Reactive oxygen species (ROS) were formerly known as mere byproducts of metabolism with damaging effects on cellular structures. The discovery and description of NADPH oxidases (Nox) as a whole enzyme family that only produce this harmful group of molecules was surprising. Among the Nox isoforms, the NADPH oxidase 3 is the perhaps most underrated Nox enzyme, since it was firstly discovered in the inner ear. Despite the fact that Nox3 is expressed not only in the inner ear but also in various cell types and organs, the “inner ear stigma” remains until today. However ,the involvment of Nox3 is not just limited to the inner ear but extends to various organs and the related diseases.
- NADPH oxidase
- Nox3
- reactive oxygen species
- cardivascular diseases
- hearing loss
- lung diseases
1. Role of Nox3 in Hearing Loss
1.1. Noise-Induced Ototoxicity
1.2. Cisplatin-Induced Ototoxicity
1.3. Cytomegalovirus-Induced Hearing Loss
1.4. Age-Induced Hearing Loss
2. Role of Nox3 during Vertigo
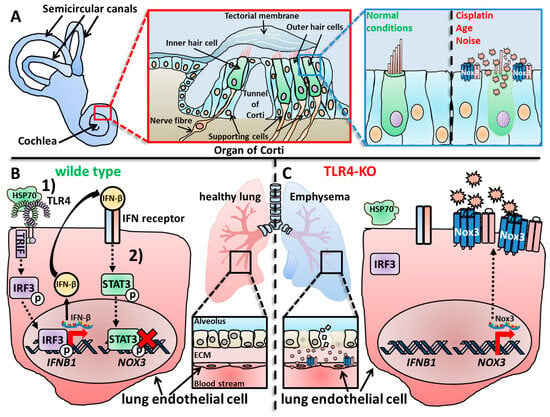
3. Role of Nox3 during Lung Diseases
4. Role of Nox3 during Cardiovasclar Diseases
4.1. Nox3 and Type 2 Diabetes
4.2. Nox3 and Adipositas
4.3. Nox3 and Stroke
4.4. Nox3 and Heart Failure
5. Role of Nox3 during Renal Diseases
6. Role of Nox3 during Gastrointetinal Disaeses
7. Role of Nox3 in Other Diseases
This entry is adapted from the peer-reviewed paper 10.3390/antiox13020219
References
- Thorne, P.R.; Nuttall, A.L.; Scheibe, F.; Miller, J.M. Sound-induced artifact in cochlear blood flow measurements using the laser doppler flowmeter. Hear. Res. 1987, 31, 229–234.
- Muller, U.; Barr-Gillespie, P.G. New treatment options for hearing loss. Nat. Rev. Drug Discov. 2015, 14, 346–365.
- Petit, C.; Bonnet, C.; Safieddine, S. Deafness: From genetic architecture to gene therapy. Nat. Rev. Genet. 2023, 24, 665–686.
- Kamogashira, T.; Fujimoto, C.; Yamasoba, T. Reactive oxygen species, apoptosis, and mitochondrial dysfunction in hearing loss. BioMed Res. Int. 2015, 2015, 617207.
- Pittman, C.A.; Ward, B.K.; Nieman, C.L. A review of adult-onset hearing loss: A primer for neurologists. Curr. Treat Options Neurol. 2021, 23, 20.
- Le, T.N.; Straatman, L.V.; Lea, J.; Westerberg, B. Current insights in noise-induced hearing loss: A literature review of the underlying mechanism, pathophysiology, asymmetry, and management options. J. Otolaryngol. Head Neck Surg. Le J. D’oto-Rhino-Laryngol. Chir. Cervico-Faciale 2017, 46, 41.
- Wang, J.; Puel, J.L. Presbycusis: An update on cochlear mechanisms and therapies. J. Clin. Med. 2020, 9, 218.
- Rousset, F.; Nacher-Soler, G.; Coelho, M.; Ilmjarv, S.; Kokje, V.B.C.; Marteyn, A.; Cambet, Y.; Perny, M.; Roccio, M.; Jaquet, V.; et al. Redox activation of excitatory pathways in auditory neurons as mechanism of age-related hearing loss. Redox Biol. 2020, 30, 101434.
- Liberman, M.C.; Kujawa, S.G. Cochlear synaptopathy in acquired sensorineural hearing loss: Manifestations and mechanisms. Hear. Res. 2017, 349, 138–147.
- Wu, P.Z.; Liberman, L.D.; Bennett, K.; de Gruttola, V.; O’Malley, J.T.; Liberman, M.C. Primary neural degeneration in the human cochlea: Evidence for hidden hearing loss in the aging ear. Neuroscience 2019, 407, 8–20.
- Peineau, T.; Belleudy, S.; Pietropaolo, S.; Bouleau, Y.; Dulon, D. Synaptic release potentiation at aging auditory ribbon synapses. Front. Aging Neurosci. 2021, 13, 756449.
- Frolenkov, G.I.; Belyantseva, I.A.; Friedman, T.B.; Griffith, A.J. Genetic insights into the morphogenesis of inner ear hair cells. Nat. Rev. Genet. 2004, 5, 489–498.
- Schwander, M.; Kachar, B.; Muller, U. Review series: The cell biology of hearing. J. Cell Biol. 2010, 190, 9–20.
- Atkinson, P.J.; Huarcaya Najarro, E.; Sayyid, Z.N.; Cheng, A.G. Sensory hair cell development and regeneration: Similarities and differences. Development 2015, 142, 1561–1571.
- Furness, D.N. Molecular basis of hair cell loss. Cell Tissue Res. 2015, 361, 387–399.
- Kujawa, S.G.; Liberman, M.C. Translating animal models to human therapeutics in noise-induced and age-related hearing loss. Hear. Res. 2019, 377, 44–52.
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear Hear. 2006, 27, 1–19.
- Ramkumar, V.; Mukherjea, D.; Dhukhwa, A.; Rybak, L.P. Oxidative stress and inflammation caused by cisplatin ototoxicity. Antioxidants 2021, 10, 1919.
- Rousset, F.; Carnesecchi, S.; Senn, P.; Krause, K.H. Nox3-targeted therapies for inner ear pathologies. Curr. Pharm. Des. 2015, 21, 5977–5987.
- Clerici, W.J.; Yang, L. Direct effects of intraperilymphatic reactive oxygen species generation on cochlear function. Hear. Res. 1996, 101, 14–22.
- Bielefeld, E.C.; Hu, B.H.; Harris, K.C.; Henderson, D. Damage and threshold shift resulting from cochlear exposure to paraquat-generated superoxide. Hear. Res. 2005, 207, 35–42.
- Wong, A.C.; Ryan, A.F. Mechanisms of sensorineural cell damage, death and survival in the cochlea. Front. Aging Neurosci. 2015, 7, 58.
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Paludetti, G.; Troiani, D. Targeting dysregulation of redox homeostasis in noise-induced hearing loss: Oxidative stress and ros signaling. Free Radic. Biol. Med. 2019, 135, 46–59.
- Fujioka, M.; Kanzaki, S.; Okano, H.J.; Masuda, M.; Ogawa, K.; Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J. Neurosci. Res. 2006, 83, 575–583.
- Kaygusuz, I.; Ozturk, A.; Ustundag, B.; Yalcin, S. Role of free oxygen radicals in noise-related hearing impairment. Hear. Res. 2001, 162, 43–47.
- Yamane, H.; Nakai, Y.; Takayama, M.; Iguchi, H.; Nakagawa, T.; Kojima, A. Appearance of free radicals in the guinea pig inner ear after noise-induced acoustic trauma. Eur. Arch. Oto-Rhino-Laryngol. 1995, 252, 504–508.
- Ohlemiller, K.K.; Wright, J.S.; Dugan, L.L. Early elevation of cochlear reactive oxygen species following noise exposure. Audiol. Neuro-Otol. 1999, 4, 229–236.
- Clerici, W.J.; Hensley, K.; DiMartino, D.L.; Butterfield, D.A. Direct detection of ototoxicant-induced reactive oxygen species generation in cochlear explants. Hear. Res. 1996, 98, 116–124.
- Priuska, E.M.; Schacht, J. Formation of free radicals by gentamicin and iron and evidence for an iron/gentamicin complex. Biochem. Pharmacol. 1995, 50, 1749–1752.
- Hirose, K.; Hockenbery, D.M.; Rubel, E.W. Reactive oxygen species in chick hair cells after gentamicin exposure in vitro. Hear. Res. 1997, 104, 1–14.
- Dehne, N.; Lautermann, J.; Petrat, F.; Rauen, U.; de Groot, H. Cisplatin ototoxicity: Involvement of iron and enhanced formation of superoxide anion radicals. Toxicol. Appl. Pharmacol. 2001, 174, 27–34.
- Paparella, M.M.; Oda, M.; Hiraide, F.; Brady, D. Pathology of sensorineural hearing loss in otitis media. Ann. Otol. Rhinol. Laryngol. 1972, 81, 632–647.
- Merchant, S.N.; Gopen, Q. A human temporal bone study of acute bacterial meningogenic labyrinthitis. Am. J. Otol. 1996, 17, 375–385.
- Fujimoto, C.; Yamasoba, T. Oxidative stresses and mitochondrial dysfunction in age-related hearing loss. Oxidative Med. Cell. Longev. 2014, 2014, 582849.
- Warraich, U.E.; Hussain, F.; Kayani, H.U.R. Aging—Oxidative stress, antioxidants and computational modeling. Heliyon 2020, 6, e04107.
- Kozakiewicz, M.; Kornatowski, M.; Krzywinska, O.; Kedziora-Kornatowska, K. Changes in the blood antioxidant defense of advanced age people. Clin. Interv. Aging 2019, 14, 763–771.
- Yamashita, D.; Jiang, H.Y.; Schacht, J.; Miller, J.M. Delayed production of free radicals following noise exposure. Brain Res. 2004, 1019, 201–209.
- Ohlemiller, K.K.; McFadden, S.L.; Ding, D.L.; Lear, P.M.; Ho, Y.S. Targeted mutation of the gene for cellular glutathione peroxidase (gpx1) increases noise-induced hearing loss in mice. J. Assoc. Res. Otolaryngol. JARO 2000, 1, 243–254.
- Yamane, H.; Nakai, Y.; Takayama, M.; Konishi, K.; Iguchi, H.; Nakagawa, T.; Shibata, S.; Kato, A.; Sunami, K.; Kawakatsu, C. The emergence of free radicals after acoustic trauma and strial blood flow. Acta Oto-Laryngol. Suppl. 1995, 519, 87–92.
- Henderson, D.; McFadden, S.L.; Liu, C.C.; Hight, N.; Zheng, X.Y. The role of antioxidants in protection from impulse noise. Ann. N. Y. Acad. Sci. 1999, 884, 368–380.
- Hu, B.H.; Henderson, D.; Nicotera, T.M. Involvement of apoptosis in progression of cochlear lesion following exposure to intense noise. Hear. Res. 2002, 166, 62–71.
- Bohne, B. Mechanisms of Noise Damage in the Inner Ear; John H. Mills Raven Press: New York, NY, USA, 1976; pp. 41–68.
- Bohne, B.A.; Harding, G.W.; Lee, S.C. Death pathways in noise-damaged outer hair cells. Hear. Res. 2007, 223, 61–70.
- Murai, N.; Kirkegaard, M.; Jarlebark, L.; Risling, M.; Suneson, A.; Ulfendahl, M. Activation of jnk in the inner ear following impulse noise exposure. J. Neurotrauma 2008, 25, 72–77.
- Seidman, M.D.; Shivapuja, B.G.; Quirk, W.S. The protective effects of allopurinol and superoxide dismutase on noise-induced cochlear damage. Otolaryngol. Head Neck Surg. 1993, 109, 1052–1056.
- Liu, Z. Experimental study on the mechanism of free radical in blast trauma induced hearing loss. Zhonghua Er Bi Yan Hou Ke Za Zhi 1992, 27, 24–26.
- Hamanaka, R.B.; Chandel, N.S. Mitochondrial reactive oxygen species regulate cellular signaling and dictate biological outcomes. Trends Biochem. Sci. 2010, 35, 505–513.
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. Tlr signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480.
- Myers, A.L.; Harris, C.M.; Choe, K.M.; Brennan, C.A. Inflammatory production of reactive oxygen species by drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun. 2018, 505, 726–732.
- Lambeth, J.D.; Neish, A.S. Nox enzymes and new thinking on reactive oxygen: A double-edged sword revisited. Annu. Rev. Pathol. 2014, 9, 119–145.
- Brandes, R.P.; Weissmann, N.; Schroder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226.
- Vermot, A.; Petit-Hartlein, I.; Smith, S.M.E.; Fieschi, F. NADPH oxidases (nox): An overview from discovery, molecular mechanisms to physiology and pathology. Antioxidants 2021, 10, 890.
- Costa, R.A.; Romagna, C.D.; Pereira, J.L.; Souza-Pinto, N.C. The role of mitochondrial DNA damage in the citotoxicity of reactive oxygen species. J. Bioenerg. Biomembr. 2011, 43, 25–29.
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta 1998, 1366, 139–149.
- Li, P.; Li, S.; Wang, L.; Li, H.; Wang, Y.; Liu, H.; Wang, X.; Zhu, X.; Liu, Z.; Ye, F.; et al. Mitochondrial dysfunction in hearing loss: Oxidative stress, autophagy and nlrp3 inflammasome. Front. Cell Dev. Biol. 2023, 11, 1119773.
- Pak, J.H.; Kim, Y.; Yi, J.; Chung, J.W. Antioxidant therapy against oxidative damage of the inner ear: Protection and preconditioning. Antioxidants 2020, 9, 1076.
- Kishimoto-Urata, M.; Urata, S.; Fujimoto, C.; Yamasoba, T. Role of oxidative stress and antioxidants in acquired inner ear disorders. Antioxidants 2022, 11, 1469.
- Fay, R.R. Comparative psychoacoustics. Hear. Res. 1988, 34, 295–305.
- Mohri, H.; Ninoyu, Y.; Sakaguchi, H.; Hirano, S.; Saito, N.; Ueyama, T. Nox3-derived superoxide in cochleae induces sensorineural hearing loss. J. Neurosci. 2021, 41, 4716–4731.
- Nacher-Soler, G.; Lenglet, S.; Coelho, M.; Thomas, A.; Voruz, F.; Krause, K.H.; Senn, P.; Rousset, F. Local cisplatin delivery in mouse reliably models sensorineural ototoxicity without systemic adverse effects. Front. Cell. Neurosci. 2021, 15, 701783.
- Mukherjea, D.; Jajoo, S.; Kaur, T.; Sheehan, K.E.; Ramkumar, V.; Rybak, L.P. Transtympanic administration of short interfering (si)rna for the nox3 isoform of NADPH oxidase protects against cisplatin-induced hearing loss in the rat. Antioxid. Redox Signal. 2010, 13, 589–598.
- Rousset, F.; Nacher-Soler, G.; Kokje, V.B.C.; Sgroi, S.; Coelho, M.; Krause, K.H.; Senn, P. NADPH oxidase 3 deficiency protects from noise-induced sensorineural hearing loss. Front. Cell Dev. Biol. 2022, 10, 832314.
- Zhang, Y.; Shan, P.; Srivastava, A.; Jiang, G.; Zhang, X.; Lee, P.J. An endothelial hsp70-tlr4 axis limits Nox3 expression and protects against oxidant injury in lungs. Antioxid. Redox Signal. 2016, 24, 991–1012.
- Zhang, X.; Shan, P.; Jiang, G.; Cohn, L.; Lee, P.J. Toll-like receptor 4 deficiency causes pulmonary emphysema. J. Clin. Investig. 2006, 116, 3050–3059.
- Nagamani, S.C.; Erez, A.; Eng, C.; Ou, Z.; Chinault, C.; Workman, L.; Coldwell, J.; Stankiewicz, P.; Patel, A.; Lupski, J.R.; et al. Interstitial deletion of 6q25.2-q25.3: A novel microdeletion syndrome associated with microcephaly, developmental delay, dysmorphic features and hearing loss. Eur. J. Hum. Genet. EJHG 2009, 17, 573–581.
- Milosevic, J.; Kalicanin, P. Long arm deletion of chromosome no. 6 in a mentally retarded boy with multiple physical malformations. J. Ment. Defic. Res. 1975, 19, 139–144.
- Hopkin, R.J.; Schorry, E.; Bofinger, M.; Milatovich, A.; Stern, H.J.; Jayne, C.; Saal, H.M. New insights into the phenotypes of 6q deletions. Am. J. Med. Genet. 1997, 70, 377–386.
