There exist two main biological types of seeds that have been termed “orthodox” and “recalcitrant” seeds
[13]. The majority of seeds referred to as orthodox require desiccation tolerance during their development, allowing them to be stored for long periods under air-dry storage
[3][26]. On the other hand, recalcitrant seeds have a high moisture content at shedding and do not tolerate desiccation; therefore, they are able to be stored in a dry state
[18][27][28][29]. More recently, a third category of seeds, named “intermediate”, has been defined
[30][31]; they survive the loss of water, but they become damaged and die during dry storage at low temperatures.
The Compendium of Information on Seed Storage Behaviour [17][32] recognizes these three biological types.
“Orthodox” seeds generally undergo dehydration prior to shedding; they are desiccation tolerant and can be stored successfully in a dehydrated state at a temperature below freezing for very long periods (decades or longer). At maturity (or harvest), they contain no more than 10–15% water and survive drying to very low moisture content (3–5%). They therefore tolerate subsequent storage at sub-zero temperatures
[33]. Orthodox seeds acquire desiccation tolerance relatively early during their development and usually before the maturation drying phase. Several metabolic changes occur with respect to the protection of seed cells against dehydration damage
[18][26][33][34][35]. In particular, carbohydrate metabolism
[36][37] and specific proteins (dehydrins, late embryogenesis abundant proteins: LEA, and heat shock proteins: HSPs)
[33] seem to be involved in this process. Some soluble sugars, such as sucrose and oligosaccharides (raffinose, stachyose, and verbascose), might also play an important part in this process by facilitating the stabilization of lipids and proteins in cell membranes or by promoting the vitrification of water and then the protection of cytosolic structures
[35][37][38].
The repartition of the three categories of seeds (orthodox, intermediate, and recalcitrant) depends on the family. At the family level, the percentage of recalcitrant seeds varies from 77.1–80.0% in the Lauraceae and the Fagaceae to less than 2% in the Melastomataceae, with it being around 65.4% in the Sapotaceae, 42.1–48.8% in the Clusiaceae and the Moraceae, and 25.8–31.0% in the Arecaceae and Sapindaceae
[1][69]. All of these families also present orthodox seeds: 3.8% (Sapotaceae), 17.4% (Fagaceae), 27.8% (Aracaceae), 50–51.2% (Maraceae and Rutaceae), 57.9% (Clusiaceae), 63.1% (Sapindaceae), and 83.9% (Myrtaceae). The majority of angiosperms (Asteraceae, Brassicaceae, Caryophyllaceae, Cyperaceae, Ericaceae, Gentianaceae, Poaceae, Ranunuculaceae, Scrophylariaceae, and Solanaceae) have 95–100% orthodox seeds
[69]. The intermediate seeds characterize some species such as the Arecaceae, the Rutaceae, the Sapotaceae, and the Zingiberaceae
[69].
Macrobiotic seeds are defined as seeds capable of surviving for more than 15 years in the soil or under ambient conditions, but seeds with longevity exceeding 50 or 100 years are far from rare. They are often hard seeds (i.e., seeds with seed coats impermeable to water). This has been shown by Becquerel
[71] who succeeded in germinating very old seeds from the collections of the Natural History Museum of Paris and estimated their longevity as 55 years (
Melilotus lutea), 63–68 years (
Cytisus austriacus, Lavatera pseudo-olbia, Ervum lens, and
Trifolium arvense), 81–86 years (
Mimosa glomerata, Cytisus biflorus, and
Astragalus massiliensis), and a period of 100–158 years (
Cassia multijuga and
Dioclea pauciflora)
[1]. Evidence from hundreds to thousands of years of seeds has been described
[1]. Lotus (
Nelumbo nucifera, the Indian lotus) seeds are also well known for their longevity of several hundred years
[1]. For example, Dum
[72] also achieved the germination of
Chenopodium album and
Spergula arvensis seeds that were about 1700 years old. The literature reveals that the record is held by
Lupinus arcticus seeds which successfully germinated despite being more than 10,000 years old
[73]. However, such long longevity must be regarded with skepticism without direct dating of the seeds
[1][34]. Radiocarbon dating allows us to determine the age of seeds at about 2000 years for the date (
Phoenix dactylifera)
[74], at 1300 years for lotus (
Nelumbo nucifera)
[75], and at 600 years for canna (
Canna compacta)
[76]. The myth concerning the longevity of mummy grains discovered in Egyptian tombs and supposed to remain able to germinate is in fact a mistake or a hoax
[1][34].
Although this classification gives information concerning the putative survival behavior of numerous species, it is debatable because it does not take into account the main factors of storage (temperature and seed moisture content). Indeed, seed survival is both genetically and environmentally controlled. Depending on the conditions of storage, microbiotic seeds could become mesobiotic or macrobiotic ones.
