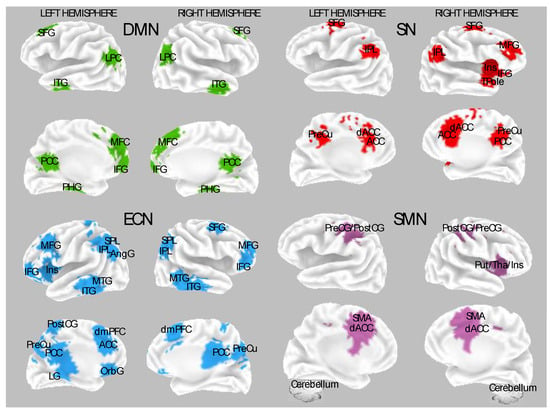
Resting-state functional magnetic resonance imaging (fMRI) and structural MRI investigations offer complementary insights into the distinctive frontal functioning associated with high hypnotizability. Studies indicate that individuals with high hypnotizability display more pronounced reductions in activation of the medial prefrontal or dorsal anterior cingulate cortex following neutral hypnotic induction than their low hypnotizable ones. Moreover, research has disclosed that induction-specific reductions in the activation of the Default Mode Network (DMN) regions correspond to spontaneous changes in cognitive and perceptual states, and the state of attentional absorption during a hypnotic induction has been associated with reduced DMN activity and increased prefrontal attentional system activity.
- functional neuroimaging
- EEG oscillations
- functional connectivity
- hypnosis
1. Introduction
2. Electroencephalography (EEG) Spectral Analysis and Functional Neuroimaging Techniques
2.1. EEG and Neuroimaging
2.2. PET and fMRI Methods
3. Functional Neuroimaging Correlates of Resting Following Hypnotic Induction without Specific Suggestions
4. Structural and Functional Neuroimaging Correlates of Hypnotizability
5. EEG Oscillations and Their Associations with Hypnotizability and Hypnosis
6. EEG Connectivity of Hypnosis and Hypnotizability
6.1. EEG Functional Connectivity Measure
6.2. EEG Functional Connectivity Findings under Resting Hypnosis
6.3. EEG Functional Connectivity Correlates with Hypnotizability
7. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/brainsci14020115
References
- Kihlstrom, J.F. The domain of hypnosis, revisited. In The Oxford Handbook of Hypnosis: Theory, Research and Practice; Oxford University Press: New York, NY, USA, 2008; pp. 21–52.
- Halsband, U.; Mueller, S.; Hinterberger, T.; Strickner, S. Plasticity changes in the brain in hypnosis and meditation. Contemp. Hypn. 2009, 26, 194–215.
- Kihlstrom, J.F.; Glisky, M.L.; McGovern, S.; Rapcsak, S.Z.; Mennemeier, M.S. Hypnosis in the right hemisphere. Cortex 2013, 49, 393–399.
- Oakley, D.A.; Halligan, P.W. Hypnotic suggestion: Opportunities for cognitive neuroscience. Nat. Rev. Neurosci. 2013, 14, 565–576.
- Terhune, D.B.; Cleeremans, A.; Raz, A.; Lynn, S.J. Hypnosis and top-down regulation of consciousness. Neurosci. Biobehav. Rev. 2017, 81, 59–74.
- Vanhaudenhuyse, A.; Laureys, S.; Faymonville, M.E. Neurophysiology of hypnosis. Clin. Neurophysiol. 2014, 44, 343–353.
- Cox, R.E.; Bryant, R.A. Advances in hypnosis research: Methods, designs and contributions of intrinsic and instrumental hypnosis. In The Oxford Handbook of Hypnosis: Theory, Research, and Practice; Oxford University Press: New York, NY, USA, 2008; pp. 311–336.
- McGeown, W.J.; Mazzoni, G.; Venneri, A.; Kirsch, I. Hypnotic induction decreases anterior default mode activity. Conscious. Cogn. 2009, 18, 848–855.
- Oakley, D.A.; Halligan, P.W. Hypnotic suggestion and cognitive neuroscience. Trends Cogn. Sci. 2009, 13, 264–270.
- Rainville, P.; Price, D.D. Hypnosis Phenomenology and the Neurobiology of Consciousness. Int. J. Clin. Exp. Hypn. 2003, 51, 105–129.