- Sukumar, S.; Wang, S.; Hoang, K.; Vanchiere, C.M.; England, K.; Fick, R.; Pagon, B.; Reddy, K.S. Subtle overlapping deletions in the terminal region of chromosome 6q24.2–q26: Three cases studied using fish. Am. J. Med. Genet. 1999, 87, 17–22.
- Pandya, A.; Braverman, N.; Pyeritz, R.E.; Ying, K.L.; Kline, A.D.; Falk, R.E. Interstitial deletion of the long arm of chromosome 6 associated with unusual limb anomalies: Report of two new patients and review of the literature. Am. J. Med. Genet. 1995, 59, 38–43.
- Schuster, M.; Lohscheller, J.; Kummer, P.; Eysholdt, U.; Rosanowski, F. Severe sensory hearing loss in del(6q)-syndrome. Int. J. Pediatr. Otorhinolaryngol. 2003, 67, 1263–1266.
- Stucken, E.Z.; Hong, R.S. Noise-induced hearing loss: An occupational medicine perspective. Curr. Opin. Otolaryngol. Head Neck Surg. 2014, 22, 388–393.
- Dobie, R.A. The burdens of age-related and occupational noise-induced hearing loss in the united states. Ear Hear. 2008, 29, 565–577.
- Yankaskas, K. Prelude: Noise-induced tinnitus and hearing loss in the military. Hear. Res. 2013, 295, 3–8.
- Guski, R.; Schreckenberg, D.; Schuemer, R. Who environmental noise guidelines for the european region: A systematic review on environmental noise and annoyance. Int. J. Environ. Res. Public Health 2017, 14, 1539.
- Saffree Jeffree, M.; Ismail, N.; Awang Lukman, K. Hearing impairment and contributing factors among fertilizer factory workers. J. Occup. Health 2016, 58, 434–443.
- Habybabady, R.H.; Mohammadi, M.; Mortazavi, S.B.; Khavanin, A.; Mirzaei, R.; Malvajerdi, M.S. The effect of simultaneous exposure to cigarette smoke and noise on distortion product otoacoustic emissions in rats. Toxicol. Ind. Health 2019, 35, 349–357.
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 14077–14085.
- Kaur, T.; Clayman, A.C.; Nash, A.J.; Schrader, A.D.; Warchol, M.E.; Ohlemiller, K.K. Lack of fractalkine receptor on macrophages impairs spontaneous recovery of ribbon synapses after moderate noise trauma in c57bl/6 mice. Front. Neurosci. 2019, 13, 620.
- Pourbakht, A.; Yamasoba, T. Cochlear damage caused by continuous and intermittent noise exposure. Hear. Res. 2003, 178, 70–78.
- Op de Beeck, K.; Schacht, J.; Van Camp, G. Apoptosis in acquired and genetic hearing impairment: The programmed death of the hair cell. Hear. Res. 2011, 281, 18–27.
- Frederiksen, T.W.; Ramlau-Hansen, C.H.; Stokholm, Z.A.; Grynderup, M.B.; Hansen, A.M.; Kristiansen, J.; Vestergaard, J.M.; Bonde, J.P.; Kolstad, H.A. Noise-induced hearing loss—A preventable disease? Results of a 10-year longitudinal study of workers exposed to occupational noise. Noise Health 2017, 19, 103–111.
- Shargorodsky, J.; Curhan, S.G.; Curhan, G.C.; Eavey, R. Change in prevalence of hearing loss in us adolescents. JAMA 2010, 304, 772–778.
- Rybak, L.P.; Ramkumar, V. Ototoxicity. Kidney Int. 2007, 72, 931–935.
- Wang, X.; Xu, P.; Li, P.; Wang, Z.; Zhao, F.; Gao, Z.; Xu, L.; Luo, Y.J.; Fan, J.; Liu, P. Alterations in gray matter volume due to unilateral hearing loss. Sci. Rep. 2016, 6, 25811.
- Nakajima, K.; Kanda, E.; Hosobuchi, A.; Suwa, K. Subclinical hearing loss, longer sleep duration, and cardiometabolic risk factors in japanese general population. Int. J. Otolaryngol. 2014, 2014, 218218.
- Lie, A.; Skogstad, M.; Johannessen, H.A.; Tynes, T.; Mehlum, I.S.; Nordby, K.C.; Engdahl, B.; Tambs, K. Occupational noise exposure and hearing: A systematic review. Int. Arch. Occup. Environ. Health 2016, 89, 351–372.
- Sliwinska-Kowalska, M.; Pawelczyk, M. Contribution of genetic factors to noise-induced hearing loss: A human studies review. Mutat. Res. 2013, 752, 61–65.
- Heinonen-Guzejev, M.; Vuorinen, H.S.; Mussalo-Rauhamaa, H.; Heikkilä, K.; Koskenvuo, M.; Kaprio, J. Genetic component of noise sensitivity. Twin Res. Hum. Genet. 2012, 8, 245–249.
- Erway, L.C.; Shiau, Y.W.; Davis, R.R.; Krieg, E.F. Genetics of age-related hearing loss in mice. Iii. Susceptibility of inbred and f1 hybrid strains to noise-induced hearing loss. Hear. Res. 1996, 93, 181–187.
- Kozel, P.J.; Davis, R.R.; Krieg, E.F.; Shull, G.E.; Erway, L.C. Deficiency in plasma membrane calcium atpase isoform 2 increases susceptibility to noise-induced hearing loss in mice. Hear. Res. 2002, 164, 231–239.
- Holme, R.H.; Steel, K.P. Progressive hearing loss and increased susceptibility to noise-induced hearing loss in mice carrying a cdh23 but not a myo7a mutation. J. Assoc. Res. Otolaryngol. JARO 2004, 5, 66–79.
- Ramkumar, V.; Whitworth, C.A.; Pingle, S.C.; Hughes, L.F.; Rybak, L.P. Noise induces a1 adenosine receptor expression in the chinchilla cochlea. Hear. Res. 2004, 188, 47–56.
- Yang, W.; Vethanayagam, R.R.; Dong, Y.; Cai, Q.; Hu, B.H. Activation of the antigen presentation function of mononuclear phagocyte populations associated with the basilar membrane of the cochlea after acoustic overstimulation. Neuroscience 2015, 303, 1–15.
- Hashimoto, S.; Billings, P.; Harris, J.P.; Firestein, G.S.; Keithley, E.M. Innate immunity contributes to cochlear adaptive immune responses. Audiol. Neuro-Otol. 2005, 10, 35–43.
- Wakabayashi, K.; Fujioka, M.; Kanzaki, S.; Okano, H.J.; Shibata, S.; Yamashita, D.; Masuda, M.; Mihara, M.; Ohsugi, Y.; Ogawa, K.; et al. Blockade of interleukin-6 signaling suppressed cochlear inflammatory response and improved hearing impairment in noise-damaged mice cochlea. Neurosci. Res. 2010, 66, 345–352.
- Keithley, E.M.; Wang, X.; Barkdull, G.C. Tumor necrosis factor alpha can induce recruitment of inflammatory cells to the cochlea. Otol. Neurotol. 2008, 29, 854–859.
- Paciello, F.; Di Pino, A.; Rolesi, R.; Troiani, D.; Paludetti, G.; Grassi, C.; Fetoni, A.R. Anti-oxidant and anti-inflammatory effects of caffeic acid: In Vivo evidences in a model of noise-induced hearing loss. Food Chem. Toxicol. 2020, 143, 111555.
- Kurabi, A.; Keithley, E.M.; Housley, G.D.; Ryan, A.F.; Wong, A.C. Cellular mechanisms of noise-induced hearing loss. Hear. Res. 2017, 349, 129–137.
- Bennett, B.J.; Farber, C.R.; Orozco, L.; Kang, H.M.; Ghazalpour, A.; Siemers, N.; Neubauer, M.; Neuhaus, I.; Yordanova, R.; Guan, B.; et al. A high-resolution association mapping panel for the dissection of complex traits in mice. Genome Res. 2010, 20, 281–290.
- Farber, C.R.; Bennett, B.J.; Orozco, L.; Zou, W.; Lira, A.; Kostem, E.; Kang, H.M.; Furlotte, N.; Berberyan, A.; Ghazalpour, A.; et al. Mouse genome-wide association and systems genetics identify asxl2 as a regulator of bone mineral density and osteoclastogenesis. PLoS Genet. 2011, 7, e1002038.
- Park, C.C.; Gale, G.D.; de Jong, S.; Ghazalpour, A.; Bennett, B.J.; Farber, C.R.; Langfelder, P.; Lin, A.; Khan, A.H.; Eskin, E.; et al. Gene networks associated with conditional fear in mice identified using a systems genetics approach. BMC Syst. Biol. 2011, 5, 43.
- Lavinsky, J.; Crow, A.L.; Pan, C.; Wang, J.; Aaron, K.A.; Ho, M.K.; Li, Q.; Salehide, P.; Myint, A.; Monges-Hernadez, M.; et al. Genome-wide association study identifies Nox3 as a critical gene for susceptibility to noise-induced hearing loss. PLoS Genet. 2015, 11, e1005094.
- Zhao, T.; Wang, Y.; Li, Z.; Xu, X.; Lei, S.; Huang, L.; Xu, L.; Zhang, M.; Yang, L. Associations of noise kurtosis, genetic variations in nox3 and lifestyle factors with noise-induced hearing loss. Environ. Health A Glob. Access Sci. Source 2020, 19, 13.
- Lei, S.F.; Ahroon, W.A.; Hamernik, R.P. The application of frequency and time domain kurtosis to the assessment of hazardous noise exposures. J. Acoust. Soc. Am. 1994, 96, 1435–1444.
- Zhao, Y.M.; Qiu, W.; Zeng, L.; Chen, S.S.; Cheng, X.R.; Davis, R.I.; Hamernik, R.P. Application of the kurtosis statistic to the evaluation of the risk of hearing loss in workers exposed to high-level complex noise. Ear Hear. 2010, 31, 527–532.
- Sato, S.; Kitamura, T.; Sakai, H.; Ando, Y. The loudness of “complex noise” in relation to the factors extracted from the auto-correlation function. J. Sound Vib. 2001, 241, 97–103.
- Erdreich, J. A distribution based definition of impulse noise. J. Acoust. Soc. Am. 1986, 79, 990–998.
- Xie, H.W.; Qiu, W.; Heyer, N.J.; Zhang, M.B.; Zhang, P.; Zhao, Y.M.; Hamernik, R.P. The use of the kurtosis-adjusted cumulative noise exposure metric in evaluating the hearing loss risk for complex noise. Ear Hear. 2016, 37, 312–323.
- Davis, R.I.; Qiu, W.; Heyer, N.J.; Zhao, Y.; Qiuling Yang, M.S.; Li, N.; Tao, L.; Zhu, L.; Zeng, L.; Yao, D. The use of the kurtosis metric in the evaluation of occupational hearing loss in workers in china: Implications for hearing risk assessment. Noise Health 2012, 14, 330–342.
- Banfi, B.; Malgrange, B.; Knisz, J.; Steger, K.; Dubois-Dauphin, M.; Krause, K.H. Nox3, a superoxide-generating NADPH oxidase of the inner ear. J. Biol. Chem. 2004, 279, 46065–46072.
- Flaherty, J.P.; Fairfield, H.E.; Spruce, C.A.; McCarty, C.M.; Bergstrom, D.E. Molecular characterization of an allelic series of mutations in the mouse Nox3 gene. Mamm. Genome 2011, 22, 156–169.
- Du, Z.; Li, S.; Liu, L.; Yang, Q.; Zhang, H.; Gao, C. NADPH oxidase 3-associated oxidative stress and caspase 3-dependent apoptosis in the cochleae of d-galactose-induced aged rats. Mol. Med. Rep. 2015, 12, 7883–7890.
- Vlajkovic, S.M.; Lin, S.C.; Wong, A.C.; Wackrow, B.; Thorne, P.R. Noise-induced changes in expression levels of NADPH oxidases in the cochlea. Hear. Res. 2013, 304, 145–152.
- Dhukhwa, A.; Bhatta, P.; Sheth, S.; Korrapati, K.; Tieu, C.; Mamillapalli, C.; Ramkumar, V.; Mukherjea, D. Targeting inflammatory processes mediated by trpvi and tnf-alpha for treating noise-induced hearing loss. Front. Cell. Neurosci. 2019, 13, 444.
- Kaur, T.; Borse, V.; Sheth, S.; Sheehan, K.; Ghosh, S.; Tupal, S.; Jajoo, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Adenosine a1 receptor protects against cisplatin ototoxicity by suppressing the nox3/stat1 inflammatory pathway in the cochlea. J. Neurosci. 2016, 36, 3962–3977.
- Nakano, Y.; Banfi, B.; Jesaitis, A.J.; Dinauer, M.C.; Allen, L.A.; Nauseef, W.M. Critical roles for p22phox in the structural maturation and subcellular targeting of Nox3. Biochem. J. 2007, 403, 97–108.
- Paffenholz, R.; Bergstrom, R.A.; Pasutto, F.; Wabnitz, P.; Munroe, R.J.; Jagla, W.; Heinzmann, U.; Marquardt, A.; Bareiss, A.; Laufs, J.; et al. Vestibular defects in head-tilt mice result from mutations in Nox3, encoding an NADPH oxidase. Genes Dev. 2004, 18, 486–491.
- Kiss, P.J.; Knisz, J.; Zhang, Y.; Baltrusaitis, J.; Sigmund, C.D.; Thalmann, R.; Smith, R.J.; Verpy, E.; Banfi, B. Inactivation of NADPH oxidase organizer 1 results in severe imbalance. Curr. Biol. CB 2006, 16, 208–213.
- Zahra, G.; Esmaeil, K.; Mohammad, F.; Rashidy-Pour, A.; Mahdi, M.; Mahdi, A.; Ali, K. Combined effects of the exposure to silver nanoparticles and noise on hearing function and cochlea structure of the male rats. Life Sci. 2022, 304, 120724.
- Peralta-Videa, J.R.; Zhao, L.; Lopez-Moreno, M.L.; de la Rosa, G.; Hong, J.; Gardea-Torresdey, J.L. Nanomaterials and the environment: A review for the biennium 2008–2010. J. Hazard. Mater. 2011, 186, 1–15.
- Roda, E.; Bottone, M.G.; Biggiogera, M.; Milanesi, G.; Coccini, T. Pulmonary and hepatic effects after low dose exposure to nanosilver: Early and long-lasting histological and ultrastructural alterations in rat. Toxicol. Rep. 2019, 6, 1047–1060.
- Recordati, C.; De Maglie, M.; Bianchessi, S.; Argentiere, S.; Cella, C.; Mattiello, S.; Cubadda, F.; Aureli, F.; D’Amato, M.; Raggi, A. Tissue distribution and acute toxicity of silver after single intravenous administration in mice: Nano-specific and size-dependent effects. Part. Fibre Toxicol. 2015, 13, 12.
- Xu, L.; Li, X.; Takemura, T.; Hanagata, N.; Wu, G.; Chou, L.L. Genotoxicity and molecular response of silver nanoparticle (np)-based hydrogel. J. Nanobiotechnol. 2012, 10, 16.
- Ferdous, Z.; Nemmar, A. Health impact of silver nanoparticles: A review of the biodistribution and toxicity following various routes of exposure. Int. J. Mol. Sci. 2020, 21, 2375.
- AshaRani, P.V.; Low Kah Mun, G.; Hande, M.P.; Valiyaveettil, S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 2009, 3, 279–290.
- Kawata, K.; Osawa, M.; Okabe, S. In vitro toxicity of silver nanoparticles at noncytotoxic doses to hepg2 human hepatoma cells. Environ. Sci. Technol. 2009, 43, 6046–6051.
- Kim, H.R.; Kim, M.J.; Lee, S.Y.; Oh, S.M.; Chung, K.H. Genotoxic effects of silver nanoparticles stimulated by oxidative stress in human normal bronchial epithelial (beas-2b) cells. Mutat. Res. 2011, 726, 129–135.
- Neely, S.T.; Johnson, T.A.; Kopun, J.; Dierking, D.M.; Gorga, M.P. Distortion-product otoacoustic emission input/output characteristics in normal-hearing and hearing-impaired human ears. J. Acoust. Soc. Am. 2009, 126, 728–738.
- Martin, G.K.; Stagner, B.B.; Chung, Y.S.; Lonsbury-Martin, B.L. Characterizing distortion-product otoacoustic emission components across four species. J. Acoust. Soc. Am. 2011, 129, 3090–3103.
- Shahtaheri, S.J.; Goodarzi, Z.; Karami, E.; Khavanin, A.; Khansari, M.G.; Kiani, M.; Rashidy-Pour, A. Effects of acute exposure to al(2)o(3)-nps (alpha and gamma) and white noise and their combination on cochlea structure and function in wistar rats. Environ. Sci. Pollut. Res. Int. 2023, 30, 89859–89876.
