Nuclear receptor-binding SET domain protein 3 (NSD3) is a member of the NSD histone methyltransferase family of proteins. In recent years, it has been identified as a potential oncogene in certain types of cancer. The NSD3 gene encodes three isoforms, the long version (NSD3L), a short version (NSD3S) and the WHISTLE isoforms. Importantly, the NSD3S isoform corresponds to the N-terminal region of the full-length protein, lacking the methyltransferase domain. The chromosomal location of NSD3 is frequently amplified across cancer types, such as breast, lung, and colon, among others. This amplification has been correlated to a chromothripsis event, that could explain the different NSD3 alterations found in cancer. The fusion proteins containing NSD3 have also been reported in leukemia (NSD3-NUP98), and in NUT (nuclear protein of the testis) midline carcinoma (NSD3-NUT). Its role as an oncogene has been described by modulating different cancer pathways through its methyltransferase activity, or the short isoform of the protein, through protein interactions.
- cancer
- molecular oncology
- oncogenes
- NSD3
- NSD3S
1. Introduction
2. NSD3 Protein Structure
3. NSD3 Alterations in Cancer
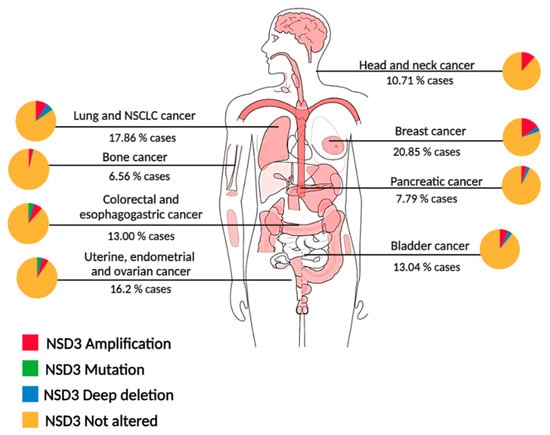
3.1. Study of the Amplicon 8p11-12: Chromothripsis
3.1.1. Amplification
3.1.2. Fusion Proteins

3.1.3. Mutations
4. NSD3 Involvement in Cancer
4.1. Methyltransferase-Dependent Function of NSD3 in Cancer
4.1.1. NOTCH Pathway
4.1.2. mTOR Pathway
4.1.3. EGFR Pathway
4.1.4. IFN Pathway
4.1.5. Cyclin Dependent Kinase (CDK) Pathway
4.2. NSD3S Isoform Function as an Adaptor Protein
4.2.1. NSD3-NUT Fusion
4.2.2. NSD3S-BRD4-CHD8 Interactions
4.2.3. NSD3S-MYC Interaction
This entry is adapted from the peer-reviewed paper 10.3390/ijms25020944
References
- Kornberg, R.D.; Lorch, Y. Twenty-Five Years of the Nucleosome, Fundamental Particle of the Eukaryote Chromosome. Cell 1999, 98, 285–294.
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-Resolution Profiling of Histone Methylations in the Human Genome. Cell 2007, 129, 823–837.
- Kouzarides, T. Histone Methylation in Transcriptional Control. Curr. Opin. Genet. Dev. 2002, 12, 198–209.
- Morishita, M.; Di Luccio, E. Structural Insights into the Regulation and the Recognition of Histone Marks by the SET Domain of NSD1. Biochem. Biophys. Res. Commun. 2011, 412, 214–219.
- Li, W.; Tian, W.; Yuan, G.; Deng, P.; Sengupta, D.; Cheng, Z.; Cao, Y.; Ren, J.; Qin, Y.; Zhou, Y.; et al. Molecular Basis of Nucleosomal H3K36 Methylation by NSD Methyltransferases. Nature 2021, 590, 498–503.
- Douglas, J.; Coleman, K.; Tatton-Brown, K.; Hughes, H.E.; Temple, I.K.; Cole, T.R.P.; Rahman, N. Evaluation of NSD2 and NSD3 in Overgrowth Syndromes. Eur. J. Hum. Genet. 2005, 13, 150–153.
