Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Microbiology
Bacteria in the viable but non-culturable (VBNC) state exhibit a remarkable phenomenon: they are unable to grow and form colonies on conventional culture media, yet they remain alive and able to restart their metabolic activity. Cells in this status typically display reduced levels of metabolic activity and undergo significant metabolic alterations, such as reductions in nutrient transport and respiration rates and macromolecular synthesis, and form resistance structures similar to spores. However, a feature that distinguishes the VNBC state is the continuous gene expression within these cells.
- VBNC state
- stress
- beneficial bacteria
- rhizosphere
- latency
- survival
1. Introduction
Beneficial bacteria play a crucial role in maintaining life on our planet. Some realize nitrogen fixation [1], mineral solubilization [2], and greenhouse gas consumption and are thus viewed as gatekeepers preventing excessive methane emissions from escaping the atmosphere [3]. Due to the several processes in which beneficial bacteria participate, they have been used to increase crop production [4]. Some beneficial properties of these bacteria include plant growth promotion [5][6], the control or inhibition of the activity of plant pathogens [7][8], improvements in soil structure, bioaccumulation, or the microbial leaching of inorganics [9], the bioremediation of xenobiotic compounds [10][11], or the production of compounds of industrial interest [12][13].
Certain bacteria that interact with their hosts establish mutually beneficial relationships. Probiotics, for instance, interact with humans to improve overall health. They can eliminate or remove pathogens [14], reinforce the epithelial barrier, and induce the migration of fibroblasts and epithelial cells [15]. In the immune system, probiotics are related to the modulation and activation of intraepithelial lymphocytes, natural killer cells, and macrophages through the induced production of cytokines [16].
On the other hand, there are beneficial bacteria with industrial applications that favor the production of certain foods, prebiotics, and beverages. In the brewing and wine industry, bacteria play a crucial role in the fermentation process. Lactic acid bacteria (LAB) and yeasts are instrumental in this context. LAB catalyze the conversion of dicarboxylic malic acid into monocarboxylic lactic acid and carbon dioxide (malolactic fermentation MLF) and yeasts convert sugars into alcohol (alcoholic fermentation) [17]. During malolactic fermentation by LAB, no free intermediary products are formed, achieving a more palatable wine by reducing the tart taste of malic acid. Additionally, malolactic fermentation reduces the amount of residual nutrients available to support microbial growth, enhances the wine aroma, improves the microbial stability, and reduces the acidity of wine, making the wine more stable before being bottled [18]. Despite the crucial importance and diverse benefits of bacteria across different levels, our comprehension of beneficial bacteria in the VBNC state remains significantly limited. The progress in this field has primarily concentrated on pathogenic bacteria due to their profound impact on human health.
Table 1 shows the beneficial bacteria reported so far to enter the viable but non-culturable state under different conditions. This table displays a variety of conditions that can induce the VBNC state and the respective taxonomic groups to which each described species belongs.
Table 1. Main species of beneficial bacteria that enter the viable but non-culturable state.
| Group of Bacteria | Species | Conditions that Induce VBNC State | References | |
|---|---|---|---|---|
| Proteobecteria | Alphaproteobacteria | Acetobacter aceti | Treatment with SO2 at a concentration of 30 and 50 mg/L | [19] |
| Acetobacter pasteurianus | High acid stress during fermentation | [20] | ||
| Methylosinus sporium | Freeze drying and cryopreservation (liquid nitrogen) |
[3][21] | ||
| Methylosinus trichosporium | ||||
| Methylocystis hirsuta | ||||
| Methylocystis parvus | ||||
| Methylocella tundrae | ||||