- Ates, M.; Demir, V.; Arslan, Z.; Daniels, J.; Farah, I.O.; Bogatu, C. Evaluation of alpha and gamma aluminum oxide nanoparticle accumulation, toxicity, and depuration in artemia salina larvae. Environ. Toxicol. 2015, 30, 109–118.
- Patlolla, A.K.; Kumari, S.A.; Madhusudhanachary, P.; Turner, T.; Tchounwou, P.B. Biochemical and histopathological evaluation of al(2)o(3) nanomaterials in kidney of wistar rats. Curr. Top. Biochem. Res. 2018, 19, 1–12.
- Jacukowicz-Sobala, I.; Ocinski, D.; Kociolek-Balawejder, E. Iron and aluminium oxides containing industrial wastes as adsorbents of heavy metals: Application possibilities and limitations. Waste Manag. Res. 2015, 33, 612–629.
- Brown, D.M.; Brown, A.M.; Willitsford, A.H.; Dinello-Fass, R.; Airola, M.B.; Siegrist, K.M.; Thomas, M.E.; Chang, Y. Lidar measurements of solid rocket propellant fire particle plumes. Appl. Opt. 2016, 55, 4657–4669.
- Hunter, D.; Milton, R.; Perry, K.M.A.; Thompson, D.R. Effect of aluminium and alumina on the lung in grinders of duralumin aeroplane propellers. Br. J. Ind. Med. 1944, 1, 159–164.
- Sikkeland, L.; Alexis, N.E.; Fry, R.C.; Martin, E.; Danielsen, T.E.; Sostrand, P.; Kongerud, J. Inflammation in induced sputum after aluminium oxide exposure: An experimental chamber study. Occup. Environ. Med. 2016, 73, 199–205.
- Xing, M.; Zou, H.; Gao, X.; Chang, B.; Tang, S.; Zhang, M. Workplace exposure to airborne alumina nanoparticles associated with separation and packaging processes in a pilot factory. Environ. Sci. Process Impacts 2015, 17, 656–666.
- Lu, J.; Li, W.; Du, X.; Ewert, D.L.; West, M.B.; Stewart, C.; Floyd, R.A.; Kopke, R.D. Antioxidants reduce cellular and functional changes induced by intense noise in the inner ear and cochlear nucleus. J. Assoc. Res. Otolaryngol. JARO 2014, 15, 353–372.
- Fram, R.J. Cisplatin and platinum analogues: Recent advances. Curr. Opin. Oncol. 1992, 4, 1073–1079.
- Boulikas, T.; Vougiouka, M. Recent clinical trials using cisplatin, carboplatin and their combination chemotherapy drugs (review). Oncol. Rep. 2004, 11, 559–595.
- Laurell, G. Pharmacological intervention in the field of ototoxicity. Hno 2019, 67, 434–439.
- Wang, D.; Lippard, S.J. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov. 2005, 4, 307–320.
- Jordan, J.A.; Schwade, N.D.; Truelson, J.M. Fosfomycin does not inhibit the tumoricidal efficacy of cisplatinum. Laryngoscope 1999, 109, 1259–1262.
- Saleh, S.; El-Demerdash, E. Protective effects of l-arginine against cisplatin-induced renal oxidative stress and toxicity: Role of nitric oxide. Basic Clin. Pharmacol. Toxicol. 2005, 97, 91–97.
- Bodenner, D.L.; Dedon, P.C.; Keng, P.C.; Borch, R.F. Effect of diethyldithiocarbamate on cis-diamminedichloroplatinum(ii)-induced cytotoxicity, DNA cross-linking, and gamma-glutamyl transpeptidase inhibition. Cancer Res. 1986, 46, 2745–2750.
- Barabas, K.; Milner, R.; Lurie, D.; Adin, C. Cisplatin: A review of toxicities and therapeutic applications. Vet. Comp. Oncol. 2008, 6, 1–18.
- Hartmann, J.T.; Lipp, H.P. Toxicity of platinum compounds. Expert Opin. Pharmacother. 2003, 4, 889–901.
- Schacht, J.; Talaska, A.E.; Rybak, L.P. Cisplatin and aminoglycoside antibiotics: Hearing loss and its prevention. Anat. Rec. 2012, 295, 1837–1850.
- Koike, N.; Sasaki, A.; Murakami, T.; Suzuki, K. Effect of edaravone against cisplatin-induced chronic renal injury. Drug Chem. Toxicol. 2021, 44, 437–446.
- Thadhani, R.; Pascual, M.; Bonventre, J.V. Acute renal failure. N. Engl. J. Med. 1996, 334, 1448–1460.
- Rybak, L.P.; Mukherjea, D.; Jajoo, S.; Ramkumar, V. Cisplatin ototoxicity and protection: Clinical and experimental studies. Tohoku J. Exp. Med. 2009, 219, 177–186.
- McKeage, M.J. Comparative adverse effect profiles of platinum drugs. Drug Saf. 1995, 13, 228–244.
- Bokemeyer, C.; Berger, C.C.; Hartmann, J.T.; Kollmannsberger, C.; Schmoll, H.J.; Kuczyk, M.A.; Kanz, L. Analysis of risk factors for cisplatin-induced ototoxicity in patients with testicular cancer. Br. J. Cancer 1998, 77, 1355–1362.
- Huang, E.; Teh, B.S.; Strother, D.R.; Davis, Q.G.; Chiu, J.K.; Lu, H.H.; Carpenter, L.S.; Mai, W.Y.; Chintagumpala, M.M.; South, M.; et al. Intensity-modulated radiation therapy for pediatric medulloblastoma: Early report on the reduction of ototoxicity. Int. J. Radiat. Oncol. Biol. Phys. 2002, 52, 599–605.
- Anniko, M.; Sobin, A. Cisplatin: Evaluation of its ototoxic potential. Am. J. Otolaryngol. 1986, 7, 276–293.
- Tsukasaki, N.; Whitworth, C.A.; Rybak, L.P. Acute changes in cochlear potentials due to cisplatin. Hear. Res. 2000, 149, 189–198.
- Rybak, L.P.; Kelly, T. Ototoxicity: Bioprotective mechanisms. Curr. Opin. Otolaryngol. Head Neck Surg. 2003, 11, 328–333.
- Watanabe, K.I.; Hess, A.; Bloch, W.; Michel, O. Nitric oxide synthase inhibitor suppresses the ototoxic side effect of cisplatin in guinea pigs. Anticancer Drugs 2000, 11, 401–406.
- Hong, O.; Kerr, M.J.; Poling, G.L.; Dhar, S. Understanding and preventing noise-induced hearing loss. Dis.-A-Mon. DM 2013, 59, 110–118.
- Kopke, R.D.; Liu, W.; Gabaizadeh, R.; Jacono, A.; Feghali, J.; Spray, D.; Garcia, P.; Steinman, H.; Malgrange, B.; Ruben, R.J.; et al. Use of organotypic cultures of corti’s organ to study the protective effects of antioxidant molecules on cisplatin-induced damage of auditory hair cells. Am. J. Otol. 1997, 18, 559–571.
- Rybak, L.P.; Husain, K.; Whitworth, C.; Somani, S.M. Dose dependent protection by lipoic acid against cisplatin-induced ototoxicity in rats: Antioxidant defense system. Toxicol. Sci. Off. J. Soc. Toxicol. 1999, 47, 195–202.
- Waissbluth, S.; Salehi, P.; He, X.; Daniel, S.J. Systemic dexamethasone for the prevention of cisplatin-induced ototoxicity. Eur. Arch. Oto-Rhino-Laryngol. 2013, 270, 1597–1605.
- Rybak, L.P. Cis-platinum associated hearing loss. J. Laryngol. Otol. 1981, 95, 745–747.
- Fleischman, R.W.; Stadnicki, S.W.; Ethier, M.F.; Schaeppi, U. Ototoxicity of cis-dichlorodiammine platinum (ii) in the guinea pig. Toxicol. Appl. Pharmacol. 1975, 33, 320–332.
- Stadnicki, S.W.; Fleischman, R.W.; Schaeppi, U.; Merriam, P. Cis-dichlorodiammineplatinum (ii) (nsc-119875): Hearing loss and other toxic effects in rhesus monkeys. Cancer Chemother. Rep. 1975, 59, 467–480.
- Staecker, H.; Liu, W.; Malgrange, B.; Lefebvre, P.P.; Van De Water, T.R. Vector-mediated delivery of bcl-2 prevents degeneration of auditory hair cells and neurons after injury. ORL 2007, 69, 43–50.
- Zhang, N.; Cai, J.; Xu, L.; Wang, H.; Liu, W. Cisplatin-induced stria vascularis damage is associated with inflammation and fibrosis. Neural Plast. 2020, 2020, 8851525.
- Sluyter, S.; Klis, S.F.; de Groot, J.C.; Smoorenburg, G.F. Alterations in the stria vascularis in relation to cisplatin ototoxicity and recovery. Hear. Res. 2003, 185, 49–56.
- Kanzaki, J.; Ouchi, T. Steroid-responsive bilateral sensorineural hearing loss and immune complexes. Arch. Oto-Rhino-Laryngol. 1981, 230, 5–9.
- Yoshida, K.; Ichimiya, I.; Suzuki, M.; Mogi, G. Effect of proinflammatory cytokines on cultured spiral ligament fibrocytes. Hear. Res. 1999, 137, 155–159.
- Satoh, H.; Firestein, G.S.; Billings, P.B.; Harris, J.P.; Keithley, E.M. Proinflammatory cytokine expression in the endolymphatic sac during inner ear inflammation. J. Assoc. Res. Otolaryngol. JARO 2003, 4, 139–147.
- Kim, H.J.; So, H.S.; Lee, J.H.; Park, C.; Lee, J.B.; Youn, M.J.; Kim, S.J.; Yang, S.H.; Lee, K.M.; Kwon, K.B.; et al. Role of proinflammatory cytokines in cisplatin-induced vestibular hair cell damage. Head Neck 2008, 30, 1445–1456.
- So, H.; Kim, H.; Kim, Y.; Kim, E.; Pae, H.O.; Chung, H.T.; Kim, H.J.; Kwon, K.B.; Lee, K.M.; Lee, H.Y.; et al. Evidence that cisplatin-induced auditory damage is attenuated by downregulation of pro-inflammatory cytokines via nrf2/ho-1. J. Assoc. Res. Otolaryngol. JARO 2008, 9, 290–306.
- Watanabe, K.; Inai, S.; Jinnouchi, K.; Bada, S.; Hess, A.; Michel, O.; Yagi, T. Nuclear-factor kappa b (nf-kappa b)-inducible nitric oxide synthase (inos/nos ii) pathway damages the stria vascularis in cisplatin-treated mice. Anticancer. Res. 2002, 22, 4081–4085.
- Ries, F.; Klastersky, J. Nephrotoxicity induced by cancer chemotherapy with special emphasis on cisplatin toxicity. Am. J. Kidney Dis. 1986, 8, 368–379.
- Leibbrandt, M.E.; Wolfgang, G.H.; Metz, A.L.; Ozobia, A.A.; Haskins, J.R. Critical subcellular targets of cisplatin and related platinum analogs in rat renal proximal tubule cells. Kidney Int. 1995, 48, 761–770.
- Sugiyama, S.; Hayakawa, M.; Kato, T.; Hanaki, Y.; Shimizu, K.; Ozawa, T. Adverse effects of anti-tumor drug, cisplatin, on rat kidney mitochondria: Disturbances in glutathione peroxidase activity. Biochem. Biophys. Res. Commun. 1989, 159, 1121–1127.
- So, H.S.; Park, C.; Kim, H.J.; Lee, J.H.; Park, S.Y.; Lee, J.H.; Lee, Z.W.; Kim, H.M.; Kalinec, F.; Lim, D.J.; et al. Protective effect of t-type calcium channel blocker flunarizine on cisplatin-induced death of auditory cells. Hear. Res. 2005, 204, 127–139.
- Goncalves, M.S.; Silveira, A.F.; Teixeira, A.R.; Hyppolito, M.A. Mechanisms of cisplatin ototoxicity: Theoretical review. J. Laryngol. Otol. 2013, 127, 536–541.
- Matsushima, H.; Yonemura, K.; Ohishi, K.; Hishida, A. The role of oxygen free radicals in cisplatin-induced acute renal failure in rats. J. Lab. Clin. Med. 1998, 131, 518–526.
- Rybak, L.P.; Whitworth, C.; Somani, S. Application of antioxidants and other agents to prevent cisplatin ototoxicity. Laryngoscope 1999, 109, 1740–1744.
- Sergi, B.; Ferraresi, A.; Troiani, D.; Paludetti, G.; Fetoni, A.R. Cisplatin ototoxicity in the guinea pig: Vestibular and cochlear damage. Hear. Res. 2003, 182, 56–64.
- Takumida, M.; Anniko, M. Simultaneous detection of both nitric oxide and reactive oxygen species in guinea pig vestibular sensory cells. ORL 2002, 64, 143–147.
- Darlington, C.L.; Smith, P.F. Vestibulotoxicity following aminoglycoside antibiotics and its prevention. Curr. Opin. Investig. Drugs 2003, 4, 841–846.
- Rivolta, M.N.; Grix, N.; Lawlor, P.; Ashmore, J.F.; Jagger, D.J.; Holley, M.C. Auditory hair cell precursors immortalized from the mammalian inner ear. Proceedings. Biol. Sci. 1998, 265, 1595–1603.
- Mukherjea, D.; Jajoo, S.; Whitworth, C.; Bunch, J.R.; Turner, J.G.; Rybak, L.P.; Ramkumar, V. Short interfering rna against transient receptor potential vanilloid 1 attenuates cisplatin-induced hearing loss in the rat. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 13056–13065.
- Kim, H.J.; Lee, J.H.; Kim, S.J.; Oh, G.S.; Moon, H.D.; Kwon, K.B.; Park, C.; Park, B.H.; Lee, H.K.; Chung, S.Y.; et al. Roles of NADPH oxidases in cisplatin-induced reactive oxygen species generation and ototoxicity. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 3933–3946.
- Mukherjea, D.; Whitworth, C.A.; Nandish, S.; Dunaway, G.A.; Rybak, L.P.; Ramkumar, V. Expression of the kidney injury molecule 1 in the rat cochlea and induction by cisplatin. Neuroscience 2006, 139, 733–740.
- Benkafadar, N.; Menardo, J.; Bourien, J.; Nouvian, R.; Francois, F.; Decaudin, D.; Maiorano, D.; Puel, J.L.; Wang, J. Reversible p53 inhibition prevents cisplatin ototoxicity without blocking chemotherapeutic efficacy. EMBO Mol. Med. 2017, 9, 7–26.
- Fredholm, B.B.; AP, I.J.; Jacobson, K.A.; Linden, J.; Muller, C.E. International union of basic and clinical pharmacology. Lxxxi. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34.
- Ramkumar, V.; Ravi, R.; Wilson, M.C.; Gettys, T.W.; Whitworth, C.; Rybak, L.P. Identification of a1 adenosine receptors in rat cochlea coupled to inhibition of adenylyl cyclase. Am. J. Physiol. 1994, 267, C731–C737.
- Ford, M.S.; Nie, Z.; Whitworth, C.; Rybak, L.P.; Ramkumar, V. Up-regulation of adenosine receptors in the cochlea by cisplatin. Hear. Res. 1997, 111, 143–152.
- Ford, M.S.; Maggirwar, S.B.; Rybak, L.P.; Whitworth, C.; Ramkumar, V. Expression and function of adenosine receptors in the chinchilla cochlea. Hear. Res. 1997, 105, 130–140.
- Bryant, G.M.; Barron, S.E.; Norris, C.H.; Guth, P.S. Adenosine is a modulator of hair cell-afferent neurotransmission. Hear. Res. 1987, 30, 231–237.
- Nario, K.; Kitano, I.; Mori, N.; Matsunaga, T. The effect of adenosine on cochlear potentials in the guinea pig. Eur. Arch. Oto-Rhino-Laryngol. 1994, 251, 428–433.
- Hight, N.G.; McFadden, S.L.; Henderson, D.; Burkard, R.F.; Nicotera, T. Noise-induced hearing loss in chinchillas pre-treated with glutathione monoethylester and r-pia. Hear. Res. 2003, 179, 21–32.
- Vlajkovic, S.M.; Guo, C.X.; Telang, R.; Wong, A.C.; Paramananthasivam, V.; Boison, D.; Housley, G.D.; Thorne, P.R. Adenosine kinase inhibition in the cochlea delays the onset of age-related hearing loss. Exp. Gerontol. 2011, 46, 905–914.