- De Pascalis, V.; Scacchia, P.; Vecchio, A. Influences of hypnotic suggestibility, contextual factors, and EEG alpha on placebo analgesia. Am. J. Clin. Hypn. 2021, 63, 302–328.
- Cardeña, E.; Terhune, D.B. Hypnotizability, personality traits, and the propensity to experience alterations of consciousness. Psychol. Conscious. Theory Res. Pract. 2014, 1, 292–307.
- Piccione, C.; Hilgard, E.R.; Zimbardo, P.G. On the degree of stability of measured hypnotizability over a 25-year period. J. Personal. Soc. Psychol. 1989, 56, 289–295.
- Gorassini, D.R.; Spanos, N.P. A social-cognitive skills approach to the successful modification of hypnotic susceptibility. J. Personal. Soc. Psychol. 1986, 50, 1004–1012.
- Gorassini, D.R.; Spanos, N.P. The Carleton Skill Training Program for modifying hypnotic suggestibility: Original version and variations. In Clinical Hypnosis and Self-Regulation: Cognitive-Behavioral Perspectives; American Psychological Association: Washington, DC, USA, 1999; pp. 141–177.
- Rossi, E.L. Hypnosis and Ultradian Cycles: A New State(s) Theory of Hypnosis? Am. J. Clin. Hypn. 1982, 25, 21–32.
- Green, J.P.; Smith, R.J.; Kromer, M. Diurnal Variations in Hypnotic Responsiveness: Is There an Optimal Time to Be Hypnotized? Int. J. Clin. Exp. Hypn. 2015, 63, 171–181.
- Fassler, O.; Lynn, S.J.; Knox, J. Is hypnotic suggestibility a stable trait? Conscious. Cogn. 2008, 17, 240–253.
- Lynn, S.J.; Maxwell, R.; Green, J.P. The hypnotic induction in the broad scheme of hypnosis: A sociocognitive perspective. In Hypnotic Induction, 1st ed.; Routledge: London, UK, 2017; pp. 109–130.
- Lynn, S.J. Hypnotic reponsiveness can be modified after hypnotic training. In Proceedings of the Oral Presentation at XXI World Congress of Medical and Clinical Hypnosis, Montréal, QC, Canada, 22–25 August 2018.
- Gable, P.; Miller, M.; Bernat, E. The Oxford Handbook of EEG Frequency; Oxford University Press: New York, NY, USA, 2022.
- Donoghue, T.; Haller, M.; Peterson, E.J.; Varma, P.; Sebastian, P.; Gao, R.; Noto, T.; Lara, A.H.; Wallis, J.D.; Knight, R.T.; et al. Parameterizing neural power spectra into periodic and aperiodic components. Nat. Neurosci. 2020, 23, 1655–1665.
- Manning, J.R.; Jacobs, J.; Fried, I.; Kahana, M.J. Broadband Shifts in Local Field Potential Power Spectra Are Correlated with Single-Neuron Spiking in Humans. J. Neurosci. 2009, 29, 13613.
- Miller, K.J.; Hermes, D.; Honey, C.J.; Hebb, A.O.; Ramsey, N.F.; Knight, R.T.; Ojemann, J.G.; Fetz, E.E. Human motor cortical activity is selectively phase-entrained on underlying rhythms. PLoS Comput. Biol. 2012, 8, e1002655.
- Ouyang, G.; Hildebrandt, A.; Schmitz, F.; Herrmann, C.S. Decomposing alpha and 1/f brain activities reveals their differential associations with cognitive processing speed. NeuroImage 2020, 205, 116304.
- Güntekin, B.; Başar, E. A review of brain oscillations in perception of faces and emotional pictures. Neuropsychologia 2014, 58, 33–51.
- Buzsáki, G. Rhythms of the Brain; Oxford University Press Inc.: NewYork, NY, USA, 2006.
- Cohen, M.X. Analyzing Neural Time Series Data: Theory and Practice; MIT Press: Cambridge, MA, USA, 2014.
- Keil, A.; Thigpen, N. From Neural Oscillations to Cognitive Processes. In The Oxford Handbook of EEG Frequency; Gable, P.A., Miller, M.W., Bernat, E.M., Eds.; Oxford University Press: New York, NY, USA, 2022; pp. 40–64.
- Ray, W.J.; Oathes, D. Brain Imaging Techniques. Int. J. Clin. Exp. Hypn. 2003, 51, 97–104.