| Rhizobium leguminosarum | Cupric sulfate to a concentration of 60 ppm | [22] | ||
| Sinorhizobium meliloti | Incubation at 25 °C in tap water (microcosm-water) Incubation under anoxic conditions in liquid microcosms Incubation in nitrocellulose filters at relative humidity of 22% for three days at 20 °C in the dark |
[23] | ||
| Betaproteobacteria | Cupriavidus metallidurans | Incubation in artificial soil at 30 °C for 12 days, without any C source or H2O | [24] | |
| Gamaproteobacteria | Methylomonas methanica | Lyophilization and cryopreservation (liquid nitrogen) |
[3][21] | |
| Methylosarcina fibrata | ||||
| Methylocaldum gracile | ||||
| Methylomicrobium alcaliphilum | ||||
| Methylococcus capsulatus | ||||
| Microbulbifer aggregans | Incubation in modified artificial seawater (ASW) for 4 h at 30 °C | [25] | ||
| Pseudomonas fluorescens | Incubation in saline solution (NaCl 0.9% w/v) at 37 °C Exposure to benzalkonium chloride (BAC) |
[26] | ||
| Pseudomonas putida KT2440 | Desiccation at 30 °C and 50% relative humidity | [27] | ||
| Vibrio fischeri | Incubation at 22 °C in nutrient-limited artificial seawater (ASW) |
[28] | ||
| Terrabacteria | Actinobacteria | Bifidobacterium animalis subsp. lactis |
|
[29] |
| Bifidobacterium longum | Storage in fermented foods | [30] | ||
| Arthrobacter albidus, reclassified as Sinomonas albida |
Absence of resuscitation promoting factor (Rpf) protein in the culture medium |
[31][32] | ||
| Firmicutes | Bacillus coagulans | Incubation at pH 2 for 24 h and subsequent incubation at 140 °C for 5 min | [33] | |
| Lactiplantibacillus plantarum | Treatment for 30 min at 100 °C or with 1 mol/L HCl Incubation in beer at 0 °C temperature |
[34] | ||
| Oenococcus oeni | Sulfur dioxide and histidine decarboxylase activity in wines | [19] | ||
The phylogenetic relationships between different beneficial bacterial species reported to enter the VBNC state were analyzed (Figure 2). The sequences were compared with the data available in the National Center for Biotechnology Information (NCBI) database. To evaluate viable but unculturable strains, phylogenetic trees were constructed by the neighbor-joining method [35] using the Clustal X 2.1, BioEdit 7.7, and Mega 4 ©1993–2011 software. A bootstrap confidence analysis was applied on 1000 replicates to determine the reliability of the topology obtained [36]. The phylogenetic tree showed that these bacteria are very diverse. The furthest phylogenetic group corresponds to Methylosinus, Methylocystis, and Methylocella. The genera Rhizobium, Sinorhizobium, and Acetobacter are groups that are phylogenetically closer to each other. Methylococcus, Methylocaldum, Methylomicrobium, Methylotuvimicrobium, Methylosarcina, and Methylomonas are phylogenetically close to Vibrio, Microbulbifer, and Pseudomonas. Another group that can be identified is formed by Arthrobacter, Bifidobacterium, Bacillus, Lactobacillus, and Oenococcus, with some strains being phylogenetically closely related and others being very distant species. When analyzing the phylogenetic relationships, it can be observed that the viable but non-culturable state is not exclusive to any taxonomic group or group of species, and it is a widely distributed strategy in phylogenetically close and distant species.
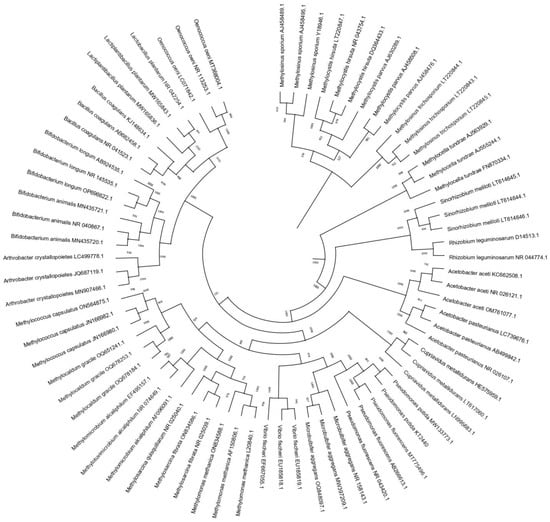
Figure 2. Phylogenetic tree of beneficial bacterial species entering the VBNC state. Phylogenetic trees were constructed by the neighbor-joining method [35] using Clustal X 2.1, BioEdit 7.7, and Mega 4 ©1993–2011 software. A bootstrap confidence analysis was applied on 1000 replicates to determine the reliability of the topology obtained.
The beneficial species in which the viable but non-culturable state has been reported are described below, highlighting their main applications in different areas, such as biotechnology, agro-biotechnology, health, and industrial applications.
2. Alphaproteobacteria and the VBNC State
Within the group of Alphaproteobacteria that enter the VBNC state are organisms belonging to the genera Acetobacter, Methylosinus, Methylocistis, Methylocella, Rhizobium, and Sinorhizobium (Table 1).