- Whitworth, C.A.; Ramkumar, V.; Jones, B.; Tsukasaki, N.; Rybak, L.P. Protection against cisplatin ototoxicity by adenosine agonists. Biochem. Pharmacol. 2004, 67, 1801–1807.
- Kaur, T.; Mukherjea, D.; Sheehan, K.; Jajoo, S.; Rybak, L.P.; Ramkumar, V. Short interfering rna against stat1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death Dis. 2011, 2, e180.
- Demmler, G.J. Infectious diseases society of america and centers for disease control. Summary of a workshop on surveillance for congenital cytomegalovirus disease. Rev. Infect. Dis. 1991, 13, 315–329.
- Stagno, S.; Pass, R.F.; Cloud, G.; Britt, W.J.; Henderson, R.E.; Walton, P.D.; Veren, D.A.; Page, F.; Alford, C.A. Primary cytomegalovirus infection in pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA 1986, 256, 1904–1908.
- Barbi, M.; Binda, S.; Caroppo, S.; Primache, V. Neonatal screening for congenital cytomegalovirus infection and hearing loss. J. Clin. Virol. 2006, 35, 206–209.
- Dreher, A.M.; Arora, N.; Fowler, K.B.; Novak, Z.; Britt, W.J.; Boppana, S.B.; Ross, S.A. Spectrum of disease and outcome in children with symptomatic congenital cytomegalovirus infection. J. Pediatr. 2014, 164, 855–859.
- Shi, X.; Dong, Y.; Li, Y.; Zhao, Z.; Li, H.; Qiu, S.; Li, Y.; Guo, W.; Qiao, Y. Inflammasome activation in mouse inner ear in response to mcmv induced hearing loss. J. Otol. 2015, 10, 143–149.
- Zhuang, W.; Wang, C.; Shi, X.; Qiu, S.; Zhang, S.; Xu, B.; Chen, M.; Jiang, W.; Dong, H.; Qiao, Y. Mcmv triggers ros/nlrp3-associated inflammasome activation in the inner ear of mice and cultured spiral ganglion neurons, contributing to sensorineural hearing loss. Int. J. Mol. Med. 2018, 41, 3448–3456.
- Schachtele, S.J.; Mutnal, M.B.; Schleiss, M.R.; Lokensgard, J.R. Cytomegalovirus-induced sensorineural hearing loss with persistent cochlear inflammation in neonatal mice. J. Neurovirol. 2011, 17, 201–211.
- Neag, M.A.; Mocan, A.; Echeverria, J.; Pop, R.M.; Bocsan, C.I.; Crisan, G.; Buzoianu, A.D. Berberine: Botanical occurrence, traditional uses, extraction methods, and relevance in cardiovascular, metabolic, hepatic, and renal disorders. Front. Pharmacol. 2018, 9, 557.
- Zhuang, W.; Li, T.; Wang, C.; Shi, X.; Li, Y.; Zhang, S.; Zhao, Z.; Dong, H.; Qiao, Y. Berberine exerts antioxidant effects via protection of spiral ganglion cells against cytomegalovirus-induced apoptosis. Free Radic. Biol. Med. 2018, 121, 127–135.
- Katsarkas, A.; Ayukawa, H. Hearing loss due to aging (presbycusis). J. Otolaryngol. 1986, 15, 239–244.
- Gennis, V.; Garry, P.J.; Haaland, K.Y.; Yeo, R.A.; Goodwin, J.S. Hearing and cognition in the elderly. New findings and a review of the literature. Arch. Intern. Med. 1991, 151, 2259–2264.
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120.
- Panza, F.; Solfrizzi, V.; Logroscino, G. Age-related hearing impairment-a risk factor and frailty marker for dementia and ad. Nat. Rev. Neurol. 2015, 11, 166–175.
- Rutherford, B.R.; Brewster, K.; Golub, J.S.; Kim, A.H.; Roose, S.P. Sensation and psychiatry: Linking age-related hearing loss to late-life depression and cognitive decline. Am. J. Psychiatry 2018, 175, 215–224.
- Yang, C.H.; Schrepfer, T.; Schacht, J. Age-related hearing impairment and the triad of acquired hearing loss. Front. Cell. Neurosci. 2015, 9, 276.
- Scholtz, A.W.; Kammen-Jolly, K.; Felder, E.; Hussl, B.; Rask-Andersen, H.; Schrott-Fischer, A. Selective aspects of human pathology in high-tone hearing loss of the aging inner ear. Hear. Res. 2001, 157, 77–86.
- Chen, B.; Zhong, Y.; Peng, W.; Sun, Y.; Kong, W.J. Age-related changes in the central auditory system: Comparison of d-galactose-induced aging rats and naturally aging rats. Brain Res. 2010, 1344, 43–53.
- Chen, B.; Zhong, Y.; Peng, W.; Sun, Y.; Hu, Y.J.; Yang, Y.; Kong, W.J. Increased mitochondrial DNA damage and decreased base excision repair in the auditory cortex of d-galactose-induced aging rats. Mol. Biol. Rep. 2011, 38, 3635–3642.
- Du, Z.; Yang, Y.; Hu, Y.; Sun, Y.; Zhang, S.; Peng, W.; Zhong, Y.; Huang, X.; Kong, W. A long-term high-fat diet increases oxidative stress, mitochondrial damage and apoptosis in the inner ear of d-galactose-induced aging rats. Hear. Res. 2012, 287, 15–24.
- Cui, X.; Zuo, P.; Zhang, Q.; Li, X.; Hu, Y.; Long, J.; Packer, L.; Liu, J. Chronic systemic d-galactose exposure induces memory loss, neurodegeneration, and oxidative damage in mice: Protective effects of r-alpha-lipoic acid. J. Neurosci. Res. 2006, 83, 1584–1590.
- Hua, X.; Lei, M.; Zhang, Y.; Ding, J.; Han, Q.; Hu, G.; Xiao, M. Long-term d-galactose injection combined with ovariectomy serves as a new rodent model for alzheimer’s disease. Life Sci. 2007, 80, 1897–1905.
- Lu, J.; Zheng, Y.L.; Wu, D.M.; Luo, L.; Sun, D.X.; Shan, Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by d-galactose. Biochem. Pharmacol. 2007, 74, 1078–1090.
- Chen, C.F.; Lang, S.Y.; Zuo, P.P.; Yang, N.; Wang, X.Q.; Xia, C. Effects of d-galactose on the expression of hippocampal peripheral-type benzodiazepine receptor and spatial memory performances in rats. Psychoneuroendocrinology 2006, 31, 805–811.
- Kumar, A.; Prakash, A.; Dogra, S. Naringin alleviates cognitive impairment, mitochondrial dysfunction and oxidative stress induced by d-galactose in mice. Food Chem. Toxicol. 2010, 48, 626–632.
- Lu, J.; Wu, D.M.; Zheng, Y.L.; Hu, B.; Zhang, Z.F. Purple sweet potato color alleviates d-galactose-induced brain aging in old mice by promoting survival of neurons via pi3k pathway and inhibiting cytochrome c-mediated apoptosis. Brain Pathol. 2010, 20, 598–612.
- Zhang, Z.F.; Lu, J.; Zheng, Y.L.; Hu, B.; Fan, S.H.; Wu, D.M.; Zheng, Z.H.; Shan, Q.; Liu, C.M. Purple sweet potato color protects mouse liver against d-galactose-induced apoptosis via inhibiting caspase-3 activation and enhancing pi3k/akt pathway. Food Chem. Toxicol. 2010, 48, 2500–2507.
- Lei, M.; Hua, X.; Xiao, M.; Ding, J.; Han, Q.; Hu, G. Impairments of astrocytes are involved in the d-galactose-induced brain aging. Biochem. Biophys. Res. Commun. 2008, 369, 1082–1087.
- Hsieh, H.M.; Wu, W.M.; Hu, M.L. Soy isoflavones attenuate oxidative stress and improve parameters related to aging and alzheimer’s disease in c57bl/6j mice treated with d-galactose. Food Chem. Toxicol. 2009, 47, 625–632.
- Cui, X.; Wang, L.; Zuo, P.; Han, Z.; Fang, Z.; Li, W.; Liu, J. D-galactose-caused life shortening in drosophila melanogaster and musca domestica is associated with oxidative stress. Biogerontology 2004, 5, 317–325.
- Kong, W.J.; Wang, Y.; Wang, Q.; Hu, Y.J.; Han, Y.C.; Liu, J. The relation between d-galactose injection and mitochondrial DNA 4834 bp deletion mutation. Exp. Gerontol. 2006, 41, 628–634.
- Kong, W.J.; Hu, Y.J.; Wang, Q.; Wang, Y.; Han, Y.C.; Cheng, H.M.; Kong, W.; Guan, M.X. The effect of the mtdna4834 deletion on hearing. Biochem. Biophys. Res. Commun. 2006, 344, 425–430.
- Zhong, Y.; Hu, Y.J.; Yang, Y.; Peng, W.; Sun, Y.; Chen, B.; Huang, X.; Kong, W.J. Contribution of common deletion to total deletion burden in mitochondrial DNA from inner ear of d-galactose-induced aging rats. Mutat. Res. 2011, 712, 11–19.
- Zhong, Y.; Hu, Y.J.; Chen, B.; Peng, W.; Sun, Y.; Yang, Y.; Zhao, X.Y.; Fan, G.R.; Huang, X.; Kong, W.J. Mitochondrial transcription factor a overexpression and base excision repair deficiency in the inner ear of rats with d-galactose-induced aging. FEBS J. 2011, 278, 2500–2510.
- Yamasoba, T.; Someya, S.; Yamada, C.; Weindruch, R.; Prolla, T.A.; Tanokura, M. Role of mitochondrial dysfunction and mitochondrial DNA mutations in age-related hearing loss. Hear. Res. 2007, 226, 185–193.
- Druzhyna, N.M.; Wilson, G.L.; LeDoux, S.P. Mitochondrial DNA repair in aging and disease. Mech. Ageing Dev. 2008, 129, 383–390.
- Meissner, C.; Bruse, P.; Mohamed, S.A.; Schulz, A.; Warnk, H.; Storm, T.; Oehmichen, M. The 4977 bp deletion of mitochondrial DNA in human skeletal muscle, heart and different areas of the brain: A useful biomarker or more? Exp. Gerontol. 2008, 43, 645–652.
- Markaryan, A.; Nelson, E.G.; Hinojosa, R. Quantification of the mitochondrial DNA common deletion in presbycusis. Laryngoscope 2009, 119, 1184–1189.
- Hiona, A.; Leeuwenburgh, C. The role of mitochondrial DNA mutations in aging and sarcopenia: Implications for the mitochondrial vicious cycle theory of aging. Exp. Gerontol. 2008, 43, 24–33.
- Basu, S.; Xie, X.; Uhler, J.P.; Hedberg-Oldfors, C.; Milenkovic, D.; Baris, O.R.; Kimoloi, S.; Matic, S.; Stewart, J.B.; Larsson, N.G.; et al. Accurate mapping of mitochondrial DNA deletions and duplications using deep sequencing. PLoS Genet. 2020, 16, e1009242.
- Schon, E.A.; Rizzuto, R.; Moraes, C.T.; Nakase, H.; Zeviani, M.; DiMauro, S. A direct repeat is a hotspot for large-scale deletion of human mitochondrial DNA. Science 1989, 244, 346–349.
- Lee, H.C.; Pang, C.Y.; Hsu, H.S.; Wei, Y.H. Differential accumulations of 4,977 bp deletion in mitochondrial DNA of various tissues in human ageing. Biochim. Biophys. Acta 1994, 1226, 37–43.
- Mohamed, S.A.; Hanke, T.; Erasmi, A.W.; Bechtel, M.J.; Scharfschwerdt, M.; Meissner, C.; Sievers, H.H.; Gosslau, A. Mitochondrial DNA deletions and the aging heart. Exp. Gerontol. 2006, 41, 508–517.
- Nakano, Y.; Longo-Guess, C.M.; Bergstrom, D.E.; Nauseef, W.M.; Jones, S.M.; Banfi, B. Mutation of the cyba gene encoding p22phox causes vestibular and immune defects in mice. J. Clin. Investig. 2008, 118, 1176–1185.
- Chen, P.; Chai, Y.; Liu, H.; Li, G.; Wang, L.; Yang, T.; Wu, H. Postnatal development of microglia-like cells in mouse cochlea. Neural Plast. 2018, 2018, 1970150.
- Sun, S.; Yu, H.; Yu, H.; Honglin, M.; Ni, W.; Zhang, Y.; Guo, L.; He, Y.; Xue, Z.; Ni, Y.; et al. Inhibition of the activation and recruitment of microglia-like cells protects against neomycin-induced ototoxicity. Mol. Neurobiol. 2015, 51, 252–267.
- Morioka, S.; Sakaguchi, H.; Yamaguchi, T.; Ninoyu, Y.; Mohri, H.; Nakamura, T.; Hisa, Y.; Ogita, K.; Saito, N.; Ueyama, T. Hearing vulnerability after noise exposure in a mouse model of reactive oxygen species overproduction. J. Neurochem. 2018, 146, 459–473.
- Issa, N.; Lachance, G.; Bellmann, K.; Laplante, M.; Stadler, K.; Marette, A. Cytokines promote lipolysis in 3t3-l1 adipocytes through induction of NADPH oxidase 3 expression and superoxide production. J. Lipid Res. 2018, 59, 2321–2328.
- Brault, J.; Vigne, B.; Meunier, M.; Beaumel, S.; Mollin, M.; Park, S.; Stasia, M.J. Nox4 is the main NADPH oxidase involved in the early stages of hematopoietic differentiation from human induced pluripotent stem cells. Free Radic. Biol. Med. 2020, 146, 107–118.
- Chen, G.; Gharib, T.G.; Huang, C.C.; Taylor, J.M.; Misek, D.E.; Kardia, S.L.; Giordano, T.J.; Iannettoni, M.D.; Orringer, M.B.; Hanash, S.M.; et al. Discordant protein and mrna expression in lung adenocarcinomas. Mol. Cell Proteom. 2002, 1, 304–313.
- Pascal, L.E.; True, L.D.; Campbell, D.S.; Deutsch, E.W.; Risk, M.; Coleman, I.M.; Eichner, L.J.; Nelson, P.S.; Liu, A.Y. Correlation of mrna and protein levels: Cell type-specific gene expression of cluster designation antigens in the prostate. BMC Genom. 2008, 9, 246.
- Gluschko, A.; Herb, M.; Wiegmann, K.; Krut, O.; Neiss, W.F.; Utermohlen, O.; Kronke, M.; Schramm, M. The β2 integrin mac-1 induces protective lc3-associated phagocytosis of listeria monocytogenes. Cell Host Microbe 2018, 23, 324–337.e325.
- Rousset, F.; Salmon, P.; Bredl, S.; Cherpin, O.; Coelho, M.; Myburgh, R.; Alessandrini, M.; Perny, M.; Roccio, M.; Speck, R.F.; et al. Optimizing synthetic mirna minigene architecture for efficient mirna hairpin concatenation and multi-target gene knockdown. Mol. Ther. Nucleic Acids 2019, 14, 351–363.
- Zhang, S.; Xing, J.; Gong, Y.; Li, P.; Wang, B.; Xu, L. Downregulation of vdr in benign paroxysmal positional vertigo patients inhibits otolith-associated protein expression levels. Mol. Med. Rep. 2021, 24, 591.
- Froehling, D.A.; Silverstein, M.D.; Mohr, D.N.; Beatty, C.W.; Offord, K.P.; Ballard, D.J. Benign positional vertigo: Incidence and prognosis in a population-based study in olmsted county, minnesota. Mayo Clin. Proc. 1991, 66, 596–601.
- Imai, T.; Higashi-Shingai, K.; Takimoto, Y.; Masumura, C.; Hattori, K.; Inohara, H. New scoring system of an interview for the diagnosis of benign paroxysmal positional vertigo. Acta Oto-Laryngol. 2016, 136, 283–288.
- Fife, T.D.; Iverson, D.J.; Lempert, T.; Furman, J.M.; Baloh, R.W.; Tusa, R.J.; Hain, T.C.; Herdman, S.; Morrow, M.J.; Gronseth, G.S.; et al. Practice parameter: Therapies for benign paroxysmal positional vertigo (an evidence-based review): Report of the quality standards subcommittee of the american academy of neurology. Neurology 2008, 70, 2067–2074.
- Bhattacharyya, N.; Gubbels, S.P.; Schwartz, S.R.; Edlow, J.A.; El-Kashlan, H.; Fife, T.; Holmberg, J.M.; Mahoney, K.; Hollingsworth, D.B.; Roberts, R.; et al. Clinical practice guideline: Benign paroxysmal positional vertigo (update). Otolaryngol.-Head Neck Surg. 2017, 156, S1–S47.