- Tauchmann, S.; Schwaller, J. NSD1: A Lysine Methyltransferase between Developmental Disorders and Cancer. Life 2021, 11, 877.
- Toyokawa, G.; Cho, H.-S.; Masuda, K.; Yamane, Y.; Yoshimatsu, M.; Hayami, S.; Takawa, M.; Iwai, Y.; Daigo, Y.; Tsuchiya, E.; et al. Histone Lysine Methyltransferase Wolf-Hirschhorn Syndrome Candidate 1 Is Involved in Human Carcinogenesis through Regulation of the Wnt Pathway. Neoplasia 2011, 13, 887-IN11.
- Liu, L.; Kimball, S.; Liu, H.; Holowatyj, A.; Yang, Z.-Q. Genetic Alterations of Histone Lysine Methyltransferases and Their Significance in Breast Cancer. Oncotarget 2015, 6, 2466–2482.
- Mahmood, S.F.; Gruel, N.; Nicolle, R.; Chapeaublanc, E.; Delattre, O.; Radvanyi, F.; Bernard-Pierrot, I. PPAPDC1B and WHSC1L1 Are Common Drivers of the 8p11-12 Amplicon, Not Only in Breast Tumors But Also in Pancreatic Adenocarcinomas and Lung Tumors. Am. J. Pathol. 2013, 183, 1634–1644.
- Rooney, C.; Geh, C.; Williams, V.; Heuckmann, J.M.; Menon, R.; Schneider, P.; Al-Kadhimi, K.; Dymond, M.; Smith, N.R.; Baker, D.; et al. Characterization of FGFR1 Locus in sqNSCLC Reveals a Broad and Heterogeneous Amplicon. PLoS ONE 2016, 11, e0149628.
- Stec, I.; Van Ommen, G.-J.B.; Den Dunnen, J.T. WHSC1L1, on Human Chromosome 8p11.2, Closely Resembles WHSC1 and Maps to a Duplicated Region Shared with 4p16.3. Genomics 2001, 76, 5–8.
- Angrand, P.-O.; Apiou, F.; Stewart, A.F.; Dutrillaux, B.; Losson, R.; Chambon, P. NSD3, a New SET Domain-Containing Gene, Maps to 8p12 and Is Amplified in Human Breast Cancer Cell Lines. Genomics 2001, 74, 79–88.
- He, C.; Li, F.; Zhang, J.; Wu, J.; Shi, Y. The Methyltransferase NSD3 Has Chromatin-Binding Motifs, PHD5-C5HCH, That Are Distinct from Other NSD (Nuclear Receptor SET Domain) Family Members in Their Histone H3 Recognition. J. Biol. Chem. 2013, 288, 4692–4703.
- Wu, H.; Zeng, H.; Lam, R.; Tempel, W.; Amaya, M.F.; Xu, C.; Dombrovski, L.; Qiu, W.; Wang, Y.; Min, J. Structural and Histone Binding Ability Characterizations of Human PWWP Domains. PLoS ONE 2011, 6, e18919.
- Rona, G.B.; Almeida, D.S.G.; Pinheiro, A.S.; Eleutherio, E.C.A. The PWWP Domain of the Human Oncogene WHSC1L1/NSD3 Induces a Metabolic Shift toward Fermentation. Oncotarget 2017, 8, 54068–54081.
- Vermeulen, M.; Eberl, H.C.; Matarese, F.; Marks, H.; Denissov, S.; Butter, F.; Lee, K.K.; Olsen, J.V.; Hyman, A.A.; Stunnenberg, H.G.; et al. Quantitative Interaction Proteomics and Genome-Wide Profiling of Epigenetic Histone Marks and Their Readers. Cell 2010, 142, 967–980.
- Stec, I.; Nagl, S.B.; van Ommen, G.-J.B. The PWWP Domain: A Potential Protein-Protein Interaction Domain in Nuclear Proteins Influencing Differentiation? FEBS Lett. 2000, 473, 1–5.
- Jain, K.; Fraser, C.S.; Marunde, M.R.; Parker, M.M.; Sagum, C.; Burg, J.M.; Hall, N.; Popova, I.K.; Rodriguez, K.L.; Vaidya, A.; et al. Characterization of the Plant Homeodomain (PHD) Reader Family for Their Histone Tail Interactions. Epigenetics Chromatin 2020, 13, 3.