Acetobacter aceti and Acetobacter pasteurianus have a great relevance in vinegar production, since they can transform ethanol into acetic acid through oxidative fermentation [37]. The VBNC state in A. aceti has been documented in wine production [19], and A. pasteurianus enters the VBNC state under high acid stress generated during the fermentation process [20]. Studies are still lacking that support the idea that A. aceti and A. pasteurianus, under a non-culturable state, can follow the fermentation process, which would help to increase production. The ability of these species to persist under adverse conditions represents a challenge in developing novel ways to improve industrial yields, enhance wine quality, and drive vinegar production on a larger scale.
The methylotrophic bacteria Methylosinus sporium, Methylosinus trichosporium, Methylocystis hirsuta, Methylocystis parvus, and Methylocella tundrae are methanotrophic microorganisms, also called methane-oxidizing bacteria (MOB), capable of generating energy through the oxidation of methane gas [38]. MOB have different biotechnological applications, mainly the biological mitigation of the methane greenhouse gas, the production of high-value products from methane, and the bioremediation of pollutants [39][40]. Because of the ecological importance of these microorganisms, it is important to understand how different conditions can affect methane consumption. One of the conditions that induces the VBNC state in MOB is freeze-drying and cryopreservation (Table 1). This represents a challenge to researchers, because culturable cells are needed for beneficial application. Most studies focus on how to avoid the loss of cultivability, but it is likely that MOB can return to a culturable state when in an environment with methane gas. This has been observed with different pathogen species of bacteria that, when found in favorable conditions, leave the non-culturable state [41]. Research with this group of bacteria in the VBNC state is scarce, representing a challenge in understanding this survival strategy. In the future, this could stimulate these organisms to consume methane, aiding in the degradation of pollutants and the production of high-value biomass.
Other species of Alphaproteobacteria are Sinorhizobium meliloti and Rhizobium leguminosarum, found mainly in the soil, which have acquired, by horizontal gene transfer, the ability to associate in symbiosis with leguminous plant roots. The association of rhizobia and legumes occurs through a complex signaling process that generates nodules, organs specialized in the fixation of atmospheric nitrogen [42]. Biological nitrogen fixation is a vital process in agriculture, allowing the production of nitrogen through legume–rhizobium symbiosis, which contributes to increased nitrogen levels in the soil, resulting in increased plant growth [43]. In addition to the ability to fix nitrogen, R. leguminosarum isolated from fava bean root nodules was shown to generate an induced systemic response against infection by bean yellow mosaic virus [44]. The VBNC state in this bacterium is related to the presence of cupric sulfate (Table 1). For S. meliloti, the factors that induce the VBNC state are temperatures of 20 °C to 25 °C, incubation under anoxic conditions, and incubation in nitrocellulose filters at low relative humidity (Table 1). It is interesting that, under these conditions, the bacteria can persist. This could explain their survival capacity under this type of stress in the environment. The importance of rhizobia for agriculture highlights the need to learn more about their beneficial functions in the VBNC state, because it is unknown whether, under a non-culturable state, bacteria can continue performing nitrogen fixation, generate an induced systemic response, or even develop nodules.
3. Betaproteobacteria and the VBNC State
In the group of Betaproteobacteria, the only beneficial bacterium where the VBNC state has been documented is Cupriavidus metallidurans. This bacterium is metallophilic, found in environments containing high concentrations of heavy metals and industrial wastes rich in toxic heavy metals, often mixed with recalcitrant organic compounds and hydrocarbons [45]. It is an ideal bacterium for bioaugmentation purposes in environmental applications due to its strong resistance to environmental stress factors and its adaptation capacity [46]. Another specific application is its ability to convert gold chloride into 24-carat gold in one week [47]. These observations suggest that bacteria actively contribute to the formation of gold grains in surface environments [48]. With this bacterium, the conditions that can induce its loss of cultivability have been studied, as well as the conditions that can return it to a cultivable state. The addition of water and gluconate is sufficient for C. metallidurans to be culturable in 24 h [24]. Due to its interesting applications and ability to persist in toxic environments, it is an excellent model to study the mechanisms of coping with heavy metal stress. This unique ability to metabolize toxic substances and enter the VBNC state without water and carbon sources could help us to understand how the origin of life occurred.