4. Loss of Seed Viability
4.1. Change in Viability during Storage: Viability Equations
At harvest, the initial seed viability is a product of the seed history through development on the mother plant
[1][34]. Subsequent seed longevity depends on post-harvest treatments (drying, cleaning, sorting, coating, etc.) and the conditions of storage (temperature, moisture content, and oxygen availability)
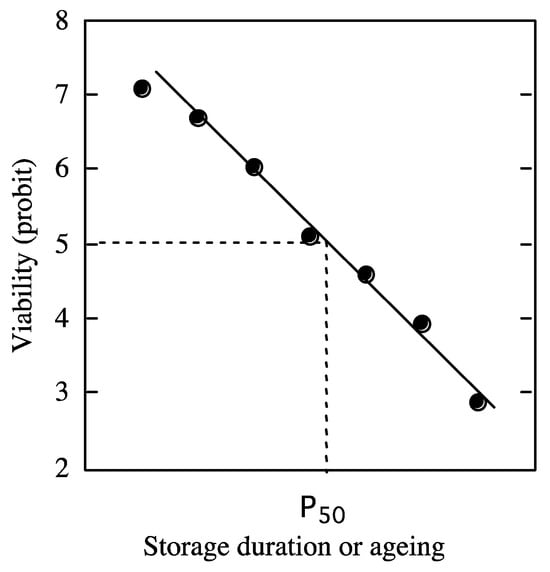
[34]. The viability equations are based on fitting a negative cumulative normal distribution to viability percentages. The conversion of the negatively sigmoidal curve obtained to probits linearizes the curve
[1][2][34][78] (
Figure 1).
Figure 1. Theoretical curve of loss of viability in probit during dry storage. A probit value of 5 determines the half-viability period. From
[77].
Roberts and Coll
[13][79][80][81] developed and generalized a descriptive equation, known as the “viability equation”, taking into account the conditions of seed storage on the loss of viability:
where v represents the probit % germination after p days of storage; Ki is the probit of initial germinability for the seed lot; K
E, C
w, C
H, and C
Q are constants depending on the species; m is the seed moisture content (expressed on a fresh weight basis); and t is the storage temperature in °C.
This equation indicates that longevity increases as the storage temperature decreases. Dickie et al.
[82] working on eight species (barley, chickpea, cowpea, soybean, elm, mahogany, terb, and lettuce) placed over a wide range of storage conditions (temperature from −13 to 90 °C; moisture content from 1.8 to 25% fresh weight) demonstrated that the temperature coefficients (C
H and C
Q) of the equation do not differ significantly between these species, with them being equal to 0.0329 and 0.000478, respectively, i.e., the effect of temperature on longevity appears to be similar. In contrast, the effect of moisture content on longevity evaluated by the coefficient C
W differs between species
[83][84][85]. This variation in C
W results in part from differences in seed composition; for example, the higher the seed oil content, the lower the value of C
W [1][2]. Data obtained by Ellis et al.
[84][85] indicate that longevity is doubled for each 8.4–8.7% reduction in seed equilibrium in relative humidity between 90 and 10%
[2].
Roberts
[13][79] derived a simple mathematical equation, allowing us to calculate the P
50 value or half-viability period.
where P
50 is the time taken for 50% of the seed population to lose viability, m is the moisture content expressed on a fresh weight basis, t is the temperature in °C and K
v, and C
1 and C
2 are constants.
Figure 1 shows a theoretical curve of loss of viability in probit during ageing, and
Table 4 gives some examples of an estimated half-viability period of seeds from various cultivated species stored under open storage conditions in a temperate climate with the mean temperature at about 10 °C and the average RH at about 60–75%
[1][86]. Under these conditions, the P
50 value varies from 3.4–4.1 years (parsley, parsnip, and celery) to 10.5–15.9 years (lucerne, French bean, garden pea, and broad bean) and 24.5 years for tomato. Generally, seeds containing a high amount of starch (e.g., cereals) can be stored well (7.2–12.9 years), but this is not the case for rye (4.5 years). Seeds containing high levels of oil are often considered to be relatively short-lived, but this link between the amount of seed oil and P
50 is debatable: for example, tomato seeds that contain a high level of oil (more than 30%) have high longevity (P
50 = 24.5 years) whilst onion seeds that are poor in oil (often less than 10%) are difficult to store (a P
50 of about 5.4 years). On the other hand, oily seeds such as soybean and sunflower are characterized by a short P
50 (3.4 and 5.4 years, respectively), while the P
50 of rape seeds is about 13.9 years.
Table 4. Estimated half-viability period (P
50) of some cultivated species. The seeds are stored under open storage conditions in a temperate climate where the mean temperature is 10 °C and the average RH is about 60–75%. Modified from
[1][77][86].
It is often difficult to compare the data concerning seed longevity across species and seed lots because the conditions of storage are different depending on the laboratory. For example, when seeds were stored at 5 °C with 5 ± 2% water in the USDA National Seed Storage Laboratory, the P
50 calculated
[86] was higher than that obtained in open storage (as indicated in
Table 4). It reaches 53 years for
Helianthus annuus (sunflower), 36 years for
Secale cereale (rye), 30 years for
Glycine max (soybean), 22 years for
Lactuca sativa (lettuce), and 18 years for
Allium cepa (onion) and is longer than 80 years for
Medicago sativa (lucerne),
Pisum sativum (garden pea), and
Lycopersicon esculentum (tomato).
