studies have reported evidence of the role of exosomal lncRNAs in glioma and glioblastoma angiogenesis. An in vitro study with glioma cell lines demonstrated that HUVECs can take up exosomal CCAT2 to promote migration, proliferation, tubular-like structure formation, and arteriole formation [
94]. Similarly, the POU class 3 homeobox 3 (POU3F3) was upregulated in glioma tissue. When human brain microvascular endothelial cells (HBMVECs) were treated with exosomes loaded with POU3F3, they exhibited better migration, proliferation, tubular-like structure formation, and arteriole formation. Mechanistically, POU3F3 was shown to upregulate bFGF, bFGFR, VEGFA, and Angio [
95]. Moreover, cell line A172 was cultured to demonstrate that EVs loaded with the HOX transcript antisense RNA (HOTAIR) had a pro-angiogenic activity in HBMVECS via VEGFA [
96]. In glioblastoma, lncRNA HIF1A antisense RNA 2 (AHIF) was found upregulated in tissue samples, and when cultured with glioblastoma cell lines, exosomal AHIF regulated factors associated with migration and angiogenesis [
97].
3.7. Other Cancer Types
In ovarian cancer, an in vitro study revealed that lncRNA activated by TGF-β (ATB) promoted viability and angiogenesis of HUVECs by sponging miR-204-3p and thus upregulating TGFβ-R2 [
127]. Similarly, elevated serum exosomal metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) promoted angiogenesis and was highly correlated with an advanced and metastatic phenotype of epithelial ovarian cancer [
118]. Another study demonstrated taurine-upregulated 1 (TUG1) overexpression in human cervical cancer cell lines. When TUG1 was depleted, the exosome-mediated pro-angiogenic potential of HUVECs was impaired by modulating angiogenesis-related genes like VEGFA, MMP9, TGFβ, IL-8, and bFGF [
119]. In breast cancer cell lines, the metadherin (MTDH) gene improved cell viability and angiogenesis in endothelial cells. The molecular cascade was promoted by exosomal circular homeodomain-interacting protein kinase 3 (circ-HIPK3), which sponged miR-124-3p and in turn upregulated MTDH [
120]. Liu et al. Campo [
121] demonstrated that exosomal overexpression of the FYVE, RhoGF, and PH domain-containing 5 antisense 1 (FGD5-AS1) enhanced the proliferation, migration, angiogenesis, and permeability of HUVECs by regulating the endothelial miR-6838-5p/Vav guanine nucleotide exchange factor 2 (VAV2) axis. A total of 25 peripheral blood samples from 20 multiple myeloma patients and 5 matched healthy controls showed overexpression of the circular ATPase phospholipid-transporting 10A (circ-ATP10A) in the multiple myeloma samples, mechanistically acting as a sponge of several miRNAs to consequently regulate the expression of downstream VEGFB, HIF1A, PDGFA, and FGF [
122].