4. Gammaproteobacteria and the VBNC State
One of the groups with the most beneficial species in which the VBNC status has been demonstrated is Gammaproteobacteria. Methylomonas methanica, Methylosarcina fibrate, Methylocaldum gracile, Methylomicrobium alcaliphilum, and Methylococcus capsulatus are methylotrophs. This type of bacteria uses methanol and methane as the only carbon source [49]. Methylotrophs can inhabit soil, water, and plants, powering the carbon cycle [50]. When M. fibrate was co-inoculated with species of Methylomonas and Cupriavidus taiwanensis LMG 19424, its growth was highly stimulated [51]. Moreover, methane oxidation was elevated when methanotrophs interacted with algae and moss [52]. Cryopreservation is one method that induces the VBNC state in methanotrophs (Table 1). The study of the VBNC state in methanotrophs is essential due to its role in the carbon cycle and therefore in the planet’s life. It is relevant to consider that interaction with other microorganisms or with plants could be an alternative strategy for these microorganisms to return to the cultivable state, being able to continue exerting their beneficial effects.
Microbulbifer aggregans, another Gammaproteobacteria species, is a halophilic, Gram-negative bacterium isolated from sediment in the Matang mangrove forest, Malaysia [53]; its importance is centered on its capability to reduce sulfur due to the presence of genes involved in this process [25]. Sulfur is an essential element for life and is present in amino acids, proteins, enzymes, vitamins, and other biological molecules [54]. M. aggregans is a rarely explored bacterium. Recently, it was observed that non-culturable cells exhibited a change in cell shape from rod to coccus, and the genes responsible for sulfate reduction were upregulated in the VBNC state [25]. These findings demonstrate this species’ importance in the environment in which it lives, because the effective reduction of sulfur in the non-culturable state may occur in response to changes in the sulfur concentration in the environment, playing a relevant role in the sulfur cycle in marine environments.
Within the Gammaproteobacteria, there is also the genus Pseudomonas. The species of this genus have a great capacity to use different nutrients as carbon sources, which explains their ubiquity. Their enzymatic activity makes them an important group of microorganisms responsible for the aerobic degradation of many compounds in different ecosystems [55]. Some species of Pseudomonas can promote plant growth by suppressing pathogenic microorganisms, synthesizing growth-stimulating plant hormones, and promoting increased plant disease resistance [56]. Pseudomonas fluorescens and Pseudomonas putida KT2440 are bacteria able to colonize plants’ roots and promote their growth. Some strains of P. fluorescens have been shown to degrade a variety of organic compounds, thus being important in bioremediation [57]. This bacterium is used for biocontrol to protect plants against soilborne fungal pathogens. One mechanism for the biocontrol by P. fluorescens is the ability to produce antibiotics [58]. It also stimulates induced systemic resistance (ISR) [59] and the production of volatile compounds [60].
On the other hand, P. putida KT2440 is a bacterium capable of using different aromatic compounds as a carbon source [61]. Metabolizing xenobiotic compounds, it can colonize the roots of plants such as corn, wheat, strawberry, sugarcane, and spinach [62] and is capable of promoting the growth and health of plants [63][64]. P. putida KT2440 has been used in various bacterial formulations to enhance plant growth [64][65].
The potential of P. putida KT2440 and P. fluorescens to exert beneficial effects can be affected by their exposure to different stresses that induce the VBNC state, such as saline environments for P. fluorescens and desiccation for P. putida KT2440 (Table 1). However, it has been shown that plant–bacteria interaction is a mechanism that allows these bacteria return to the cultivable state. Therefore, their use in the formulation of stable bacterial inoculants that can stimulate plant growth after rehydration [27], or the development of formulations to perform biological control [66], could be an alternative to reduce the overuse of nitrogen fertilizers and pesticides, decreasing the damage caused by these chemical products [67].
Vibrio fischeri, also belonging to the Gammaproteobacteria, is a luminous marine bacterium that lives freely or in symbiosis with different species of fish and squid [68]. The most studied interaction in this microorganism is the symbiosis with the Hawaiian squid, Euprymna scolopes, inducing bioluminescence that the squid uses to avoid predation during nocturnal activity [69]. In the squid–Vibrio symbiosis, bacteria are found in a ventral tissue called the lumen organ. The relationship between V. fischeri and E. scolopes is characterized by daily rhythmic cycles that control the population dynamics of bacteria. At night, when squid take feed, the light organ fills with bioluminescent V. fischeri. At dawn, the squid bury themselves in the sand and ventilate approximately 90% of the bacterial population to the environment; the remaining bacteria repopulate the crypts and are ready to produce light at dusk [70]. This example of symbiosis shows beneficial microorganisms’ important role in their hosts’ health and activities. The role of the VBNC state in V. fischeri living freely in the marine environment or in symbiosis is still unknown. It has only been discovered that non-culturable cells lose their luminescence in response to fluctuations in salinity [28]. The loss of luminescence could have relevance in the diagnosis of marine environments, allowing the determination of alterations in salinity, temperature, and the concentrations of nutrients related to the number of luminescent bacteria of V. fischeri, with the interpretation that the fewer luminescent bacteria observed, the greater the disturbance present in the marine environment.