- Michel, C.M.; Murray, M.M. Towards the utilization of EEG as a brain imaging tool. NeuroImage 2012, 61, 371–385.
- De Lucia, M.; Michel, C.M.; Murray, M.M. Comparing ICA-based and Single-Trial Topographic ERP Analyses. Brain Topogr. 2010, 23, 119–127.
- Stephan, K.E.; Friston, K.J. Analyzing effective connectivity with functional magnetic resonance imaging. WIREs Cogn. Sci. 2010, 1, 446–459.
- Kiebel, S.J.; Garrido, M.I.; Moran, R.J.; Friston, K.J. Dynamic causal modelling for EEG and MEG. Cogn. Neurodynamics 2008, 2, 121–136.
- Kaminski, M.J.; Blinowska, K.J. A new method of the description of the information flow in the brain structures. Biol. Cybern. 1991, 65, 203–210.
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198.
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. NeuroImage 2011, 55, 1548–1565.
- Kwong, K.K.; Belliveau, J.W.; Chesler, D.A.; Goldberg, I.E.; Weisskoff, R.M.; Poncelet, B.P.; Kennedy, D.N.; Hoppel, B.E.; Cohen, M.S.; Turner, R. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc. Natl. Acad. Sci. USA 1992, 89, 5675–5679.
- Ogawa, S.; Tank, D.W.; Menon, R.; Ellermann, J.M.; Kim, S.G.; Merkle, H.; Ugurbil, K. Intrinsic signal changes accompanying sensory stimulation: Functional brain mapping with magnetic resonance imaging. Proc. Natl. Acad. Sci. USA 1992, 89, 5951–5955.
- Bandettini, P.A.; Wong, E.C.; Hinks, R.S.; Tikofsky, R.S.; Hyde, J.S. Time course EPI of human brain function during task activation. Magn. Reson. Med. 1992, 25, 390–397.
- Logothetis, N.K.; Pfeuffer, J. On the nature of the BOLD fMRI contrast mechanism. Magn. Reson. Imaging 2004, 22, 1517–1531.
- Landry, M.; Lifshitz, M.; Raz, A. Brain correlates of hypnosis: A systematic review and meta-analytic exploration. Neurosci. Biobehav. Rev. 2017, 81, 75–98.
- Halsband, U. Learning in trance: Functional brain imaging studies and neuropsychology. J. Physiol. Paris 2006, 99, 470–482.
- Egner, T.; Jamieson, G.; Gruzelier, J. Hypnosis decouples cognitive control from conflict monitoring processes of the frontal lobe. NeuroImage 2005, 27, 969–978.
- Faymonville, M.E.; Laureys, S.; Degueldre, C.; Del Fiore, G.; Luxen, A.; Franck, G.; Lamy, M.; Maquet, P. Neural mechanisms of antinociceptive effects of hypnosis. Anesthesiol. Phila. Then Hagerstown 2000, 92, 1257–1267.
- Grond, M.; Pawlik, G.; Walter, H.; Lesch, O.M.; Heiss, W.-D. Hypnotic catalepsy-induced changes of regional cerebral glucose metabolism. Psychiatry Res. Neuroimaging 1995, 61, 173–179.
- Jiang, H.; White, M.P.; Greicius, M.D.; Waelde, L.C.; Spiegel, D. Brain Activity and Functional Connectivity Associated with Hypnosis. Cereb. Cortex 2017, 27, 4083–4093.
- Kosslyn, S.M.; Thompson, W.L.; Costantini-Ferrando, M.F.; Alpert, N.M.; Spiegel, D. Hypnotic Visual Illusion Alters Color Processing in the Brain. Am. J. Psychiatry 2000, 157, 1279–1284.
- Maquet, P.; Faymonville, M.E.; Degueldre, C.; Delfiore, G.; Franck, G.; Luxen, A.; Lamy, M. Functional neuroanatomy of hypnotic state. Biol. Psychiatry 1999, 45, 327–333.
- Rainville, P. Brain mechanisms of pain affect and pain modulation. Curr. Opin. Neurobiol. 2002, 12, 195–204.
- Rainville, P.; Hofbauer, R.K.; Paus, T.; Duncan, G.H.; Bushnell, M.C.; Price, D.D. Cerebral mechanisms of hypnotic induction and suggestion. J. Cogn. Neurosci. 1999, 11, 110–125.