- Morishita, M.; Mevius, D.; Di Luccio, E. In Vitro Histone Lysine Methylation by NSD1, NSD2/MMSET/WHSC1 and NSD3/WHSC1L. BMC Struct. Biol. 2014, 14, 1–13.
- Yang, Z.-Q.; Liu, G.; Bollig-Fischer, A.; Giroux, C.N.; Ethier, S.P. Transforming Properties of 8p11-12 Amplified Genes in Human Breast Cancer. Cancer Res. 2010, 70, 8487–8497.
- Shen, C.; Ipsaro, J.J.; Shi, J.; Milazzo, J.P.; Wang, E.; Roe, J.-S.; Suzuki, Y.; Pappin, D.J.; Joshua-Tor, L.; Vakoc, C.R. NSD3-Short Is an Adaptor Protein That Couples BRD4 to the CHD8 Chromatin Remodeler. Mol. Cell 2015, 60, 847–859.
- Irish, J.C.; Mills, J.N.; Turner-Ivey, B.; Wilson, R.C.; Guest, S.T.; Rutkovsky, A.; Dombkowski, A.; Kappler, C.S.; Hardiman, G.; Ethier, S.P. Amplification of WHSC1L1 Regulates Expression and Estrogen-independent Activation of ERα in SUM-44 Breast Cancer Cells and Is Associated with ERα Over-expression in Breast Cancer. Mol. Oncol. 2016, 10, 850–865.
- Kim, S.M.; Kee, H.J.; Eom, G.H.; Choe, N.W.; Kim, J.Y.; Kim, Y.S.; Kim, S.K.; Kook, H.; Kook, H.; Seo, S.B. Characterization of a Novel WHSC1-Associated SET Domain Protein with H3K4 and H3K27 Methyltransferase Activity. Biochem. Biophys. Res. Commun. 2006, 345, 318–323.
- The ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium; Aaltonen, L.A.; Abascal, F.; Abeshouse, A.; Aburatani, H.; Adams, D.J.; Agrawal, N.; Ahn, K.S.; Ahn, S.-M.; Aikata, H.; et al. Pan-Cancer Analysis of Whole Genomes. Nature 2020, 578, 82–93.
- Stephens, P.J.; Greenman, C.D.; Fu, B.; Yang, F.; Bignell, G.R.; Mudie, L.J.; Pleasance, E.D.; Lau, K.W.; Beare, D.; Stebbings, L.A.; et al. Massive Genomic Rearrangement Acquired in a Single Catastrophic Event during Cancer Development. Cell 2011, 144, 27–40.
- Forment, J.V.; Kaidi, A.; Jackson, S.P. Chromothripsis and Cancer: Causes and Consequences of Chromosome Shattering. Nat. Rev. Cancer 2012, 12, 663–670.
- Krupina, K.; Goginashvili, A.; Cleveland, D.W. Scrambling the Genome in Cancer: Causes and Consequences of Complex Chromosome Rearrangements. Nat. Rev. Genet. 2023.
- Voutsadakis, I.A. 8p11.23 Amplification in Breast Cancer: Molecular Characteristics, Prognosis and Targeted Therapy. J. Clin. Med. 2020, 9, 3079.
- Bernard-Pierrot, I.; Gruel, N.; Stransky, N.; Vincent-Salomon, A.; Reyal, F.; Raynal, V.; Vallot, C.; Pierron, G.; Radvanyi, F.; Delattre, O. Characterization of the Recurrent 8p11-12 Amplicon Identifies PPAPDC1B, a Phosphatase Protein, as a New Therapeutic Target in Breast Cancer. Cancer Res. 2008, 68, 7165–7175.
- Turner-Ivey, B.; Smith, E.L.; Rutkovsky, A.C.; Spruill, L.S.; Mills, J.N.; Ethier, S.P. Development of Mammary Hyperplasia, Dysplasia, and Invasive Ductal Carcinoma in Transgenic Mice Expressing the 8p11 Amplicon Oncogene NSD3. Breast Cancer Res. Treat. 2017, 164, 349–358.