5. Actinobacteria and the VBNC State
The Actinobacteria group includes Sinomonas albida, formerly known as Arthrobacter albidus, isolated from a seep substrate composed of volcanic rock from Niigata, Japan [71]. Among the main applications of this microorganism, it has a demonstrated ability to exit the VBNC state in the presence of the RpF protein, along with its efficient flocculant activity [31]. This flocculent activity represents a possible application in wastewater treatment and residual sludge dewatering. At present, little is known about this species, which represents an alternative and challenge in terms of taking advantage of the biotechnological properties that S. albida can provide.
The presence of bacteria in the human digestive tract offers various properties related to health, such as the regulation of intestinal microbial homeostasis, the inhibition of pathogenic bacteria, the modulation of the immune response, anticancer effects, the production of bacteriocins, or the bioconversion of diet components into bioactive compounds [72]. Recently, the production and consumption of products with beneficial strains for human health has increased considerably [73]. Bifidobacterium longum and Bifidobacterium animalis subsp. lactis are multifunctional probiotic Actinobacteria with clinical effectiveness, including immunomodulatory, anti-inflammatory, antimutagenic, and anticancer properties and alleviating gastrointestinal diseases [73]. In both species, it has been observed that their storage at low temperatures in fermented foods can cause a loss of cultivability (Table 1). In this respect, the determination of the viability and activity of probiotic bacteria is of great economic, regulatory, and technological importance to ensure that fermented products or formulations with probiotics carry bacteria capable of exerting their beneficial effects.
6. Firmicutes and the VBNC State
Bacillus coagulans is a species of Firmicutes that is considered probiotic-safe. This bacterium has the ability to endure high temperatures and it has developed genetic stability through several years of commercial production [74]. Its main benefits for human health include the modulation of gastrointestinal disorders, immune system stimulation, and lowering cholesterol [75]. This bacterium can form endospores and survive for decades in unfavorable environmental conditions. It was previously thought that its high persistence was due only to its ability to form spores; however, it has been observed that B. coagulans can enter the VBNC state as a strategy to face adverse conditions [33]. Little is known about the return to cultivability when humans consume these beneficial microorganisms. Some studies have shown that bacteria secrete certain specific proteins to exit the latency state [76], which is a possibility in the case of B. coagulans.
Lactiplantibacillus plantarum is a Firmicutes species widely distributed in various environments, such as the gastrointestinal, vaginal, and urogenital tracts, and in dairy products, vegetables, meat, hay, and wine. This ability to adapt to different conditions demonstrates its metabolic diversity [77], having potential for various applications. The main applications of L. plantarum include the fermentation of foods such as cheese, kefir, sauerkraut, fermented meat products, fermented vegetables, and beverages [78]. It has been reported that L. plantarum produces antimicrobial substances such as plantaricin [79] and can remove microcystins, the main toxins produced by cyanobacteria [80], which makes it an alternative food preservative or a compound to fight infections. Studies of the VBNC state in L. plantarum have reported that it can remain latent during beer storage, which contributes to the deterioration of this beverage [34]. Another study showed that L. plantarum in the VBNC state can inhibit microcystins, which contributes to the preservation of fermented foods [80]. It is interesting to observe that the VBNC state in this species can be detrimental for beer production but beneficial for food preservation, reflecting the versatility of non-culturable L. plantarum to continue performing its functions. This suggests that when used as a probiotic to improve human health, it can provide its benefits without being culturable. In short, it is a microorganism that should be studied in greater depth, particularly regarding what happens during the VBNC state, to take advantage of its diversity of applications.