- Lundberg, Y.W.; Xu, Y.; Thiessen, K.D.; Kramer, K.L. Mechanisms of otoconia and otolith development. Dev. Dyn. 2015, 244, 239–253.
- Jang, Y.S.; Hwang, C.H.; Shin, J.Y.; Bae, W.Y.; Kim, L.S. Age-related changes on the morphology of the otoconia. Laryngoscope 2006, 116, 996–1001.
- Baloh, R.W.; Honrubia, V.; Jacobson, K. Benign positional vertigo: Clinical and oculographic features in 240 cases. Neurology 1987, 37, 371–378.
- Nakashima, T.; Pyykko, I.; Arroll, M.A.; Casselbrant, M.L.; Foster, C.A.; Manzoor, N.F.; Megerian, C.A.; Naganawa, S.; Young, Y.H. Meniere’s disease. Nat. Rev. Dis. Primers 2016, 2, 16028.
- Jeong, S.H.; Kim, J.S.; Shin, J.W.; Kim, S.; Lee, H.; Lee, A.Y.; Kim, J.M.; Jo, H.; Song, J.; Ghim, Y. Decreased serum vitamin d in idiopathic benign paroxysmal positional vertigo. J. Neurol. 2013, 260, 832–838.
- Wu, Y.; Gu, C.; Han, W.; Lu, X.; Chen, C.; Fan, Z. Reduction of bone mineral density in native chinese female idiopathic benign paroxysmal positional vertigo patients. Am J Otolaryngol 2018, 39, 31–33.
- Yoda, S.; Cureoglu, S.; Yildirim-Baylan, M.; Morita, N.; Fukushima, H.; Harada, T.; Paparella, M.M. Association between type 1 diabetes mellitus and deposits in the semicircular canals. Otolaryngol. Head Neck Surg. 2011, 145, 458–462.
- Holick, M.F. Resurrection of vitamin d deficiency and rickets. J. Clin. Investig. 2006, 116, 2062–2072.
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin d: Metabolism, molecular mechanism of action, and pleiotropic effects. Physiol. Rev. 2016, 96, 365–408.
- Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery; Society of Otorhinolaryngology Head and Neck Surgery Chinese Medical Association. Guideline of diagnosis and treatment of benign paroxysmal positional vertigo (2017). Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi Chin. J. Otorhinolaryngol. Head Neck Surg. 2017, 52, 173–177.
- Barnes, P.J.; Shapiro, S.D.; Pauwels, R.A. Chronic obstructive pulmonary disease: Molecular and cellular mechanisms. Eur. Respir. J. 2003, 22, 672–688.
- Lokke, A.; Lange, P.; Scharling, H.; Fabricius, P.; Vestbo, J. Developing copd: A 25 year follow up study of the general population. Thorax 2006, 61, 935–939.
- Rangasamy, T.; Cho, C.Y.; Thimmulappa, R.K.; Zhen, L.; Srisuma, S.S.; Kensler, T.W.; Yamamoto, M.; Petrache, I.; Tuder, R.M.; Biswal, S. Genetic ablation of nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J. Clin. Investig. 2004, 114, 1248–1259.
- MacNee, W. Pulmonary and systemic oxidant/antioxidant imbalance in chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005, 2, 50–60.
- Taggart, C.; Cervantes-Laurean, D.; Kim, G.; McElvaney, N.G.; Wehr, N.; Moss, J.; Levine, R.L. Oxidation of either methionine 351 or methionine 358 in alpha 1-antitrypsin causes loss of anti-neutrophil elastase activity. J. Biol. Chem. 2000, 275, 27258–27265.
- Zhang, X.; Shan, P.; Sasidhar, M.; Chupp, G.L.; Flavell, R.A.; Choi, A.M.; Lee, P.J. Reactive oxygen species and extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase mediate hyperoxia-induced cell death in lung epithelium. Am. J. Respir. Cell Mol. Biol. 2003, 28, 305–315.
- Ishii, Y.; Itoh, K.; Morishima, Y.; Kimura, T.; Kiwamoto, T.; Iizuka, T.; Hegab, A.E.; Hosoya, T.; Nomura, A.; Sakamoto, T.; et al. Transcription factor nrf2 plays a pivotal role in protection against elastase-induced pulmonary inflammation and emphysema. J. Immunol. 2005, 175, 6968–6975.
- Pons, J.; Sauleda, J.; Regueiro, V.; Santos, C.; Lopez, M.; Ferrer, J.; Agusti, A.G.; Bengoechea, J.A. Expression of toll-like receptor 2 is up-regulated in monocytes from patients with chronic obstructive pulmonary disease. Respir. Res. 2006, 7, 64.
- Sukkar, M.B.; Xie, S.; Khorasani, N.M.; Kon, O.M.; Stanbridge, R.; Issa, R.; Chung, K.F. Toll-like receptor 2, 3, and 4 expression and function in human airway smooth muscle. J. Allergy Clin. Immunol. 2006, 118, 641–648.
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008, 133, 235–249.
- Chung, K.F.; Marwick, J.A. Molecular mechanisms of oxidative stress in airways and lungs with reference to asthma and chronic obstructive pulmonary disease. Ann. N. Y. Acad. Sci. 2010, 1203, 85–91.
- Pratico, D.; Basili, S.; Vieri, M.; Cordova, C.; Violi, F.; Fitzgerald, G.A. Chronic obstructive pulmonary disease is associated with an increase in urinary levels of isoprostane f2alpha-iii, an index of oxidant stress. Am. J. Respir. Crit. Care Med. 1998, 158, 1709–1714.
- Lius, E.E.; Syafaah, I. Hyperoxia in the management of respiratory failure: A literature review. Ann. Med. Surg. 2022, 81, 104393.
- Lilien, T.A.; van Meenen, D.M.P.; Schultz, M.J.; Bos, L.D.J.; Bem, R.A. Hyperoxia-induced lung injury in acute respiratory distress syndrome: What is its relative impact? Am. J. Physiol. Lung Cell. Mol. Physiol. 2023, 325, L9–L16.
- Minkove, S.; Dhamapurkar, R.; Cui, X.; Li, Y.; Sun, J.; Cooper, D.; Eichacker, P.Q.; Torabi-Parizi, P. Effect of low-to-moderate hyperoxia on lung injury in preclinical animal models: A systematic review and meta-analysis. Intensive Care Med. Exp. 2023, 11, 22.
- Kallet, R.H.; Matthay, M.A. Hyperoxic acute lung injury. Respir. Care 2013, 58, 123–141.
- Zhang, X.; Shan, P.; Qureshi, S.; Homer, R.; Medzhitov, R.; Noble, P.W.; Lee, P.J. Cutting edge: Tlr4 deficiency confers susceptibility to lethal oxidant lung injury. J. Immunol. 2005, 175, 4834–4838.
- Zhang, Y.; Zhang, X.; Shan, P.; Hunt, C.R.; Pandita, T.K.; Lee, P.J. A protective hsp70-tlr4 pathway in lethal oxidant lung injury. J. Immunol. 2013, 191, 1393–1403.
- Hunt, C.R.; Dix, D.J.; Sharma, G.G.; Pandita, R.K.; Gupta, A.; Funk, M.; Pandita, T.K. Genomic instability and enhanced radiosensitivity in hsp70.1- and hsp70.3-deficient mice. Mol. Cell. Biol. 2004, 24, 899–911.
- Fitzgerald, K.A.; Palsson-McDermott, E.M.; Bowie, A.G.; Jefferies, C.A.; Mansell, A.S.; Brady, G.; Brint, E.; Dunne, A.; Gray, P.; Harte, M.T.; et al. Mal (myd88-adapter-like) is required for toll-like receptor-4 signal transduction. Nature 2001, 413, 78–83.
- Rajpoot, S.; Wary, K.K.; Ibbott, R.; Liu, D.; Saqib, U.; Thurston, T.L.M.; Baig, M.S. Tirap in the mechanism of inflammation. Front. Immunol. 2021, 12, 697588.
- Ruwanpura, S.M.; McLeod, L.; Lilja, A.R.; Brooks, G.; Dousha, L.F.; Seow, H.J.; Bozinovski, S.; Vlahos, R.; Hertzog, P.J.; Anderson, G.P.; et al. Non-essential role for tlr2 and its signaling adaptor mal/tirap in preserving normal lung architecture in mice. PLoS ONE 2013, 8, e78095.
- Ruwanpura, S.M.; McLeod, L.; Miller, A.; Jones, J.; Bozinovski, S.; Vlahos, R.; Ernst, M.; Armes, J.; Bardin, P.G.; Anderson, G.P.; et al. Interleukin-6 promotes pulmonary emphysema associated with apoptosis in mice. Am. J. Respir. Cell Mol. Biol. 2011, 45, 720–730.
- Ruwanpura, S.M.; McLeod, L.; Miller, A.; Jones, J.; Vlahos, R.; Ramm, G.; Longano, A.; Bardin, P.G.; Bozinovski, S.; Anderson, G.P.; et al. Deregulated stat3 signaling dissociates pulmonary inflammation from emphysema in gp130 mutant mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L627–L639.
- Hantos, Z.; Adamicza, A.; Janosi, T.Z.; Szabari, M.V.; Tolnai, J.; Suki, B. Lung volumes and respiratory mechanics in elastase-induced emphysema in mice. J. Appl. Physiol. 2008, 105, 1864–1872.
- Yasuoka, H.; Garrett, S.M.; Nguyen, X.X.; Artlett, C.M.; Feghali-Bostwick, C.A. NADPH oxidase-mediated induction of reactive oxygen species and extracellular matrix deposition by insulin-like growth factor binding protein-5. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L644–L655.
- King, T.E., Jr.; Schwarz, M.I.; Brown, K.; Tooze, J.A.; Colby, T.V.; Waldron, J.A., Jr.; Flint, A.; Thurlbeck, W.; Cherniack, R.M. Idiopathic pulmonary fibrosis: Relationship between histopathologic features and mortality. Am. J. Respir. Crit. Care Med. 2001, 164, 1025–1032.
- Navaratnam, V.; Fleming, K.M.; West, J.; Smith, C.J.; Jenkins, R.G.; Fogarty, A.; Hubbard, R.B. The rising incidence of idiopathic pulmonary fibrosis in the UK. Thorax 2011, 66, 462–467.
- Zisman, D.A.; Keane, M.P.; Belperio, J.A.; Strieter, R.M.; Lynch, J.P. Pulmonary fibrosis. In Fibrosis Research: Methods and Protocols; Varga, J., Brenner, D.A., Phan, S.H., Eds.; Humana Press: Totowa, NJ, USA, 2005; pp. 3–44.
- Bocchino, M.; Agnese, S.; Fagone, E.; Svegliati, S.; Grieco, D.; Vancheri, C.; Gabrielli, A.; Sanduzzi, A.; Avvedimento, E.V. Reactive oxygen species are required for maintenance and differentiation of primary lung fibroblasts in idiopathic pulmonary fibrosis. PLoS ONE 2010, 5, e14003.
- Manoury, B.; Nenan, S.; Leclerc, O.; Guenon, I.; Boichot, E.; Planquois, J.M.; Bertrand, C.P.; Lagente, V. The absence of reactive oxygen species production protects mice against bleomycin-induced pulmonary fibrosis. Respir. Res. 2005, 6, 11.
- Murthy, S.; Adamcakova-Dodd, A.; Perry, S.S.; Tephly, L.A.; Keller, R.M.; Metwali, N.; Meyerholz, D.K.; Wang, Y.; Glogauer, M.; Thorne, P.S.; et al. Modulation of reactive oxygen species by rac1 or catalase prevents asbestos-induced pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L846–L855.
- Kinnula, V.L.; Fattman, C.L.; Tan, R.J.; Oury, T.D. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am. J. Respir. Crit. Care Med. 2005, 172, 417–422.
- Korfei, M.; von der Beck, D.; Henneke, I.; Markart, P.; Ruppert, C.; Mahavadi, P.; Ghanim, B.; Klepetko, W.; Fink, L.; Meiners, S.; et al. Comparative proteome analysis of lung tissue from patients with idiopathic pulmonary fibrosis (ipf), non-specific interstitial pneumonia (nsip) and organ donors. J. Proteom. 2013, 85, 109–128.
- Montuschi, P.; Ciabattoni, G.; Paredi, P.; Pantelidis, P.; du Bois, R.M.; Kharitonov, S.A.; Barnes, P.J. 8-isoprostane as a biomarker of oxidative stress in interstitial lung diseases. Am. J. Respir. Crit. Care Med. 1998, 158, 1524–1527.
- Beattie, J.; Allan, G.J.; Lochrie, J.D.; Flint, D.J. Insulin-like growth factor-binding protein-5 (igfbp-5): A critical member of the igf axis. Biochem. J. 2006, 395, 1–19.
- Han, N.; Zhang, F.; Li, G.; Zhang, X.; Lin, X.; Yang, H.; Wang, L.; Cao, Y.; Du, J.; Fan, Z. Local application of igfbp5 protein enhanced periodontal tissue regeneration via increasing the migration, cell proliferation and osteo/dentinogenic differentiation of mesenchymal stem cells in an inflammatory niche. Stem Cell Res. Ther. 2017, 8, 210.
- Tremblay, K.; Lemire, M.; Potvin, C.; Tremblay, A.; Hunninghake, G.M.; Raby, B.A.; Hudson, T.J.; Perez-Iratxeta, C.; Andrade-Navarro, M.A.; Laprise, C. Genes to diseases (g2d) computational method to identify asthma candidate genes. PLoS ONE 2008, 3, e2907.
- Perez-Iratxeta, C.; Wjst, M.; Bork, P.; Andrade, M.A. G2d: A tool for mining genes associated with disease. BMC Genet 2005, 6, 45.
- Perez-Iratxeta, C.; Bork, P.; Andrade-Navarro, M.A. Update of the g2d tool for prioritization of gene candidates to inherited diseases. Nucleic Acids Res. 2007, 35, W212–W216.
- Heyer, E.; Tremblay, M. Variability of the genetic contribution of quebec population founders associated to some deleterious genes. Am. J. Hum. Genet. 1995, 56, 970–978.
- Richter, A.; Rioux, J.D.; Bouchard, J.P.; Mercier, J.; Mathieu, J.; Ge, B.; Poirier, J.; Julien, D.; Gyapay, G.; Weissenbach, J.; et al. Location score and haplotype analyses of the locus for autosomal recessive spastic ataxia of charlevoix-saguenay, in chromosome region 13q11. Am. J. Hum. Genet. 1999, 64, 768–775.
- Yin, C.; Li, K.; Yu, Y.; Huang, H.; Yu, Y.; Wang, Z.; Yan, J.; Pu, Y.; Li, Z.; Li, D.; et al. Genome-wide association study identifies loci and candidate genes for non-idiopathic pulmonary hypertension in eastern chinese han population. BMC Pulm. Med. 2018, 18, 158.
- Barman, S.A.; Chen, F.; Li, X.; Haigh, S.; Stepp, D.W.; Kondrikov, D.; Mahboubi, K.; Bordan, Z.; Traber, P.; Su, Y.; et al. Galectin-3 promotes vascular remodeling and contributes to pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2018, 197, 1488–1492.
- Soubrier, F.; Chung, W.K.; Machado, R.; Grunig, E.; Aldred, M.; Geraci, M.; Loyd, J.E.; Elliott, C.G.; Trembath, R.C.; Newman, J.H.; et al. Genetics and genomics of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2013, 62, D13–D21.
- Cantu, E.; Shah, R.J.; Lin, W.; Daye, Z.J.; Diamond, J.M.; Suzuki, Y.; Ellis, J.H.; Borders, C.F.; Andah, G.A.; Beduhn, B.; et al. Oxidant stress regulatory genetic variation in recipients and donors contributes to risk of primary graft dysfunction after lung transplantation. J. Thorac. Cardiovasc. Surg. 2015, 149, 596–602.
- Charles, E.J.; Kron, I.L. One step closer to the elimination of primary graft dysfunction. J. Thorac. Cardiovasc. Surg. 2015, 149, 602–603.
- Christie, J.D.; Carby, M.; Bag, R.; Corris, P.; Hertz, M.; Weill, D. Report of the ISHLT Working Group on Primary Lung Graft Dysfunction Part II: Definition. A consensus statement of the international society for heart and lung transplantation. J. Heart Lung Transplant 2005, 24, 1454–1459.