- Szechtman, H.; Woody, E.; Bowers, K.S.; Nahmias, C. Where the imaginal appears real: A positron emission tomography study of auditory hallucinations. Proc. Natl. Acad. Sci. USA 1998, 95, 1956–1960.
- Deeley, Q.; Oakley, D.A.; Toone, B.; Giampietro, V.; Brammer, M.J.; Williams, S.C.R.; Halligan, P.W. Modulating the Default Mode Network Using Hypnosis. Int. J. Clin. Exp. Hypn. 2012, 60, 206–228.
- Demertzi, A.; Soddu, A.; Faymonville, M.E.; Bahri, M.A.; Gosseries, O.; Vanhaudenhuyse, A.; Phillips, C.; Maquet, P.; Noirhomme, Q.; Luxen, A.; et al. Chapter 20-Hypnotic modulation of resting state fMRI default mode and extrinsic network connectivity. In Progress in Brain Research; Van Someren, E.J.W., Van Der Werf, Y.D., Roelfsema, P.R., Mansvelder, H.D., Lopes Da Silva, F.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 193, pp. 309–322.
- Müller, K.; Bacht, K.; Prochnow, D.; Schramm, S.; Seitz, R.J. Activation of thalamus in motor imagery results from gating by hypnosis. NeuroImage 2013, 66, 361–367.
- Müller, K.; Bacht, K.; Schramm, S.; Seitz, R.J. The facilitating effect of clinical hypnosis on motor imagery: An fMRI study. Behav. Brain Res. 2012, 231, 164–169.
- Vanhaudenhuyse, A.; Boly, M.; Balteau, E.; Schnakers, C.; Moonen, G.; Luxen, A.; Lamy, M.; Degueldre, C.; Brichant, J.F.; Maquet, P.; et al. Pain and non-pain processing during hypnosis: A thulium-YAG event-related fMRI study. NeuroImage 2009, 47, 1047–1054.
- Rainville, P.; Streff, A.; Chen, J.-I.; Houzé, B.; Desmarteaux, C.; Piché, M. Hypnotic Automaticity in the Brain at Rest: An Arterial Spin Labelling Study. Int. J. Clin. Exp. Hypn. 2019, 67, 512–542.
- Dienes, Z.; Beran, M.; Brandl, J.L.; Perner, J.; Proust, J. Is hypnotic responding the strategic relinquishment of metacognition. In Foundations of Metacognition; Oxford University Press: New York, NY, USA, 2012; pp. 267–277.
- Jaffer, U.; Jamieson, G.A. Hypnosis and the neuroscience of cognitive and affective control. Aust. J. Clin. Exp. Hypn. 2012, 40, 1–20.
- Chambon, V.; Wenke, D.; Fleming, S.M.; Prinz, W.; Haggard, P. An Online Neural Substrate for Sense of Agency. Cereb. Cortex 2013, 23, 1031–1037.
- Haggard, P.; Chambon, V. Sense of agency. Curr. Biol. 2012, 22, R390–R392.
- Lush, P.; Naish, P.; Dienes, Z. Metacognition of intentions in mindfulness and hypnosis. Neurosci. Conscious. 2016, 2016, niw007.
- Buchner, R.; Andrews-Hanna, J.; Schacter, D. The brain’s default network. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38.
- Sridharan, D.; Levitin, D.J.; Menon, V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc. Natl. Acad. Sci. USA 2008, 105, 12569–12574.
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711.
- Halsband, U.; Gerhard Wolf, T. Functional Changes in Brain Activity After Hypnosis: Neurobiological Mechanisms and Application to Patients with a Specific Phobia—Limitations and Future Directions. Int. J. Clin. Exp. Hypn. 2019, 67, 449–474.
- Jensen, M.P.; Adachi, T.; Hakimian, S. Brain Oscillations, Hypnosis, and Hypnotizability. Am. J. Clin. Hypn. 2015, 57, 230–253.
- Halsband, U.; Wolf, T.G. Current neuroscientific research database findings of brain activity changes after hypnosis. Am. J. Clin. Hypn. 2021, 63, 372–388.
- Wolf, T.G.; Faerber, K.A.; Rummel, C.; Halsband, U.; Campus, G. Functional Changes in Brain Activity Using Hypnosis: A Systematic Review. Brain Sci. 2022, 12, 108.