In the production of alcoholic beverages, the involvement of microorganisms is crucial for successful fermentation. A significant challenge in the brewing and winemaking industry is the spoilage of wine and beer by lactic acid bacteria, which have the ability to enter the VBNC state [34]. Oenococcus oeni is a Firmicutes bacterium, belonging to the lactic acid bacteria group, adapted to the stressful environment of wine. It is widely used as a starter microorganism to carry out malolactic fermentation (MLF), where L-malate is converted into L-lactate [81]. The use of starter cultures of O. oeni remains difficult in some wine regions, due to the hostile environment created by the low pH and the presence of SO2 and ethanol [82]. The ability of O. oeni to respond these stress conditions has great relevance in terms of increasing wine production at a lower cost. Thus far, knowledge of the VBNC status of O. oeni is limited. It is known that the presence of sulfur dioxide induces the loss of cultivability, and the addition of arginine to the medium allows a return to the cultivable state [81]. According to findings with O. oeni, although the bacteria are in the VBNC state, they may still be capable of carrying out malolactic fermentation, although perhaps not as efficiently as their cultivable counterparts. When cells are present in a rich environment and arginine is added, the bacteria return to the cultivable state, increasing the fermentation efficiency. The generation of new knowledge requires changing the paradigm and legislation that require all microorganisms used in food production to be culturable. It is feasible that the use of non-culturable cells could decrease production costs because expensive infrastructure would not be required to preserve the bacteria, since they have all the necessary equipment to preserve themselves.
The VBNC state could be an important reservoir of beneficial bacterial species, as this state constitutes a survival strategy in response to harsh environmental conditions. The capability of bacteria to enter the VBNC state in response to stress could have important biotechnological applications. It is possible consider that the VBNC state may reduce negative selection and regulate microbial dominance in soil, as the rhizosphere, where plants could induce a selective bacterial revival by releasing selected organic compounds, directly influences the diversity present in these environments. In the same way, the hosts themselves could be exerting a selection bias by releasing compounds that determine the viability of certain bacteria necessary in a given physiological state.
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms12010039
References
- Dobereiner, J.; Urquiaga, S. Alternatives for nitrogen nutrition of crops in tropical agriculture. Fertil. Res. 1994, 42, 339–346.
- Crowley, D.E.; Wang, Y.C.; Reid, C.P.P.; Szaniszlo, P.J. Mechanisms of iron acquisition from siderophores by microorganisms and plants. In Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 1991; pp. 179–198. ISBN 978-94-010-5455-3.
- Hoefman, S.; Van Hoorde, K.; Boon, N.; Vandamme, P.; De Vos, P.; Heylen, K. Survival or revival: Long-term preservation induces a reversible viable but non-culturable state in methane-oxidizing bacteria. PLoS ONE 2012, 7, e34196.
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570.
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556.
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598.
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375.
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55.
- Brierley, C.L. Microbiological mining. Sci. Am. 1982, 247, 44–53.
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis Strain SJ-101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997.
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New Advances in plant growth-promoting rhizobacteria for bioremediation. Environ. Int. 2007, 33, 406–413.
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78.
- Chemier, J.A.; Fowler, Z.L.; Koffas, M.A.G. Trends in microbial synthesis of natural products and biofuels. In Advances in Enzymology—And Related Areas of Molecular Biology; Toone, E.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; Volume 76, pp. 151–217. ISBN 978-0-470-39288-1.
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538.
- Liévin-Le Moal, V.; Servin, A.L. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: Mucins, antimicrobial peptides, and microbiota. Clin. Microbiol. Rev. 2006, 19, 315–337.
- Pagnini, C.; Saeed, R.; Bamias, G.; Arseneau, K.O.; Pizarro, T.T.; Cominelli, F. Probiotics promote gut health through stimulation of epithelial innate immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 454–459.
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Konings, W.N., Kuipers, O.P., Huis Veld, J.H.J., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 317–331. ISBN 978-90-481-5312-1.
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic acid bacteria in wine: Technological advances and evaluation of their functional role. Front. Microbiol. 2021, 11, 612118.
- Millet, V.; Lonvaud-Funel, A. The viable but non-culturable state of wine micro-organisms during storage. Lett. Appl. Microbiol. 2000, 30, 136–141.
- Xia, K.; Han, C.; Xu, J.; Liang, X. Toxin-antitoxin HicAB regulates the formation of persister cells responsible for the acid stress resistance in Acetobacter pasteurianus. Appl. Microbiol. Biotechnol. 2021, 105, 725–739.
- Dong, K.; Pan, H.; Yang, D.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Induction, detection, formation, and resuscitation of viable but non-culturable state microorganisms. Comp. Rev. Food Sci. Food Safe 2020, 19, 149–183.