- Christie, J.D.; Edwards, L.B.; Kucheryavaya, A.Y.; Aurora, P.; Dobbels, F.; Kirk, R.; Rahmel, A.O.; Stehlik, J.; Hertz, M.I. The registry of the international society for heart and lung transplantation: Twenty-seventh official adult lung and heart-lung transplant report—2010. J. Heart Lung Transplant. 2010, 29, 1104–1118.
- Suzuki, Y.; Cantu, E.; Christie, J.D. Primary graft dysfunction. Semin. Respir. Crit. Care Med. 2013, 34, 305–319.
- de Perrot, M.; Liu, M.; Waddell, T.K.; Keshavjee, S. Ischemia-reperfusion-induced lung injury. Am. J. Respir. Crit. Care Med. 2003, 167, 490–511.
- Williams, A.; Riise, G.C.; Anderson, B.A.; Kjellstrom, C.; Schersten, H.; Kelly, F.J. Compromised antioxidant status and persistent oxidative stress in lung transplant recipients. Free Radic. Res. 1999, 30, 383–393.
- Rega, F.R.; Wuyts, W.A.; Vanaudenaerde, B.M.; Jannis, N.C.; Neyrinck, A.P.; Verleden, G.M.; Lerut, T.E.; Van Raemdonck, D.E. Nebulized n-acetyl cysteine protects the pulmonary graft inside the non-heart-beating donor. J. Heart Lung Transplant. 2005, 24, 1369–1377.
- Kozower, B.D.; Christofidou-Solomidou, M.; Sweitzer, T.D.; Muro, S.; Buerk, D.G.; Solomides, C.C.; Albelda, S.M.; Patterson, G.A.; Muzykantov, V.R. Immunotargeting of catalase to the pulmonary endothelium alleviates oxidative stress and reduces acute lung transplantation injury. Nat. Biotechnol. 2003, 21, 392–398.
- Cho, H.Y.; Reddy, S.P.; Kleeberger, S.R. Nrf2 defends the lung from oxidative stress. Antioxid. Redox Signal. 2006, 8, 76–87.
- Lubos, E.; Mahoney, C.E.; Leopold, J.A.; Zhang, Y.Y.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 modulates lipopolysaccharide-induced adhesion molecule expression in endothelial cells by altering cd14 expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 2525–2532.
- Hamanishi, T.; Furuta, H.; Kato, H.; Doi, A.; Tamai, M.; Shimomura, H.; Sakagashira, S.; Nishi, M.; Sasaki, H.; Sanke, T.; et al. Functional variants in the glutathione peroxidase-1 (gpx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in japanese type 2 diabetic patients. Diabetes 2004, 53, 2455–2460.
- Liu, P.; Shi, L.; Cang, X.; Huang, J.; Wu, X.; Yan, J.; Chen, L.; Cui, S.; Ye, X. Ctbp2 ameliorates palmitate-induced insulin resistance in hepg2 cells through ros mediated jnk pathway. Gen. Comp. Endocrinol. 2017, 247, 66–73.
- Styskal, J.; Van Remmen, H.; Richardson, A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radic. Biol. Med. 2012, 52, 46–58.
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575.
- Henriksen, E.J.; Diamond-Stanic, M.K.; Marchionne, E.M. Oxidative stress and the etiology of insulin resistance and type 2 diabetes. Free Radic. Biol. Med. 2011, 51, 993–999.
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metab. Clin. Exp. 2008, 57, 1071–1077.
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792.
- Elnakish, M.T.; Hassanain, H.H.; Janssen, P.M.; Angelos, M.G.; Khan, M. Emerging role of oxidative stress in metabolic syndrome and cardiovascular diseases: Important role of rac/NADPH oxidase. J. Pathol. 2013, 231, 290–300.
- Zhang, C.H.; Zhou, B.G.; Sheng, J.Q.; Chen, Y.; Cao, Y.Q.; Chen, C. Molecular mechanisms of hepatic insulin resistance in nonalcoholic fatty liver disease and potential treatment strategies. Pharmacol. Res. 2020, 159, 104984.
- Santoleri, D.; Titchenell, P.M. Resolving the paradox of hepatic insulin resistance. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 447–456.
- Boden, G.; Shulman, G.I. Free fatty acids in obesity and type 2 diabetes: Defining their role in the development of insulin resistance and beta-cell dysfunction. Eur. J. Clin. Investig. 2002, 32, 14–23.
- Collaboration, N.C.D.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396.
- Gao, D.; Nong, S.; Huang, X.; Lu, Y.; Zhao, H.; Lin, Y.; Man, Y.; Wang, S.; Yang, J.; Li, J. The effects of palmitate on hepatic insulin resistance are mediated by NADPH oxidase 3-derived reactive oxygen species through jnk and p38mapk pathways. J. Biol. Chem. 2010, 285, 29965–29973.
- Nakamura, S.; Takamura, T.; Matsuzawa-Nagata, N.; Takayama, H.; Misu, H.; Noda, H.; Nabemoto, S.; Kurita, S.; Ota, T.; Ando, H.; et al. Palmitate induces insulin resistance in h4iiec3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009, 284, 14809–14818.
- Montonen, J.; Knekt, P.; Harkanen, T.; Jarvinen, R.; Heliovaara, M.; Aromaa, A.; Reunanen, A. Dietary patterns and the incidence of type 2 diabetes. Am. J. Epidemiol. 2005, 161, 219–227.
- Parks, E.; Yki-Jarvinen, H.; Hawkins, M. Out of the frying pan: Dietary saturated fat influences nonalcoholic fatty liver disease. J. Clin. Investig. 2017, 127, 454–456.
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14.
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the international diabetes federation diabetes atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843.
- Magnusson, I.; Rothman, D.L.; Katz, L.D.; Shulman, R.G.; Shulman, G.I. Increased rate of gluconeogenesis in type ii diabetes mellitus. A 13c nuclear magnetic resonance study. J. Clin. Investig. 1992, 90, 1323–1327.
- Bock, G.; Chittilapilly, E.; Basu, R.; Toffolo, G.; Cobelli, C.; Chandramouli, V.; Landau, B.R.; Rizza, R.A. Contribution of hepatic and extrahepatic insulin resistance to the pathogenesis of impaired fasting glucose: Role of increased rates of gluconeogenesis. Diabetes 2007, 56, 1703–1711.
- Basu, R.; Barosa, C.; Jones, J.; Dube, S.; Carter, R.; Basu, A.; Rizza, R.A. Pathogenesis of prediabetes: Role of the liver in isolated fasting hyperglycemia and combined fasting and postprandial hyperglycemia. J. Clin. Endocrinol. Metab. 2013, 98, E409–E417.
- Funatsu, H.; Yamashita, H. Molecular biology in development and progression of diabetic retinopathy. Nihon Rinsho Jpn. J. Clin. Med. 2002, 60, 162–166.
- Wilkinson, C.P.; Ferris, F.L., 3rd; Klein, R.E.; Lee, P.P.; Agardh, C.D.; Davis, M.; Dills, D.; Kampik, A.; Pararajasegaram, R.; Verdaguer, J.T.; et al. Proposed international clinical diabetic retinopathy and diabetic macular edema disease severity scales. Ophthalmology 2003, 110, 1677–1682.
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948.
- Hirosumi, J.; Tuncman, G.; Chang, L.; Gorgun, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central role for jnk in obesity and insulin resistance. Nature 2002, 420, 333–336.
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857.
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuniga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122.
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7.
- Li, L.; He, Q.; Huang, X.; Man, Y.; Zhou, Y.; Wang, S.; Wang, J.; Li, J. Nox3-derived reactive oxygen species promote tnf-alpha-induced reductions in hepatocyte glycogen levels via a JNK pathway. FEBS Lett. 2010, 584, 995–1000.
- Carnesecchi, S.; Carpentier, J.L.; Foti, M.; Szanto, I. Insulin-induced vascular endothelial growth factor expression is mediated by the NADPH oxidase Nox3. Exp. Cell Res. 2006, 312, 3413–3424.
- Bettaieb, A.; Vazquez Prieto, M.A.; Rodriguez Lanzi, C.; Miatello, R.M.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (-)-epicatechin mitigates high-fructose-associated insulin resistance by modulating redox signaling and endoplasmic reticulum stress. Free Radic. Biol. Med. 2014, 72, 247–256.
- Bettaieb, A.; Cremonini, E.; Kang, H.; Kang, J.; Haj, F.G.; Oteiza, P.I. Anti-inflammatory actions of (-)-epicatechin in the adipose tissue of obese mice. Int. J. Biochem. Cell Biol. 2016, 81, 383–392.
- Cremonini, E.; Wang, Z.; Bettaieb, A.; Adamo, A.M.; Daveri, E.; Mills, D.A.; Kalanetra, K.M.; Haj, F.G.; Karakas, S.; Oteiza, P.I. (-)-epicatechin protects the intestinal barrier from high fat diet-induced permeabilization: Implications for steatosis and insulin resistance. Redox Biol. 2018, 14, 588–599.
- Gupta, A.P.; Syed, A.A.; Garg, R.; Goand, U.K.; Singh, P.; Riyazuddin, M.; Valicherla, G.R.; Husain, A.; Gayen, J.R. Pancreastatin inhibitor psti8 attenuates hyperinsulinemia induced obesity and inflammation mediated insulin resistance via mapk/nox3-jnk pathway. Eur. J. Pharmacol. 2019, 864, 172723.
- Loh, Y.P.; Cheng, Y.; Mahata, S.K.; Corti, A.; Tota, B. Chromogranin a and derived peptides in health and disease. J. Mol. Neurosci. MN 2012, 48, 347–356.
- Hossain, Z.; Valicherla, G.R.; Gupta, A.P.; Syed, A.A.; Riyazuddin, M.; Chandra, S.; Siddiqi, M.I.; Gayen, J.R. Discovery of pancreastatin inhibitor psti8 for the treatment of insulin resistance and diabetes: Studies in rodent models of diabetes mellitus. Sci. Rep. 2018, 8, 8715.
- Sanchez-Margalet, V.; Gonzalez-Yanes, C. Pancreastatin inhibits insulin action in rat adipocytes. Am. J. Physiol. 1998, 275, E1055–E1060.
- Gayen, J.R.; Saberi, M.; Schenk, S.; Biswas, N.; Vaingankar, S.M.; Cheung, W.W.; Najjar, S.M.; O’Connor, D.T.; Bandyopadhyay, G.; Mahata, S.K. A novel pathway of insulin sensitivity in chromogranin a null mice: A crucial role for pancreastatin in glucose homeostasis. J. Biol. Chem. 2009, 284, 28498–28509.
- Tatemoto, K.; Efendic, S.; Mutt, V.; Makk, G.; Feistner, G.J.; Barchas, J.D. Pancreastatin, a novel pancreatic peptide that inhibits insulin secretion. Nature 1986, 324, 476–478.
- Bandyopadhyay, G.K.; Lu, M.; Avolio, E.; Siddiqui, J.A.; Gayen, J.R.; Wollam, J.; Vu, C.U.; Chi, N.W.; O’Connor, D.T.; Mahata, S.K. Pancreastatin-dependent inflammatory signaling mediates obesity-induced insulin resistance. Diabetes 2015, 64, 104–116.
- Broedbaek, K.; Hilsted, L. Chromogranin a as biomarker in diabetes. Biomark. Med. 2016, 10, 1181–1189.
- Allu, P.K.; Chirasani, V.R.; Ghosh, D.; Mani, A.; Bera, A.K.; Maji, S.K.; Senapati, S.; Mullasari, A.S.; Mahapatra, N.R. Naturally occurring variants of the dysglycemic peptide pancreastatin: Differential potencies for multiple cellular functions and structure-function correlation. J. Biol. Chem. 2014, 289, 4455–4469.
- Krawczyk, S.A.; Haller, J.F.; Ferrante, T.; Zoeller, R.A.; Corkey, B.E. Reactive oxygen species facilitate translocation of hormone sensitive lipase to the lipid droplet during lipolysis in human differentiated adipocytes. PLoS ONE 2012, 7, e34904.
- Caturano, A.; D’Angelo, M.; Mormone, A.; Russo, V.; Mollica, M.P.; Salvatore, T.; Galiero, R.; Rinaldi, L.; Vetrano, E.; Marfella, R.; et al. Oxidative stress in type 2 diabetes: Impacts from pathogenesis to lifestyle modifications. Curr. Issues Mol. Biol. 2023, 45, 6651–6666.
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63.
- Rajan, S.; Shankar, K.; Beg, M.; Varshney, S.; Gupta, A.; Srivastava, A.; Kumar, D.; Mishra, R.K.; Hussain, Z.; Gayen, J.R.; et al. Chronic hyperinsulinemia reduces insulin sensitivity and metabolic functions of brown adipocyte. J. Endocrinol. 2016, 230, 275–290.
- Garg, R.; Gupta, A.P.; Katekar, R.; Verma, S.; Goand, U.K.; Dadge, S.; Gayen, J.R. Pancreastatin inhibitor psti8 prevents free fatty acid-induced oxidative stress and insulin resistance by modulating jnk pathway: In vitro and in vivo findings. Life Sci. 2022, 289, 120221.
- Mahadev, K.; Motoshima, H.; Wu, X.; Ruddy, J.M.; Arnold, R.S.; Cheng, G.; Lambeth, J.D.; Goldstein, B.J. The NAD(P)H oxidase homolog nox4 modulates insulin-stimulated generation of H2O2 and plays an integral role in insulin signal transduction. Mol. Cell. Biol. 2004, 24, 1844–1854.
- Mouche, S.; Mkaddem, S.B.; Wang, W.; Katic, M.; Tseng, Y.H.; Carnesecchi, S.; Steger, K.; Foti, M.; Meier, C.A.; Muzzin, P.; et al. Reduced expression of the NADPH oxidase nox4 is a hallmark of adipocyte differentiation. Biochim. Biophys. Acta 2007, 1773, 1015–1027.
- Schroder, K.; Wandzioch, K.; Helmcke, I.; Brandes, R.P. Nox4 acts as a switch between differentiation and proliferation in preadipocytes. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 239–245.
- Malik, S.A.; Acharya, J.D.; Mehendale, N.K.; Kamat, S.S.; Ghaskadbi, S.S. Pterostilbene reverses palmitic acid mediated insulin resistance in hepg2 cells by reducing oxidative stress and triglyceride accumulation. Free Radic. Res. 2019, 53, 815–827.
- Remsberg, C.M.; Yanez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. PTR 2008, 22, 169–179.
- Acharya, J.D.; Ghaskadbi, S.S. Protective effect of pterostilbene against free radical mediated oxidative damage. BMC Complement. Altern. Med. 2013, 13, 238.
- Satheesh, M.A.; Pari, L. Effect of pterostilbene on lipids and lipid profiles in streptozotocin-nicotinamide induced type 2 diabetes mellitus. J. Appl. Biomed. 2008, 6, 31–37.
- Elango, B.; Dornadula, S.; Paulmurugan, R.; Ramkumar, K.M. Pterostilbene ameliorates streptozotocin-induced diabetes through enhancing antioxidant signaling pathways mediated by nrf2. Chem. Res. Toxicol. 2016, 29, 47–57.
- Gomez-Zorita, S.; Fernandez-Quintela, A.; Aguirre, L.; Macarulla, M.T.; Rimando, A.M.; Portillo, M.P. Pterostilbene improves glycaemic control in rats fed an obesogenic diet: Involvement of skeletal muscle and liver. Food Funct. 2015, 6, 1968–1976.
- Pihlajaniemi, T.; Myllyla, R.; Kivirikko, K.I.; Tryggvason, K. Effects of streptozotocin diabetes, glucose, and insulin on the metabolism of type iv collagen and proteoglycan in murine basement membrane-forming ehs tumor tissue. J. Biol. Chem. 1982, 257, 14914–14920.
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620.
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ros) and wound healing: The functional role of ros and emerging ros-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96.
- Kim, Y.S.; Lee, H.Y.; Jang, J.Y.; Lee, H.R.; Shin, Y.S.; Kim, C.H. Redox treatment ameliorates diabetes mellitus-induced skin flap necrosis via inhibiting apoptosis and promoting neoangiogenesis. Exp. Biol. Med. 2021, 246, 718–728.
- Toyoda, K.; Fujii, K.; Kamouchi, M.; Nakane, H.; Arihiro, S.; Okada, Y.; Ibayashi, S.; Iida, M. Free radical scavenger, edaravone, in stroke with internal carotid artery occlusion. J. Neurol. Sci. 2004, 221, 11–17.
- Liu, J.; Zhang, D.; Luo, W.; Yu, J.; Li, J.; Yu, Y.; Zhang, X.; Chen, J.; Wu, X.R.; Huang, C. E3 ligase activity of xiap ring domain is required for xiap-mediated cancer cell migration, but not for its rhogdi binding activity. PLoS ONE 2012, 7, e35682.