- Faymonville, M.E.; Boly, M.; Laureys, S. Functional neuroanatomy of the hypnotic state. J. Physiol.-Paris 2006, 99, 463–469.
- van Osch, M.J.P.; Teeuwisse, W.M.; Chen, Z.; Suzuki, Y.; Helle, M.; Schmid, S. Advances in arterial spin labelling MRI methods for measuring perfusion and collateral flow. J. Cereb. Blood Flow Metab. 2018, 38, 1461–1480.
- Blakemore, S.J.; Oakley, D.A.; Frith, C.D. Delusions of alien control in the normal brain. Neuropsychologia 2003, 41, 1058–1067.
- Darby, R.R.; Joutsa, J.; Burke, M.J.; Fox, M.D. Lesion network localization of free will. Proc. Natl. Acad. Sci. USA 2018, 115, 10792–10797.
- Jensen, M.P.; Jamieson, G.A.; Lutz, A.; Mazzoni, G.; McGeown, W.J.; Santarcangelo, E.L.; Demertzi, A.; De Pascalis, V.; Bányai, É.I.; Rominger, C.; et al. New directions in hypnosis research: Strategies for advancing the cognitive and clinical neuroscience of hypnosis. Neurosci. Conscious. 2017, 2017, nix004.
- Barnier, A.J.; Cox, R.E.; McConkey, K.M. The province of “highs”: The high hypnotizable person in the science of hypnosis and in psychological science. Psychol. Conscious. Theory Res. Pract. 2014, 1, 168–183.
- Heap, M.; Brown, R.J.; Oakley, D.A. The Highly Hypnotizable Person: Theoretical, Experimental and Clinical Issues; Routledge: London, UK, 2004.
- Cardeña, E.; Jönsson, P.; Terhune, D.B.; Marcusson-Clavertz, D. The neurophenomenology of neutral hypnosis. Cortex 2013, 49, 375–385.
- Pekala, R.J.; Kumar, V. An empirical-phenomenological approach to quantifying consciousness and states of consciousness: With particular reference to understanding the nature of hypnosis. In Hypnosis and Conscious States: The Cognitive Neuroscience Perspective; Jamieson, G.A., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 167–194.
- McGeown, W.J.; Mazzoni, G.; Vannucci, M.; Venneri, A. Structural and functional correlates of hypnotic depth and suggestibility. Psychiatry Res. Neuroimaging 2015, 231, 151–159.
- Huber, A.; Lui, F.; Duzzi, D.; Pagnoni, G.; Porro, C.A. Structural and functional cerebral correlates of hypnotic suggestibility. PLoS ONE 2014, 9, e93187.
- Horton, J.E.; Crawford, H.J.; Harrington, G.; Downs, J.H., III. Increased anterior corpus callosum size associated positively with hypnotizability and the ability to control pain. Brain 2004, 127, 1741–1747.
- Bloom, J.S.; Hynd, G.W. The Role of the Corpus Callosum in Interhemispheric Transfer of Information: Excitation or Inhibition? Neuropsychol. Rev. 2005, 15, 59–71.
- DeSouza, D.D.; Stimpson, K.H.; Baltusis, L.; Sacchet, M.D.; Gu, M.; Hurd, R.; Wu, H.; Yeomans, D.C.; Willliams, N.; Spiegel, D. Association between Anterior Cingulate NHuberbeurochemical Concentration and Individual Differences in Hypnotizability. Cereb. Cortex 2020, 30, 3644–3654.
- Hoeft, F.; Gabrieli, J.D.E.; Whitfield-Gabrieli, S.; Haas, B.W.; Bammer, R.; Menon, V.; Spiegel, D. Functional Brain Basis of Hypnotizability. Arch. Gen. Psychiatry 2012, 69, 1064–1072.
- Cojan, Y.; Piguet, C.; Vuilleumier, P. What makes your brain suggestible? Hypnotizability is associated with differential brain activity during attention outside hypnosis. NeuroImage 2015, 117, 367–374.
- Kihlstrom, J.F. Neuro-hypnotism: Prospects for hypnosis and neuroscience. Cortex 2013, 49, 365–374.