- Alexander, E.; Pham, D.; Steck, T.R. The Viable-but-nonculturable condition is induced by copper in Agrobacterium tumefaciens and Rhizobium leguminosarum. Appl. Environ. Microbiol. 1999, 65, 3754–3756.
- Vriezen, J.A.; De Bruijn, F.J.; Nüsslein, K.R. Desiccation Induces viable but non-culturable cells in Sinorhizobium meliloti 1021. AMB Expr. 2012, 2, 6.
- Giagnoni, L.; Arenella, M.; Galardi, E.; Nannipieri, P.; Renella, G. Bacterial culturability and the Viable but Non-Culturable (VBNC) state studied by a proteomic approach using an artificial soil. Soil Biol. Biochem. 2018, 118, 51–58.
- Furusawa, G.; Diyana, T.; Lau, N.-S. Metabolic strategies of dormancy of a marine bacterium Microbulbifer aggregans CCB-MM1: Its alternative electron transfer chain and sulfate-reducing pathway. Genomics 2022, 114, 443–455.
- Arana, I.; Muela, A.; Orruño, M.; Seco, C.; Garaizabal, I.; Barcina, I. Effect of temperature and starvation upon survival strategies of Pseudomonas fluorescens CHA0: Comparison with Escherichia coli. FEMS Microbiol. Ecol. 2010, 74, 500–509.
- Pazos-Rojas, L.A.; Muñoz-Arenas, L.C.; Rodríguez-Andrade, O.; López-Cruz, L.E.; López-Ortega, O.; Lopes-Olivares, F.; Luna-Suarez, S.; Baez, A.; Morales-García, Y.E.; Quintero-Hernández, V.; et al. Desiccation-Induced viable but nonculturable state in Pseudomonas putida KT2440, a Survival Strategy. PLoS ONE 2019, 14, e0219554.
- Ramaiah, N.; Ravel, J.; Straube, W.L.; Hill, R.T.; Colwell, R.R. Entry of Vibrio harveyi and Vibrio fischeri into the viable but nonculturable state. J. Appl. Microbiol. 2002, 93, 108–116.
- Verruck, S.; Silva, K.J.; De Oliveira Santeli, H.; Scariot, M.C.; Venturelli, G.L.; Prudencio, E.S.; Arisi, A.C.M. Bifidobacterium animalis ssp. lactis BB-12 enumeration by quantitative PCR assay in microcapsules with full-fat goat milk and inulin-type fructans. Food Res. Int. 2020, 133, 109131.
- Lahtinen, S.J.; Ahokoski, H.; Reinikainen, J.P.; Gueimonde, M.; Nurmi, J.; Ouwehand, A.C.; Salminen, S.J. Degradation of 16S rRNA and Attributes of viability of viable but nonculturable probiotic bacteria. Lett. Appl. Microbiol. 2008, 46, 693–698.
- Su, X.; Shen, X.; Ding, L.; Yokota, A. Study on the flocculability of the Arthrobacter sp., an actinomycete resuscitated from the VBNC state. World J. Microbiol. Biotechnol. 2012, 28, 91–97.
- Zhou, Y.; Chen, X.; Zhang, Y.; Wang, W.; Xu, J. Description of Sinomonas soli sp. nov., reclassification of Arthrobacter echigonensis and Arthrobacter albidus (Ding et al. 2009) as Sinomonas echigonensis comb. nov. and Sinomonas albida comb. nov., respectively, and emended description of the genus Sinomonas. Int. J. Syst. Evol. Microbiol. 2012, 62, 764–769.
- Majeed, M.; Majeed, S.; Nagabhushanam, K.; Punnapuzha, A.; Philip, S.; Mundkur, L. Rapid Assessment of Viable but non-culturable Bacillus coagulans MTCC 5856 in commercial formulations using flow cytometry. PLoS ONE 2018, 13, e0192836.
- Liu, J.; Li, L.; Li, B.; Peters, B.M.; Deng, Y.; Xu, Z.; Shirtliff, M.E. Study on spoilage capability and VBNC state formation and recovery of Lactobacillus plantarum. Microb. Pathog. 2017, 110, 257–261.
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425.
- Felsenstein, J. Phylogenies and the comparative method. Am. Soc. Nat. 1985, 125, 1–15.
- Andrés-Barrao, C.; Saad, M.M.; Chappuis, M.-L.; Boffa, M.; Perret, X.; Ortega Pérez, R.; Barja, F. Proteome analysis of Acetobacter pasteurianus during Acetic acid fermentation. J. Proteom. 2012, 75, 1701–1717.