- Satpathy, S.; Krug, K.; Jean Beltran, P.M.; Savage, S.R.; Petralia, F.; Kumar-Sinha, C.; Dou, Y.; Reva, B.; Kane, M.H.; Avanessian, S.C.; et al. A Proteogenomic Portrait of Lung Squamous Cell Carcinoma. Cell 2021, 184, 4348–4371.e40.
- Tonon, G.; Wong, K.-K.; Maulik, G.; Brennan, C.; Feng, B.; Zhang, Y.; Khatry, D.B.; Protopopov, A.; You, M.J.; Aguirre, A.J.; et al. High-Resolution Genomic Profiles of Human Lung Cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 9625–9630.
- Chen, Y.; McGee, J.; Chen, X.; Doman, T.N.; Gong, X.; Zhang, Y.; Hamm, N.; Ma, X.; Higgs, R.E.; Bhagwat, S.V.; et al. Identification of Druggable Cancer Driver Genes Amplified across TCGA Datasets. PLoS ONE 2014, 9, e98293.
- Rosati, R.; La Starza, R.; Veronese, A.; Aventin, A.; Schwienbacher, C.; Vallespi, T.; Negrini, M.; Martelli, M.F.; Mecucci, C. NUP98 Is Fused to the NSD3 Gene in Acute Myeloid Leukemia Associated with t(8;11)(P11.2;P15). Blood 2002, 99, 3857–3860.
- Taketani, T.; Taki, T.; Nakamura, H.; Taniwaki, M.; Masuda, J.; Hayashi, Y. NUP98–NSD3 Fusion Gene in Radiation-Associated Myelodysplastic Syndrome with t(8;11)(P11;P15) and Expression Pattern of NSD Family Genes. Cancer Genet. Cytogenet. 2009, 190, 108–112.
- Avenarius, M.R.; Miller, C.R.; Arnold, M.A.; Koo, S.; Roberts, R.; Hobby, M.; Grossman, T.; Moyer, Y.; Wilson, R.K.; Mardis, E.R.; et al. Genetic Characterization of Pediatric Sarcomas by Targeted RNA Sequencing. J. Mol. Diagn. 2020, 22, 1238–1245.
- French, C.A.; Rahman, S.; Walsh, E.M.; Kühnle, S.; Grayson, A.R.; Lemieux, M.E.; Grunfeld, N.; Rubin, B.P.; Antonescu, C.R.; Zhang, S.; et al. NSD3–NUT Fusion Oncoprotein in NUT Midline Carcinoma: Implications for a Novel Oncogenic Mechanism. Cancer Discov. 2014, 4, 928–941.
- Suzuki, S.; Kurabe, N.; Ohnishi, I.; Yasuda, K.; Aoshima, Y.; Naito, M.; Tanioka, F.; Sugimura, H. NSD3-NUT-Expressing Midline Carcinoma of the Lung: First Characterization of Primary Cancer Tissue. Pathol. Res. Pract. 2015, 211, 404–408.
- Agaimy, A.; Tögel, L.; Stoehr, R.; Meidenbauer, N.; Semrau, S.; Hartmann, A.; Mantsopoulos, K. NSD3-NUTM1-Rearranged Carcinoma of the Median Neck/Thyroid Bed Developing after Recent Thyroidectomy for Sclerosing Mucoepidermoid Carcinoma with Eosinophilia: Report of an Extraordinary Case. Virchows Arch. 2021, 479, 1095–1099.
- Chen, M.; Yang, J.; Lv, L.; Li, Y.; Tang, Y.; Liu, W.; Wang, W.; Jiang, L. Comprehensive Genetic Profiling of Six Pulmonary Nuclear Protein in Testis Carcinomas with a Novel Micropapillary Histological Subtype in Two Cases. Hum. Human. Pathol. 2021, 115, 56–66.