- Crawford, H.J.; Gruzelier, J.H. A midstream view of the neuropsychophysiology of hypnosis: Recent research and future directions. In Contemporary Hypnosis Research; Guilford Press: New York, NY, USA, 1992; pp. 227–266.
- Gruzelier, J.H. A working model of the neurophysiology of hypnosis: A review of evidence. Contemp. Hypn. 1998, 15, 3–21.
- De Pascalis, V. Psychophysiological correlates of hypnosis and hypnotic susceptibility. Int. J. Clin. Exp. Hypn. 1999, 47, 117–143.
- Kallio, S.; Revonsuo, A.; Hämäläinen, H.; Markela, J.; Gruzelier, J. Anterior brain functions and hypnosis: A test of the frontal hypothesis. Int. J. Clin. Exp. Hypn. 2001, 49, 95–108.
- Huber, A.; Lui, F.; Porro, C.A. Hypnotic susceptibility modulates brain activity related to experimental placebo analgesia. PAIN® 2013, 154, 1509–1518.
- Sheehan, P.; McConkey, K. Hypnosis and Experience (Psychology Revivals): The Exploration of Phenomena and Process; Routledge: London, UK, 2015.
- Bryant, R.A.; McConkey, K.M. Hypnotic blindness and the relevance of cognitive style. J. Personal. Soc. Psychol. 1990, 59, 756–761.
- Oakley, D.A. Hypnosis and Conversion Hysteria: A Unifying Model. Cogn. Neuropsychiatry 1999, 4, 243–265.
- Oakley, D.; Frasquilho, F. Hypnotic susceptibility, or F-bias: Its relevance to eating disorders. Contemp. Hypn. 1998, 15, 40–51.
- McConkey, K.M.; Glisky, M.L.; Kihlstrom, J.F. Individual differences among hypnotic virtuosos: A case comparison. Aust. J. Clin. Exp. Hypn. 1989, 17, 131–140.
- Santarcangelo, E.L. New views of hypnotizability. Front. Behav. Neurosci. 2014, 8, 224.
- Picerni, E.; Santarcangelo, E.L.; Laricchiuta, D.; Cutuli, D.; Petrosini, L.; Spalletta, G.; Piras, F. Cerebellar Structural Variations in Subjects with Different Hypnotizability. Cerebellum 2019, 18, 109–118.
- Santarcangelo, E.L.; Scattina, E. Responding to Sensorimotor Suggestions: From Endothelial Nitric Oxide to the Functional Equivalence between Imagery and Perception. Int. J. Clin. Exp. Hypn. 2019, 67, 394–407.
- Dienes, Z.; Hutton, S. Understanding hypnosis metacognitively: rTMS applied to left DLPFC increases hypnotic suggestibility. Cortex 2013, 49, 386–392.
- Coltheart, M.; Cox, R.; Sowman, P.; Morgan, H.; Barnier, A.; Langdon, R.; Connaughton, E.; Teichmann, L.; Williams, N.; Polito, V. Belief, delusion, hypnosis, and the right dorsolateral prefrontal cortex: A transcranial magnetic stimulation study. Cortex 2018, 101, 234–248.
- Bakan, P. Hypnotizability, Laterality of Eye-Movements and Functional Brain Asymmetry. Percept. Mot. Ski. 1969, 28, 927–932.
- Edmonston, W.E.; Grotevant, W.R. Hypnosis and Alpha Density. Am. J. Clin. Hypn. 1975, 17, 221–232.
- Engstrom, D.R.; London, P.; Hart, J.T. Hypnotic Susceptibility increased by EEG Alpha Training. Nature 1970, 227, 1261–1262.
- London, P.; Hart, J.T.; Leibovitz, M.P.; McDevitt, R.A. The psychophysiology of hypnotic susceptibility. In Psychophysiological Mechanisms of Hypnosis: An International Symposium Sponsored by the International Brain Research Organization and the Centre de Recherche, Institut de Psychiatrie La Rochefoucauld, Paris; Springer: Berlin/Heidelberg, Germany, 1969; pp. 151–172.
- Morgan, A.H.; Macdonald, H.; Hilgard, E.R. EEG Alpha: Lateral Asymmetry Related to Task, and Hypnotizability. Psychophysiology 1974, 11, 275–282.
- Ulett, G.A.; Akpinar, S.; Itil, T.M. Quantitative EEG analysis during hypnosis. Electroencephalogr. Clin. Neurophysiol. 1972, 33, 361–368.