Hay et al.
[87][88] discussed and described the most widely adopted protocols that are used to measure seed longevity, in particular, a “comparative longevity protocol” established at the Millennium Seed Bank (MSB) of the Royal Botanic Gardens, Kew
[89]. This protocol suggests choosing 60% RH and 45 °C; these conditions allow one to obtain data within an acceptable length of time.
4.2. Modulation of Viability by Storage Conditions
As indicated by the “improved viability equation”, loss of seed viability is mainly regulated by seed moisture content and temperature during storage. The storage of orthodox seeds follows two rules
[1][2][4][7][34][79][80]:
- -
-
For each 1–2% decrease in seed moisture content (when the MC ranges between 5 and 14%), the seed storage life is doubled;
- -
-
For each 10 °F (5.6 °C) decrease in seed storage temperature (between 0 °C and 50 °C), the seed storage life is doubled.
However, maximum seed longevity is achieved at a critical low moisture content limit for the application of the viability equation, and drying the seeds below this critical value does not improve seed longevity
[8][30][84][85][90][91]. It varies among species and depends on the seed composition, in particular the lipid content and temperature. It is for example about 2% (fresh weight basis) in
Arachis hypogea [85] and 6.2% in
Pisum sativa [84], and it varies in an inverse relationship with the lipid content
[8][84][85]. Maximum survival after 4–5 years at ambient temperature was determined to be in the range of 1.8–2.5% for sesame, 4.3–5% for soybean, and 7.6–9.7% for wheat
[92]. In addition, this critical moisture content also decreases with increasing temperature, with it ranging between 3 and 4% at high temperatures and 4 and6% at ambient temperature
[93].
The availability of oxygen (i.e., hermetic vs. open storage) can also influence seed viability during storage
[94][95][96]. Oxygen is generally detrimental to seed viability maintenance, but the beneficial effect of low oxygen or anaerobic conditions depends on the conditions of ageing. Ellis and Hong
[95] indicate that this negative effect increases as the seed moisture content decreases. Storage in N
2 (i.e., in the absence of oxygen) is advantageous for pea, broad beans, and barley
[97], but it has no significant effect on cabbage, onion, and red clover
[98]. In the case of non-primed and primed lettuce seeds stored in low RH (33%), longevity was extended in an anaerobic environment, but this effect was less under storage in controlled deterioration (75% RH, 50 °C)
[96]. In onion, this beneficial effect of anaerobic conditions is only observed in primed seeds
[96].
4.3. Procedures for Long-Term Storage in Genebanks
Improving seed storage techniques is a major research focus of the International Plant Genetic Resources Institute (IPGRI). Guidelines for long-term conservation recommend storage at −20 °C ± 4 °C and 15 ± 3% RH, considered a conventional method
[99]. Cryopreservation is also a possible technique to prolong the longevity of orthodox seeds with short lifespans
[100]. In contrast, recalcitrant and intermediate seeds require cryopreservation
[7][101][102].
To achieve the appropriate moisture content, Kew recommends equilibrating the seeds to 15% RH at 15 °C (
www.rbgkew.org.uk/, accessed on 1 November 2023), Ellis et al.
[30][84][85] suggest equilibrating the seeds to 10% RH at 20 °C, and Vertucci and Roos
[90][103] propose to equilibrate the seeds at 20–25% RH at storage temperature. Storing seeds in hermetically sealed containers to maintain their water content is also recommended.
Most orthodox seeds also show long longevity when ultra-dried or freeze-dried. One advantage of ultra-drying or freeze-drying is that seeds can be stored at room temperature, but they must be maintained under vacuum in tightly closed containers or bags impermeable to water vapor.
Table 5 shows that freeze-drying gives excellent results with the seeds of some vegetable species compared to storage under ambient conditions
[62][77]. However, large seeds, such as pea, soybean, and bean, can be damaged by freeze-drying, with cracks in the cotyledons occurring at thawing or during seed re-imbibition. The results obtained with vacuum-dried seeds (seeds dried under vacuum for 3 days at 20 °C) and freeze-dried seeds (seeds immersed in liquid nitrogen and placed under vacuum for 3 days at 20°) and then stored under vacuum in the presence of silica gel for 12–19 months at 5, 20, and 30 °C clearly indicated that the responsiveness of ultra-dried seeds to storage depends on the species and the temperature of storage. Ultra-dried seeds of lettuce and leek remained viable after 12 months of storage although the germination rate of leek seeds and freeze–dried lettuce seeds was slightly reduced. In onion and lamb’s lettuce seeds, ultra-drying resulted in a marked decrease in seed viability after 12 months of storage, and this deleterious effect was reinforced by increasing the temperature of germination. In addition, ultra-dried seeds stored for 12 months were more sensitive to accelerated ageing treatment (40–45 °C, 100% RH) suggesting that the seeds became progressively less vigorous during storage.
Table 5. Comparison of the longevity of seeds from some vegetable species stored in the open air or freeze-dried and stored under vacuum at ambient temperature. C, control non-freeze-dried seeds; FD, freeze-dried seeds. Modified from
[62][77].