- Xiong, Q.; Zhou, Y.; Zhang, S.; Zhang, Y.; Xu, Y.; Yang, Y.; Zhou, C.; Zeng, Z.; Han, J.; Zhu, Q. NSD3, a Member of Nuclear Receptor-Binding SET Domain Family, Is a Potential Prognostic Biomarker for Pancreatic Cancer. Cancer Med. 2023, 12, 10961–10978.
- Sun, Y.; Xie, J.; Cai, S.; Wang, Q.; Feng, Z.; Li, Y.; Lu, J.; Chen, W.; Ye, Z. Elevated Expression of Nuclear Receptor-Binding SET Domain 3 Promotes Pancreatic Cancer Cell Growth. Cell Death Dis. 2021, 12, 913.
- Fang, R.; Barbera, A.J.; Xu, Y.; Rutenberg, M.; Leonor, T.; Bi, Q.; Lan, F.; Mei, P.; Yuan, G.-C.; Lian, C.; et al. Human LSD2/KDM1b/AOF1 Regulates Gene Transcription by Modulating Intragenic H3K4me2 Methylation. Mol. Cell 2010, 39, 222–233.
- Jacques-Fricke, B.T.; Roffers-Agarwal, J.; Hussein, A.O.; Yoder, K.J.; Gearhart, M.D.; Gammill, L.S. Profiling NSD3-Dependent Neural Crest Gene Expression Reveals Known and Novel Candidate Regulatory Factors. Dev. Biol. 2021, 475, 118–130.
- Jacques-Fricke, B.T.; Gammill, L.S. Neural Crest Specification and Migration Independently Require NSD3-Related Lysine Methyltransferase Activity. MBoC 2014, 25, 4174–4186.
- Qin, S.; Min, J. Structure and Function of the Nucleosome-Binding PWWP Domain. Trends Biochem. Sci. 2014, 39, 536–547.
- Kim, S.-M.; Kee, H.-J.; Choe, N.; Kim, J.-Y.; Kook, H.; Kook, H.; Seo, S.-B. The Histone Methyltransferase Activity of WHISTLE Is Important for the Induction of Apoptosis and HDAC1-Mediated Transcriptional Repression. Exp. Cell Res. 2007, 313, 975–983.
- Jeong, G.-Y.; Park, M.K.; Choi, H.-J.; An, H.W.; Park, Y.-U.; Choi, H.-J.; Park, J.; Kim, H.-Y.; Son, T.; Lee, H.; et al. NSD3-Induced Methylation of H3K36 Activates NOTCH Signaling to Drive Breast Tumor Initiation and Metastatic Progression. Cancer Res. 2021, 81, 77–90.
- Jacinto, E.; Loewith, R.; Schmidt, A.; Lin, S.; Rüegg, M.A.; Hall, A.; Hall, M.N. Mammalian TOR Complex 2 Controls the Actin Cytoskeleton and Is Rapamycin Insensitive. Nat. Cell Biol. 2004, 6, 1122–1128.
- Thedieck, K.; Polak, P.; Kim, M.L.; Molle, K.D.; Cohen, A.; Jenö, P.; Arrieumerlou, C.; Hall, M.N. PRAS40 and PRR5-Like Protein Are New mTOR Interactors That Regulate Apoptosis. PLoS ONE 2007, 2, e1217.
- Lu, T.; Jackson, M.W.; Wang, B.; Yang, M.; Chance, M.R.; Miyagi, M.; Gudkov, A.V.; Stark, G.R. Regulation of NF-κB by NSD1/FBXL11-Dependent Reversible Lysine Methylation of P65. Proc. Natl. Acad. Sci. USA 2010, 107, 46–51.
- Yang, P.; Guo, L.; Duan, Z.J.; Tepper, C.G.; Xue, L.; Chen, X.; Kung, H.-J.; Gao, A.C.; Zou, J.X.; Chen, H.-W. Histone Methyltransferase NSD2/MMSET Mediates Constitutive NF-κB Signaling for Cancer Cell Proliferation, Survival, and Tumor Growth via a Feed-Forward Loop. Mol. Cell. Biol. 2012, 32, 3121–3131.