- De Pascalis, V.; Palumbo, G. EEG Alpha Asymmetry: Task Difficulty and Hypnotizability. Percept. Mot. Ski. 1986, 62, 139–150.
- Graffin, N.F.; Ray, W.J.; Lundy, R. EEG concomitants of hypnosis and hypnotic susceptibility. J. Abnorm. Psychol. 1995, 104, 123–131.
- MacLeod-Morgan, C. Hypnotic susceptibility, EEG theta and alpha waves, and hemispheric specificity. In Hypnosis 1979; Elsevier: Amsterdam, The Netherlands, 1979.
- Williams, J.D.; Gruzelier, J.H. Differentiation of hypnosis and relaxation by analysis of narrow band theta and alpha frequencies. Int. J. Clin. Exp. Hypn. 2001, 49, 185–206.
- Sabourin, M.E.; Cutcomb, S.D.; Crawford, H.J.; Pribram, K. EEG correlates of hypnotic susceptibility and hypnotic trance: Spectral analysis and coherence. Int. J. Psychophysiol. 1990, 10, 125–142.
- Franz, M.; Schmidt, B.; Hecht, H.; Naumann, E.; Miltner, W.H.R. Suggested visual blockade during hypnosis: Top-down modulation of stimulus processing in a visual oddball task. PLoS ONE 2021, 16, e0257380.
- Schmidt, B.; Hecht, H.; Naumann, E.; Miltner, W.H.R. The Power of mind: Blocking visual perception by hypnosis. Sci. Rep. 2017, 7, 4889.
- De Pascalis, V. EEG oscillatory activity concomitant with hypnosis and hypnotizability. In International Handbook of Clinical Hypnosis; Linden, J.H., De Benedittis, G., Sugarman, L.I., Varga, K., Eds.; Routledge: London, UK, 2024; in press.
- Crawford, H.J. Cognitive and psychophysiological correlates of hypnotic responsiveness and hypnosis. In Creative Mastery in Hypnosis and Hypnoanalysis: A Festschrift for Erika Fromm; Plenum Press: New York, NY, USA, 1990; pp. 47–54.
- De Pascalis, V.; Ray, W.J.; Tranquillo, I.; D’Amico, D. EEG activity and heart rate during recall of emotional events in hypnosis: Relationships with hypnotizability and suggestibility. Int. J. Psychophysiol. 1998, 29, 255–275.
- Tebecis, A.K.; Provins, K.A.; Farnbach, R.W.; Pentony, P. Hypnosis and the EEG: A quantitative investigation. J. Nerv. Ment. Dis. 1975, 161, 1–17.
- Crawford, H.J.; Clarke, S.W.; Kitner-Triolo, M. Self-generated happy and sad emotions in low and highly hypnotizable persons during waking and hypnosis: Laterality and regional EEG activity differences. Int. J. Psychophysiol. 1996, 24, 239–266.
- De Benedittis, G. Neural mechanisms of hypnosis and meditation. J. Physiol.-Paris 2015, 109, 152–164.
- Jensen, M.P.; Adachi, T.; Tomé-Pires, C.; Lee, J.; Osman, Z.J.; Miró, J. Mechanisms of Hypnosis: Toward the Development of a Biopsychosocial Model. Int. J. Clin. Exp. Hypn. 2015, 63, 34–75.
- De Pascalis, V.; Marucci, F.S.; Penna, P.M.; Pessa, E. Hemispheric activity of 40 Hz EEG during recall of emotional events: Differences between low and high hypnotizables. Int. J. Psychophysiol. 1987, 5, 167–180.
- De Pascalis, V.; Marucci, F.S.; Penna, P.M. 40-Hz EEG asymmetry during recall of emotional events in waking and hypnosis: Differences between low and high hypnotizables. Int. J. Psychophysiol. 1989, 7, 85–96.
- Crawford, H.J. Brain dynamics and hypnosis: Attentional and disattentional processes. Int. J. Clin. Exp. Hypn. 1994, 42, 204–232.
- Jamieson, G.A.; Burgess, A.P. Hypnotic induction is followed by state-like changes in the organization of EEG functional connectivity in the theta and beta frequency bands in high-hypnotically susceptible individuals. Front. Hum. Neurosci. 2014, 8, 528.