- Nguyen, T.; Lau, D.C. The obesity epidemic and its impact on hypertension. Can. J. Cardiol. 2012, 28, 326–333.
- Franklin, S.S.; Gustin, W.t.; Wong, N.D.; Larson, M.G.; Weber, M.A.; Kannel, W.B.; Levy, D. Hemodynamic patterns of age-related changes in blood pressure. The framingham heart study. Circulation 1997, 96, 308–315.
- Pravenec, M.; Wallace, C.; Aitman, T.J.; Kurtz, T.W. Gene expression profiling in hypertension research: A critical perspective. Hypertension 2003, 41, 3–8.
- Radkowski, P.; Wator, G.; Skupien, J.; Bogdali, A.; Wolkow, P. Analysis of gene expression to predict dynamics of future hypertension incidence in type 2 diabetic patients. BMC Proc. 2016, 10, 113–117.
- Chen, G.; Adeyemo, A.A.; Zhou, J.; Chen, Y.; Doumatey, A.; Lashley, K.; Huang, H.; Amoah, A.; Agyenim-Boateng, K.; Eghan, B.A., Jr.; et al. A genome-wide search for linkage to renal function phenotypes in west africans with type 2 diabetes. Am. J. Kidney Dis. 2007, 49, 394–400.
- Kwak, S.H.; Hernandez-Cancela, R.B.; DiCorpo, D.A.; Condon, D.E.; Merino, J.; Wu, P.; Brody, J.A.; Yao, J.; Guo, X.; Ahmadizar, F.; et al. Time-to-event genome-wide association study for incident cardiovascular disease in people with type 2 diabetes mellitus. medRxiv 2023.
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645.
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415.
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761.
- Talior, I.; Yarkoni, M.; Bashan, N.; Eldar-Finkelman, H. Increased glucose uptake promotes oxidative stress and pkc-delta activation in adipocytes of obese, insulin-resistant mice. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E295–E302.
- Wajngarten, M.; Silva, G.S. Hypertension and stroke: Update on treatment. Eur. Cardiol. 2019, 14, 111–115.
- Johansson, B.B. Hypertension mechanisms causing stroke. Clin. Exp. Pharmacol. Physiol. 1999, 26, 563–565.
- Neaton, J.D.; Blackburn, H.; Jacobs, D.; Kuller, L.; Lee, D.J.; Sherwin, R.; Shih, J.; Stamler, J.; Wentworth, D. Serum cholesterol level and mortality findings for men screened in the multiple risk factor intervention trial. Multiple risk factor intervention trial research group. Arch. Intern. Med. 1992, 152, 1490–1500.
- Peeling, J.; Yan, H.J.; Chen, S.G.; Campbell, M.; Del Bigio, M.R. Protective effects of free radical inhibitors in intracerebral hemorrhage in rat. Brain Res. 1998, 795, 63–70.
- Aronowski, J.; Zhao, X. Molecular pathophysiology of cerebral hemorrhage: Secondary brain injury. Stroke 2011, 42, 1781–1786.
- Michihara, A.; Oda, A.; Mido, M. High expression levels of NADPH oxidase 3 in the cerebrum of ten-week-old stroke-prone spontaneously hypertensive rats. Biol. Pharm. Bull. 2016, 39, 252–258.
- Okamoto, K.; Yamamoto, K.; Morita, N.; Ohta, Y.; Chikugo, T.; Higashizawa, T.; Suzuki, T. Establishment and use of the m strain of stroke-prone spontaneously hypertensive rat. J. Hypertension. Suppl. 1986, 4, S21–S24.
- Iritani, N.; Fukuda, E.; Nara, Y.; Yamori, Y. Lipid metabolism in spontaneously hypertensive rats (shr). Atherosclerosis 1977, 28, 217–222.
- Tanito, M.; Nakamura, H.; Kwon, Y.W.; Teratani, A.; Masutani, H.; Shioji, K.; Kishimoto, C.; Ohira, A.; Horie, R.; Yodoi, J. Enhanced oxidative stress and impaired thioredoxin expression in spontaneously hypertensive rats. Antioxid. Redox Signal. 2004, 6, 89–97.
- Michihara, A.; Shimatani, M.; Anraku, M.; Tomida, H.; Akasaki, K. High levels of oxidative stress exist in the brain than serum or kidneys in stroke-prone spontaneously hypertensive rats at ten weeks of age. Biol. Pharm. Bull. 2010, 33, 518–521.
- Loft, S.; Vistisen, K.; Ewertz, M.; Tjonneland, A.; Overvad, K.; Poulsen, H.E. Oxidative DNA damage estimated by 8-hydroxydeoxyguanosine excretion in humans: Influence of smoking, gender and body mass index. Carcinogenesis 1992, 13, 2241–2247.
- Nakamura, T.; Yamamoto, E.; Kataoka, K.; Yamashita, T.; Tokutomi, Y.; Dong, Y.F.; Matsuba, S.; Ogawa, H.; Kim-Mitsuyama, S. Pioglitazone exerts protective effects against stroke in stroke-prone spontaneously hypertensive rats, independently of blood pressure. Stroke 2007, 38, 3016–3022.
- Ahmarani, L.; Avedanian, L.; Al-Khoury, J.; Perreault, C.; Jacques, D.; Bkaily, G. Whole-cell and nuclear NADPH oxidases levels and distribution in human endocardial endothelial, vascular smooth muscle, and vascular endothelial cells. Can. J. Physiol. Pharmacol. 2013, 91, 71–79.
- Lassegue, B.; San Martin, A.; Griendling, K.K. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res. 2012, 110, 1364–1390.
- Nediani, C.; Borchi, E.; Giordano, C.; Baruzzo, S.; Ponziani, V.; Sebastiani, M.; Nassi, P.; Mugelli, A.; d’Amati, G.; Cerbai, E. NADPH oxidase-dependent redox signaling in human heart failure: Relationship between the left and right ventricle. J. Mol. Cell Cardiol. 2007, 42, 826–834.
- Heymes, C.; Bendall, J.K.; Ratajczak, P.; Cave, A.C.; Samuel, J.L.; Hasenfuss, G.; Shah, A.M. Increased myocardial NADPH oxidase activity in human heart failure. J. Am. Coll. Cardiol. 2003, 41, 2164–2171.
- Borchi, E.; Bargelli, V.; Stillitano, F.; Giordano, C.; Sebastiani, M.; Nassi, P.A.; d’Amati, G.; Cerbai, E.; Nediani, C. Enhanced ros production by NADPH oxidase is correlated to changes in antioxidant enzyme activity in human heart failure. Biochim. Biophys. Acta 2010, 1802, 331–338.
- Maack, C.; Kartes, T.; Kilter, H.; Schafers, H.J.; Nickenig, G.; Bohm, M.; Laufs, U. Oxygen free radical release in human failing myocardium is associated with increased activity of rac1-gtpase and represents a target for statin treatment. Circulation 2003, 108, 1567–1574.
- Zhang, M.; Perino, A.; Ghigo, A.; Hirsch, E.; Shah, A.M. NADPH oxidases in heart failure: Poachers or gamekeepers? Antioxid. Redox Signal. 2013, 18, 1024–1041.
- Li, J.; Stouffs, M.; Serrander, L.; Banfi, B.; Bettiol, E.; Charnay, Y.; Steger, K.; Krause, K.H.; Jaconi, M.E. The NADPH oxidase nox4 drives cardiac differentiation: Role in regulating cardiac transcription factors and map kinase activation. Mol. Biol. Cell 2006, 17, 3978–3988.
- Fenyo, I.M.; Florea, I.C.; Raicu, M.; Manea, A. Tyrphostin ag490 reduces napdh oxidase activity and expression in the aorta of hypercholesterolemic apolipoprotein e-deficient mice. Vasc. Pharmacol. 2011, 54, 100–106.
- Sirker, A.; Murdoch, C.E.; Protti, A.; Sawyer, G.J.; Santos, C.X.; Martin, D.; Zhang, X.; Brewer, A.C.; Zhang, M.; Shah, A.M. Cell-specific effects of nox2 on the acute and chronic response to myocardial infarction. J. Mol. Cell Cardiol. 2016, 98, 11–17.
- Bell, R.M.; Cave, A.C.; Johar, S.; Hearse, D.J.; Shah, A.M.; Shattock, M.J. Pivotal role of nox-2-containing NADPH oxidase in early ischemic preconditioning. FASEB J. 2005, 19, 2037–2039.
- Fukui, H.; Moraes, C.T. The mitochondrial impairment, oxidative stress and neurodegeneration connection: Reality or just an attractive hypothesis? Trends Neurosci. 2008, 31, 251–256.
- Wind, S.; Beuerlein, K.; Armitage, M.E.; Taye, A.; Kumar, A.H.; Janowitz, D.; Neff, C.; Shah, A.M.; Wingler, K.; Schmidt, H.H. Oxidative stress and endothelial dysfunction in aortas of aged spontaneously hypertensive rats by nox1/2 is reversed by NADPH oxidase inhibition. Hypertension 2010, 56, 490–497.
- Bkaily, G.; Najibeddine, W.; Jacques, D. Increase of NADPH oxidase 3 in heart failure of hereditary cardiomyopathy (1). Can. J. Physiol. Pharmacol. 2019, 97, 902–908.
- Dhalla, A.K.; Hill, M.F.; Singal, P.K. Role of oxidative stress in transition of hypertrophy to heart failure. J. Am. Coll. Cardiol. 1996, 28, 506–514.
- Wu, R.; Yin, D.; Sadekova, N.; Deschepper, C.F.; de Champlain, J.; Girouard, H. Protective effects of aspirin from cardiac hypertrophy and oxidative stress in cardiomyopathic hamsters. Oxidative Med. Cell. Longev. 2012, 2012, 761710.
- Mollnau, H.; Oelze, M.; August, M.; Wendt, M.; Daiber, A.; Schulz, E.; Baldus, S.; Kleschyov, A.L.; Materne, A.; Wenzel, P.; et al. Mechanisms of increased vascular superoxide production in an experimental model of idiopathic dilated cardiomyopathy. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2554–2559.
- Jacques, D.; Bkaily, G. Endocardial endothelial cell hypertrophy takes place during the development of hereditary cardiomyopathy. Mol. Cell Biochem. 2019, 453, 157–161.
- Dhalla, N.S.; Elmoselhi, A.B.; Hata, T.; Makino, N. Status of myocardial antioxidants in ischemia-reperfusion injury. Cardiovasc. Res. 2000, 47, 446–456.
- Bolli, R.; Jeroudi, M.O.; Patel, B.S.; Aruoma, O.I.; Halliwell, B.; Lai, E.K.; McCay, P.B. Marked reduction of free radical generation and contractile dysfunction by antioxidant therapy begun at the time of reperfusion. Evidence that myocardial “stunning” is a manifestation of reperfusion injury. Circ. Res. 1989, 65, 607–622.
- Zhao, Z.Q.; Nakamura, M.; Wang, N.P.; Wilcox, J.N.; Shearer, S.; Ronson, R.S.; Guyton, R.A.; Vinten-Johansen, J. Reperfusion induces myocardial apoptotic cell death. Cardiovasc. Res. 2000, 45, 651–660.
- Frangogiannis, N.G.; Smith, C.W.; Entman, M.L. The inflammatory response in myocardial infarction. Cardiovasc. Res. 2002, 53, 31–47.
- Kloner, R.A.; Jennings, R.B. Consequences of brief ischemia: Stunning, preconditioning, and their clinical implications: Part 2. Circulation 2001, 104, 3158–3167.
- Hayashidani, S.; Tsutsui, H.; Shiomi, T.; Ikeuchi, M.; Matsusaka, H.; Suematsu, N.; Wen, J.; Egashira, K.; Takeshita, A. Anti-monocyte chemoattractant protein-1 gene therapy attenuates left ventricular remodeling and failure after experimental myocardial infarction. Circulation 2003, 108, 2134–2140.
- Kaikita, K.; Hayasaki, T.; Okuma, T.; Kuziel, W.A.; Ogawa, H.; Takeya, M. Targeted deletion of cc chemokine receptor 2 attenuates left ventricular remodeling after experimental myocardial infarction. Am. J. Pathol. 2004, 165, 439–447.
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. Ccl2/monocyte chemoattractant protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ. Res. 2005, 96, 881–889.
- Morimoto, H.; Hirose, M.; Takahashi, M.; Kawaguchi, M.; Ise, H.; Kolattukudy, P.E.; Yamada, M.; Ikeda, U. Mcp-1 induces cardioprotection against ischaemia/reperfusion injury: Role of reactive oxygen species. Cardiovasc. Res. 2008, 78, 554–562.
- Kolattukudy, P.E.; Quach, T.; Bergese, S.; Breckenridge, S.; Hensley, J.; Altschuld, R.; Gordillo, G.; Klenotic, S.; Orosz, C.; Parker-Thornburg, J. Myocarditis induced by targeted expression of the mcp-1 gene in murine cardiac muscle. Am. J. Pathol. 1998, 152, 101–111.
- Vats, S.; Sundquist, K.; Sundquist, J.; Zhang, N.; Wang, X.; Acosta, S.; Gottsater, A.; Memon, A.A. Oxidative stress-related genetic variation and antioxidant vitamin intake in intact and ruptured abdominal aortic aneurysm: A swedish population-based retrospective cohort study. Eur. J. Prev. Cardiol. 2024, 31, 61–74.
- Manjer, J.; Elmstahl, S.; Janzon, L.; Berglund, G. Invitation to a population-based cohort study: Differences between subjects recruited using various strategies. Scand. J. Public Health 2002, 30, 103–112.
- Wanhainen, A.; Verzini, F.; Van Herzeele, I.; Allaire, E.; Bown, M.; Cohnert, T.; Dick, F.; van Herwaarden, J.; Karkos, C.; Koelemay, M.; et al. Editor’s choice—European society for vascular surgery (ESVS) 2019 clinical practice guidelines on the management of abdominal aorto-iliac artery aneurysms. Eur. J. Vasc. Endovasc. Surg. 2019, 57, 8–93.
- Howard, D.P.; Banerjee, A.; Fairhead, J.F.; Handa, A.; Silver, L.E.; Rothwell, P.M.; Oxford Vascular, S. Age-specific incidence, risk factors and outcome of acute abdominal aortic aneurysms in a defined population. Br. J. Surg. 2015, 102, 907–915.
- Reimerink, J.J.; van der Laan, M.J.; Koelemay, M.J.; Balm, R.; Legemate, D.A. Systematic review and meta-analysis of population-based mortality from ruptured abdominal aortic aneurysm. Br. J. Surg. 2013, 100, 1405–1413.
- Song, P.; He, Y.; Adeloye, D.; Zhu, Y.; Ye, X.; Yi, Q.; Rahimi, K.; Rudan, I.; Global Health Epidemiology Research Group. The global and regional prevalence of abdominal aortic aneurysms: A systematic review and modeling analysis. Ann. Surg. 2023, 277, 912–919.
- Sweeting, M.J.; Thompson, S.G.; Brown, L.C.; Powell, J.T.; Collaborators, R. Meta-analysis of individual patient data to examine factors affecting growth and rupture of small abdominal aortic aneurysms. Br. J. Surg. 2012, 99, 655–665.
- Guzik, B.; Sagan, A.; Ludew, D.; Mrowiecki, W.; Chwala, M.; Bujak-Gizycka, B.; Filip, G.; Grudzien, G.; Kapelak, B.; Zmudka, K.; et al. Mechanisms of oxidative stress in human aortic aneurysms--association with clinical risk factors for atherosclerosis and disease severity. Int. J. Cardiol. 2013, 168, 2389–2396.
- McCormick, M.L.; Gavrila, D.; Weintraub, N.L. Role of oxidative stress in the pathogenesis of abdominal aortic aneurysms. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 461–469.
- Kim, H.W.; Blomkalns, A.L.; Ogbi, M.; Thomas, M.; Gavrila, D.; Neltner, B.S.; Cassis, L.A.; Thompson, R.W.; Weiss, R.M.; Lindower, P.D.; et al. Role of myeloperoxidase in abdominal aortic aneurysm formation: Mitigation by taurine. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H1168–H1179.
- Ludvigsson, J.F.; Andersson, E.; Ekbom, A.; Feychting, M.; Kim, J.L.; Reuterwall, C.; Heurgren, M.; Olausson, P.O. External review and validation of the swedish national inpatient register. BMC Public Health 2011, 11, 450.
- Caramori, M.L.; Fioretto, P.; Mauer, M. Low glomerular filtration rate in normoalbuminuric type 1 diabetic patients: An indicator of more advanced glomerular lesions. Diabetes 2003, 52, 1036–1040.