- Saloura, V.; Vougiouklakis, T.; Zewde, M.; Deng, X.; Kiyotani, K.; Park, J.-H.; Matsuo, Y.; Lingen, M.; Suzuki, T.; Dohmae, N.; et al. WHSC1L1-Mediated EGFR Mono-Methylation Enhances the Cytoplasmic and Nuclear Oncogenic Activity of EGFR in Head and Neck Cancer. Sci. Rep. 2017, 7, 40664.
- Lin, R.; Heylbroeck, C.; Pitha, P.M.; Hiscott, J. Virus-Dependent Phosphorylation of the IRF-3 Transcription Factor Regulates Nuclear Translocation, Transactivation Potential, and Proteasome-Mediated Degradation. Mol. Cell. Biol. 1998, 18, 2986–2996.
- Sato, M.; Hata, N.; Asagiri, M.; Nakaya, T.; Taniguchi, T.; Tanaka, N. Positive Feedback Regulation of Type I IFN Genes by the IFN-inducible Transcription Factor IRF-7. FEBS Lett. 1998, 441, 106–110.
- Xu, D.; Liu, S.; Wu, X.; Marti, T.M.; Dorn, P.; Schmid, R.A.; Peng, R.-W.; Shu, Y. Dissecting the Immunological Profiles in NSD3-Amplified LUSC through Integrative Multi-Scale Analyses. Cancers 2022, 14, 4997.
- Saloura, V.; Vougiouklakis, T.; Zewde, M.; Kiyotani, K.; Park, J.-H.; Gao, G.; Karrison, T.; Lingen, M.; Nakamura, Y.; Hamamoto, R. WHSC1L1 Drives Cell Cycle Progression through Transcriptional Regulation of CDC6 and CDK2 in Squamous Cell Carcinoma of the Head and Neck. Oncotarget 2016, 7, 42527–42538.
- Kang, D.; Cho, H.; Toyokawa, G.; Kogure, M.; Yamane, Y.; Iwai, Y.; Hayami, S.; Tsunoda, T.; Field, H.I.; Matsuda, K.; et al. The Histone Methyltransferase Wolf–Hirschhorn Syndrome Candidate 1-like 1 (WHSC1L1) Is Involved in Human Carcinogenesis. Genes. Chromosomes Cancer 2013, 52, 126–139.
- Seo, H.R.; Lee, D.H.; Lee, H.J.; Baek, M.; Bae, S.; Soh, J.W.; Lee, S.J.; Kim, J.; Lee, Y.S. Cyclin G1 Overcomes Radiation-Induced G2 Arrest and Increases Cell Death through Transcriptional Activation of Cyclin B1. Cell Death Differ. 2006, 13, 1475–1484.
- Salem, H.; Rachmin, I.; Yissachar, N.; Cohen, S.; Amiel, A.; Haffner, R.; Lavi, L.; Motro, B. Nek7 Kinase Targeting Leads to Early Mortality, Cytokinesis Disturbance and Polyploidy. Oncogene 2010, 29, 4046–4057.
- Rahman, S.; Sowa, M.E.; Ottinger, M.; Smith, J.A.; Shi, Y.; Harper, J.W.; Howley, P.M. The Brd4 Extraterminal Domain Confers Transcription Activation Independent of pTEFb by Recruiting Multiple Proteins, Including NSD3. Mol. Cell. Biol. 2011, 31, 2641–2652.
- Zhu, H.; Bengsch, F.; Svoronos, N.; Rutkowski, M.R.; Bitler, B.G.; Allegrezza, M.J.; Yokoyama, Y.; Kossenkov, A.V.; Bradner, J.E.; Conejo-Garcia, J.R.; et al. BET Bromodomain Inhibition Promotes Anti-Tumor Immunity by Suppressing PD-L1 Expression. Cell Rep. 2016, 16, 2829–2837.
- Li, Z.; Ivanov, A.A.; Su, R.; Gonzalez-Pecchi, V.; Qi, Q.; Liu, S.; Webber, P.; McMillan, E.; Rusnak, L.; Pham, C.; et al. The OncoPPi Network of Cancer-Focused Protein–Protein Interactions to Inform Biological Insights and Therapeutic Strategies. Nat. Commun. 2017, 8, 14356.