- White, D.; Ciorciari, J.; Carbis, C.; Liley, D. EEG Correlates of Virtual Reality Hypnosis. Int. J. Clin. Exp. Hypn. 2009, 57, 94–116.
- Knauff, M.; Mulack, T.; Kassubek, J.; Salih, H.R.; Greenlee, M.W. Spatial imagery in deductive reasoning: A functional MRI study. Cogn. Brain Res. 2002, 13, 203–212.
- Kirenskaya, A.V.; Novototsky-Vlasov, V.Y.; Zvonikov, V.M. Waking EEG Spectral Power and Coherence Differences Between High and Low Hypnotizable Subjects. Int. J. Clin. Exp. Hypn. 2011, 59, 441–453.
- Schnyer, D.M.; Allen, J.J. Attention-related electroencephalographic and event-related potential predictors of responsiveness to suggested posthypnotic amnesia. Int. J. Clin. Exp. Hypn. 1995, 43, 295–315.
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kähkönen, S. Functional connectivity in the brain—Is it an elusive concept? Neurosci. Biobehav. Rev. 2005, 28, 827–836.
- Lee, L.; Harrison, L.M.; Mechelli, A. A report of the functional connectivity workshop, Dusseldorf 2002. NeuroImage 2003, 19, 457–465.
- Varela, F.; Lachaux, J.-P.; Rodriguez, E.; Martinerie, J. The brainweb: Phase synchronization and large-scale integration. Nat. Rev. Neurosci. 2001, 2, 229–239.
- Tsakiris, M.; Hesse, M.D.; Boy, C.; Haggard, P.; Fink, G.R. Neural Signatures of Body Ownership: A Sensory Network for Bodily Self-Consciousness. Cereb. Cortex 2007, 17, 2235–2244.
- Pascual-Marqui, R.D. Instantaneous and lagged measurements of linear and nonlinear dependence between groups of multivariate time series: Frequency decomposition. arXiv 2007, arXiv:0711.1455.
- Bullock, T.H.; McClune, M.C.; Enright, J.T. Are the electroencephalograms mainly rhythmic? Assessment of periodicity in wide-band time series. Neuroscience 2003, 121, 233–252.
- Fingelkurts, A.A.; Fingelkurts, A.A.; Kallio, S.; Revonsuo, A. Cortex functional connectivity as a neurophysiological correlate of hypnosis: An EEG case study. Neuropsychologia 2007, 45, 1452–1462.
- De Pascalis, V. Phase-ordered gamma oscillations and the modulation of hypnotic experience. In Hypnosis and Conscious States: The Cognitive Neuroscience Perspective; Jamieson, G.A., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 67–89.
- Woody, E.Z.; Sadler, P. Dissociation theories of hypnosis. In The Oxford Handbook of Hypnosis: Theory, Research, and Practice; Oxford University Press Inc.: New York, NY, USA, 2008; pp. 81–110.
- Woody, E.Z.; Farvolden, P. Dissociation in Hypnosis and Frontal Executive Function. Am. J. Clin. Hypn. 1998, 40, 206–216.
- Jamieson, G.A.; Woody, E. Dissociated control as a paradigm for cognitive neuroscience research and theorizing in hypnosis. In Hypnosis and Conscious States: The Cognitive Neuroscience Perspective; Jamieson, G.A., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 111–129.
- Dienes, Z.; Perner, J. Executive control without conscious awareness: The cold control theory of hypnosis. In Hypnosis and Conscious States: The Cognitive Neuroscience Perspective; Jamieson, G.A., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 293–314.
- Demertzi, A.; Vanhaudenhuyse, A.; Noirhomme, Q.; Faymonville, M.-E.; Laureys, S. Hypnosis modulates behavioural measures and subjective ratings about external and internal awareness. J. Physiol.-Paris 2015, 109, 173–179.
- Lipari, S.; Baglio, F.; Griffanti, L.; Mendozzi, L.; Garegnani, M.; Motta, A.; Cecconi, P.; Pugnetti, L. Altered and asymmetric default mode network activity in a “hypnotic virtuoso”: An fMRI and EEG study. Conscious. Cogn. 2012, 21, 393–400.
- Faerman, A.; Spiegel, D. Shared cognitive mechanisms of hypnotizability with executive functioning and information salience. Sci. Rep. 2021, 11, 5704.