- Cockcroft, D.W.; Gault, M.H. Prediction of creatinine clearance from serum creatinine. Nephron 1976, 16, 31–41.
- Hunt, S.C.; Hasstedt, S.J.; Coon, H.; Camp, N.J.; Cawthon, R.M.; Wu, L.L.; Hopkins, P.N. Linkage of creatinine clearance to chromosome 10 in utah pedigrees replicates a locus for end-stage renal disease in humans and renal failure in the fawn-hooded rat. Kidney Int. 2002, 62, 1143–1148.
- Ye, S.; Zhong, H.; Yanamadala, S.; Campese, V.M. Oxidative stress mediates the stimulation of sympathetic nerve activity in the phenol renal injury model of hypertension. Hypertension 2006, 48, 309–315.
- Suh, Y.A.; Arnold, R.S.; Lassegue, B.; Shi, J.; Xu, X.; Sorescu, D.; Chung, A.B.; Griendling, K.K.; Lambeth, J.D. Cell transformation by the superoxide-generating oxidase Mox1. Nature 1999, 401, 79–82.
- Rokutan, K.; Kawahara, T.; Kuwano, Y.; Tominaga, K.; Nishida, K.; Teshima-Kondo, S. Nox enzymes and oxidative stress in the immunopathology of the gastrointestinal tract. Semin. Immunopathol. 2008, 30, 315–327.
- Banfi, B.; Maturana, A.; Jaconi, S.; Arnaudeau, S.; Laforge, T.; Sinha, B.; Ligeti, E.; Demaurex, N.; Krause, K.H. A mammalian h+ channel generated through alternative splicing of the NADPH oxidase homolog noh-1. Science 2000, 287, 138–142.
- Teshima, S.; Kutsumi, H.; Kawahara, T.; Kishi, K.; Rokutan, K. Regulation of growth and apoptosis of cultured guinea pig gastric mucosal cells by mitogenic oxidase 1. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G1169–G1176.
- Salles, N.; Szanto, I.; Herrmann, F.; Armenian, B.; Stumm, M.; Stauffer, E.; Michel, J.P.; Krause, K.H. Expression of mrna for ros-generating NADPH oxidases in the aging stomach. Exp. Gerontol. 2005, 40, 353–357.
- Augusto, A.C.; Miguel, F.; Mendonca, S.; Pedrazzoli, J., Jr.; Gurgueira, S.A. Oxidative stress expression status associated to Helicobacter pylori virulence in gastric diseases. Clin. Biochem. 2007, 40, 615–622.
- Geiszt, M.; Witta, J.; Baffi, J.; Lekstrom, K.; Leto, T.L. Dual oxidases represent novel hydrogen peroxide sources supporting mucosal surface host defense. FASEB J. 2003, 17, 1502–1504.
- Dupuy, C.; Pomerance, M.; Ohayon, R.; Noel-Hudson, M.S.; Deme, D.; Chaaraoui, M.; Francon, J.; Virion, A. Thyroid oxidase (thox2) gene expression in the rat thyroid cell line frtl-5. Biochem. Biophys. Res. Commun. 2000, 277, 287–292.
- El Hassani, R.A.; Benfares, N.; Caillou, B.; Talbot, M.; Sabourin, J.C.; Belotte, V.; Morand, S.; Gnidehou, S.; Agnandji, D.; Ohayon, R.; et al. Dual oxidase2 is expressed all along the digestive tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G933–G942.
- Plantinga, T.S.; Arts, P.; Knarren, G.H.; Mulder, A.H.; Wakelkamp, I.M.; Hermus, A.R.; Joosten, L.A.; Netea, M.G.; Bisschop, P.H.; de Herder, W.W.; et al. Rare nox3 variants confer susceptibility to agranulocytosis during thyrostatic treatment of graves’ disease. Clin. Pharmacol. Ther. 2017, 102, 1017–1024.
- Yang, J.; Zhong, J.; Xiao, X.H.; Zhou, L.Z.; Chen, Y.J.; Liu, J.H.; Cao, R.X.; Wen, G.B. The relationship between bone marrow characteristics and the clinical prognosis of antithyroid drug-induced agranulocytosis. Endocr. J. 2013, 60, 185–189.
- Ishimaru, N.; Ohnishi, H.; Nishiuma, T.; Doukuni, R.; Umezawa, K.; Oozone, S.; Kuramoto, E.; Yoshimura, S.; Kinami, S. Antithyroid drug-induced agranulocytosis complicated by pneumococcal sepsis and upper airway obstruction. Intern. Med. 2013, 52, 2355–2359.
- Kubota, Y.; Toh Yoon, E.W. Severe drug-induced agranulocytosis successfully treated with recombinant human granulocyte colony-stimulating factor. Case Rep. Med. 2018, 2018, 8439791.
- Andrès, E.; Maloisel, F. Antibiotic-induced agranulocytosis: A monocentric study of 21 cases. Arch. Intern. Med. 2001, 161, 2619.
- di Fonzo, H.; Villegas Gutsch, M.; Castroagudin, A.; Cabrera, M.V.; Mazzei, M.E.; Rueda, D. Agranulocytosis induced by vancomycin. Case report and literature review. Am. J. Case Rep. 2018, 19, 1053–1056.
- Weissel, M. Propylthiouracil: Clinical overview of its efficacy and its side effects more than 50 years after the introduction of its use in thyrostatic treatment. Exp. Clin. Endocrinol. Diabetes 2010, 118, 101–104.
- Sato, S.; Noh, J.Y.; Sato, S.; Suzuki, M.; Yasuda, S.; Matsumoto, M.; Kunii, Y.; Mukasa, K.; Sugino, K.; Ito, K.; et al. Comparison of efficacy and adverse effects between methimazole 15 mg+inorganic iodine 38 mg/day and methimazole 30 mg/day as initial therapy for graves’ disease patients with moderate to severe hyperthyroidism. Thyroid 2015, 25, 43–50.
- Taylor, P.N.; Vaidya, B. Side effects of anti-thyroid drugs and their impact on the choice of treatment for thyrotoxicosis in pregnancy. Eur. Thyroid J. 2012, 1, 176–185.
- Andersohn, F.; Konzen, C.; Garbe, E. Systematic review: Agranulocytosis induced by nonchemotherapy drugs. Ann. Intern. Med. 2007, 146, 657–665.
- Takata, K.; Kubota, S.; Fukata, S.; Kudo, T.; Nishihara, E.; Ito, M.; Amino, N.; Miyauchi, A. Methimazole-induced agranulocytosis in patients with graves’ disease is more frequent with an initial dose of 30 mg daily than with 15 mg daily. Thyroid 2009, 19, 559–563.
- Chu, X.; Pan, C.M.; Zhao, S.X.; Liang, J.; Gao, G.Q.; Zhang, X.M.; Yuan, G.Y.; Li, C.G.; Xue, L.Q.; Shen, M.; et al. A genome-wide association study identifies two new risk loci for graves’ disease. Nat. Genet. 2011, 43, 897–901.
- Liu, W.; Wang, H.N.; Gu, Z.H.; Yang, S.Y.; Ye, X.P.; Pan, C.M.; Zhao, S.X.; Xue, L.Q.; Xie, H.J.; Yu, S.S.; et al. Identification of bach2 as a susceptibility gene for graves’ disease in the chinese han population based on a three-stage genome-wide association study. Hum. Genet. 2014, 133, 661–671.
- Roderique, J.D.; Josef, C.S.; Feldman, M.J.; Spiess, B.D. A modern literature review of carbon monoxide poisoning theories, therapies, and potential targets for therapy advancement. Toxicology 2015, 334, 45–58.
- Varma, D.R.; Mulay, S.; Chemtob, S. Chapter 20—Carbon monoxide: From public health risk to painless killer. In Handbook of Toxicology of Chemical Warfare Agents; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 271–292.
- Ginsberg, M.D. Delayed neurological deterioration following hypoxia. Adv. Neurol. 1979, 26, 21–44.
- Hara, S.; Mizukami, H.; Kurosaki, K.; Kuriiwa, F.; Mukai, T. Existence of a threshold for hydroxyl radical generation independent of hypoxia in rat striatum during carbon monoxide poisoning. Arch. Toxicol. 2011, 85, 1091–1099.
- Sun, Q.; Cai, J.; Zhou, J.; Tao, H.; Zhang, J.H.; Zhang, W.; Sun, X.J. Hydrogen-rich saline reduces delayed neurologic sequelae in experimental carbon monoxide toxicity. Crit. Care Med. 2011, 39, 765–769.
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694.
- Piantadosi, C.A.; Zhang, J.; Demchenko, I.T. Production of hydroxyl radical in the hippocampus after co hypoxia or hypoxic hypoxia in the rat. Free Radic. Biol. Med. 1997, 22, 725–732.
- Hara, S.; Kobayashi, M.; Kuriiwa, F.; Mukai, T.; Mizukami, H. Different mechanisms of hydroxyl radical production susceptible to purine p2 receptor antagonists between carbon monoxide poisoning and exogenous atp in rat striatum. Free Radic. Res. 2014, 48, 1322–1333.
- Hara, S.; Kobayash, M.; Kuriiwa, F.; Kurosaki, K.; Mizukami, H. Gene expression in rat striatum following carbon monoxide poisoning. Genom Data 2017, 12, 74–75.
- Hara, S.; Kobayashi, M.; Kuriiwa, F.; Ikematsu, K.; Mizukami, H. Hydroxyl radical production via NADPH oxidase in rat striatum due to carbon monoxide poisoning. Toxicology 2018, 394, 63–71.
- Thom, S.R.; Ohnishi, S.T.; Ischiropoulos, H. Nitric oxide released by platelets inhibits neutrophil b2 integrin function following acute carbon monoxide poisoning. Toxicol. Appl. Pharmacol. 1994, 128, 105–110.
- Nabeshima, T.; Katoh, A.; Ishimaru, H.; Yoneda, Y.; Ogita, K.; Murase, K.; Ohtsuka, H.; Inari, K.; Fukuta, T.; Kameyama, T. Carbon monoxide-induced delayed amnesia, delayed neuronal death and change in acetylcholine concentration in mice. J. Pharmacol. Exp. Ther. 1991, 256, 378–384.
- Fujita, K.; Seike, T.; Yutsudo, N.; Ohno, M.; Yamada, H.; Yamaguchi, H.; Sakumi, K.; Yamakawa, Y.; Kido, M.A.; Takaki, A.; et al. Hydrogen in drinking water reduces dopaminergic neuronal loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of parkinson’s disease. PLoS ONE 2009, 4, e7247.
- Mikkola, L.; Holopainen, S.; Pessa-Morikawa, T.; Lappalainen, A.K.; Hytonen, M.K.; Lohi, H.; Iivanainen, A. Genetic dissection of canine hip dysplasia phenotypes and osteoarthritis reveals three novel loci. BMC Genom. 2019, 20, 1027.
- Skurkova, L.; Hluchy, M.; Lackova, M.; Mihalova, M.; Ledecky, V. Relation of the norberg angle and position of the femoral head centre to the dorsal acetabular edge in evaluation of canine hip dysplasia. Vet. Comp. Orthop. Traumatol. 2010, 23, 433–438.
- Broeckx, B.J.G.; Vezzoni, A.; Bogaerts, E.; Bertal, M.; Bosmans, T.; Stock, E.; Deforce, D.; Peelman, L.; Saunders, J.H. Comparison of three methods to quantify laxity in the canine hip joint. Vet. Comp. Orthop. Traumatol. V.C.O.T 2018, 31, 23–29.
- Sanchez-Molano, E.; Woolliams, J.A.; Pong-Wong, R.; Clements, D.N.; Blott, S.C.; Wiener, P. Quantitative trait loci mapping for canine hip dysplasia and its related traits in uk labrador retrievers. BMC Genom. 2014, 15, 833.
- Lavrijsen, I.C.; Leegwater, P.A.; Martin, A.J.; Harris, S.J.; Tryfonidou, M.A.; Heuven, H.C.; Hazewinkel, H.A. Genome wide analysis indicates genes for basement membrane and cartilage matrix proteins as candidates for hip dysplasia in labrador retrievers. PLoS ONE 2014, 9, e87735.
- Fels, L.; Distl, O. Identification and validation of quantitative trait loci (qtl) for canine hip dysplasia (chd) in german shepherd dogs. PLoS ONE 2014, 9, e96618.
- Pfahler, S.; Distl, O. Identification of quantitative trait loci (qtl) for canine hip dysplasia and canine elbow dysplasia in bernese mountain dogs. PLoS ONE 2012, 7, e49782.
- Zhou, Z.; Sheng, X.; Zhang, Z.; Zhao, K.; Zhu, L.; Guo, G.; Friedenberg, S.G.; Hunter, L.S.; Vandenberg-Foels, W.S.; Hornbuckle, W.E.; et al. Differential genetic regulation of canine hip dysplasia and osteoarthritis. PLoS ONE 2010, 5, e13219.
- Carlstrom, K.E.; Ewing, E.; Granqvist, M.; Gyllenberg, A.; Aeinehband, S.; Enoksson, S.L.; Checa, A.; Badam, T.V.S.; Huang, J.; Gomez-Cabrero, D.; et al. Therapeutic efficacy of dimethyl fumarate in relapsing-remitting multiple sclerosis associates with ros pathway in monocytes. Nat. Commun. 2019, 10, 3081.
- Gopal, S.; Mikulskis, A.; Gold, R.; Fox, R.J.; Dawson, K.T.; Amaravadi, L. Evidence of activation of the nrf2 pathway in multiple sclerosis patients treated with delayed-release dimethyl fumarate in the phase 3 define and confirm studies. Mult. Scler. 2017, 23, 1875–1883.
- Gold, R.; Kappos, L.; Arnold, D.L.; Bar-Or, A.; Giovannoni, G.; Selmaj, K.; Tornatore, C.; Sweetser, M.T.; Yang, M.; Sheikh, S.I.; et al. Placebo-controlled phase 3 study of oral bg-12 for relapsing multiple sclerosis. N. Engl. J. Med. 2012, 367, 1098–1107.
- Kraaij, M.D.; Savage, N.D.; van der Kooij, S.W.; Koekkoek, K.; Wang, J.; van den Berg, J.M.; Ottenhoff, T.H.; Kuijpers, T.W.; Holmdahl, R.; van Kooten, C.; et al. Induction of regulatory t cells by macrophages is dependent on production of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2010, 107, 17686–17691.
- George-Chandy, A.; Nordstrom, I.; Nygren, E.; Jonsson, I.M.; Postigo, J.; Collins, L.V.; Eriksson, K. Th17 development and autoimmune arthritis in the absence of reactive oxygen species. Eur. J. Immunol. 2008, 38, 1118–1126.
- Mossberg, N.; Movitz, C.; Hellstrand, K.; Bergstrom, T.; Nilsson, S.; Andersen, O. Oxygen radical production in leukocytes and disease severity in multiple sclerosis. J. Neuroimmunol. 2009, 213, 131–134.
- Mix, E.; Stefan, K.; Hoppner, J.; Klauer, T.; Zettl, U.K.; Strauss, U.; Meyer-Rienecker, H.J.; Rolfs, A. Lymphocyte subpopulations, oxidative burst and apoptosis in peripheral blood cells of patients with multiple sclerosis-effect of interferon-beta. Autoimmunity 2003, 36, 291–305.
- Becanovic, K.; Jagodic, M.; Sheng, J.R.; Dahlman, I.; Aboul-Enein, F.; Wallstrom, E.; Olofsson, P.; Holmdahl, R.; Lassmann, H.; Olsson, T. Advanced intercross line mapping of eae5 reveals ncf-1 and cldn4 as candidate genes for experimental autoimmune encephalomyelitis. J. Immunol. 2006, 176, 6055–6064.
- Linker, R.A.; Lee, D.H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the nrf2 antioxidant pathway. Brain A J. Neurol. 2011, 134, 678–692.
- Li, L.; Sheng, Q.; Zeng, H.; Li, W.; Wang, Q.; Ma, G.; Xu, X.; Qiu, M.; Zhang, W.; Shan, C. Specific genetic aberrations of parathyroid in chinese patients with tertiary hyperparathyroidism using whole-exome sequencing. Front. Endocrinol. 2023, 14, 1221060.
- Messa, P.; Alfieri, C.M. Secondary and tertiary hyperparathyroidism. Front. Horm. Res. 2019, 51, 91–108.
- Alfieri, C.; Mattinzoli, D.; Messa, P. Tertiary and postrenal transplantation hyperparathyroidism. Endocrinol. Metab. Clin. North Am. 2021, 50, 649–662.
- Dream, S.; Chen, H.; Lindeman, B. Tertiary hyperparathyroidism: Why the delay? Ann. Surg. 2021, 273, e120–e122.