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010, 34, 496–531.
- Jiang, H.; Chen, Y.; Jiang, P.; Zhang, C.; Smith, T.J.; Murrell, J.C.; Xing, X.-H. Methanotrophs: Multifunctional bacteria with promising applications in environmental bioengineering. Biochem. Eng. J. 2010, 49, 277–288.
- Wendlandt, K.-D.; Stottmeister, U.; Helm, J.; Soltmann, B.; Jechorek, M.; Beck, M. The Potential of methane-oxidizing bacteria for applications in environmental biotechnology. Eng. Life Sci. 2010, 10, 87–102.
- Oliver, J.D. Recent Findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425.
- Coba De La Peña, T.; Fedorova, E.; Pueyo, J.J.; Lucas, M.M. The symbiosome: Legume and rhizobia co-evolution toward a nitrogen-fixing organelle? Front. Plant Sci. 2018, 8, 2229.
- Moawad, H.; Abd el-Rahim, W.M.; Abd el-Aleem, D.; Abo Sedera, S.A. Persistence of two Rhizobium etli inoculant strains in clay and silty loam soils. J. Basic Microbiol. 2005, 45, 438–446.
- Abdelkhalek, A.; Yassin, Y.; Abdel-Megeed, A.; Abd-Elsalam, K.; Moawad, H.; Behiry, S. Rhizobium leguminosarum bv. viciae-mediated silver nanoparticles for controlling Bean Yellow Mosaic Virus (BYMV) infection in faba bean plants. Plants 2022, 12, 45.
- Mergeay, M.; Monchy, S.; Vallaeys, T.; Auquier, V.; Benotmane, A.; Bertin, P.; Taghavi, S.; Dunn, J.; Van Der Lelie, D.; Wattiez, R. Ralstonia metallidurans, a bacterium specifically adapted to toxic metals: Towards a catalogue of metal-responsive genes. FEMS Microbiol. Rev. 2003, 27, 385–410.
- Diels, L.; Van Roy, S.; Taghavi, S.; Van Houdt, R. From industrial sites to environmental applications with Cupriavidus metallidurans. Antonie Leeuwenhoek 2009, 96, 247–258.
- Lal, D.; Nayyar, N.; Kohli, P.; Lal, R. Cupriavidus metallidurans: A modern alchemist. Indian J. Microbiol. 2013, 53, 114–115.
- Reith, F.; Etschmann, B.; Grosse, C.; Moors, H.; Benotmane, M.A.; Monsieurs, P.; Grass, G.; Doonan, C.; Vogt, S.; Lai, B.; et al. Mechanisms of gold biomineralization in the bacterium Cupriavidus metallidurans. Proc. Natl. Acad. Sci. USA 2009, 106, 17757–17762.
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of methylotrophs with plants and other heterotrophic bacteria. Microorganisms 2015, 3, 137–151.
- Semrau, J.D.; DiSpirito, A.A.; Vuilleumier, S. Facultative methanotrophy: False leads, true results, and suggestions for future research: Facultative methanotrophy. FEMS Microbiol. Lett. 2011, 323, 1–12.
- Stock, M.; Hoefman, S.; Kerckhof, F.-M.; Boon, N.; De Vos, P.; De Baets, B.; Heylen, K.; Waegeman, W. Exploration and prediction of interactions between methanotrophs and heterotrophs. Res. Microbiol. 2013, 164, 1045–1054.
- Liebner, S.; Zeyer, J.; Wagner, D.; Schubert, C.; Pfeiffer, E.-M.; Knoblauch, C. Methane oxidation associated with submerged brown mosses reduces methane emissions from siberian polygonal tundra: Moss-associated methane oxidation. J. Ecol. 2011, 99, 914–922.
- Moh, T.H.; Furusawa, G.; Amirul, A.A.-A. Microbulbifer aggregans sp. nov., isolated from estuarine sediment from a mangrove forest. Int. J. Syst. Evol. Microbiol. 2017, 67, 4089–4094.
- Komarnisky, L.A.; Christopherson, R.J.; Basu, T.K. Sulfur: Its clinical and toxicologic aspects. Nutrition 2003, 19, 54–61.
- Spiers, A.J.; Buckling, A.; Rainey, P.B. The causes of Pseudomonas diversity. Microbiology 2000, 146, 2345–2350.
- Preston, G.M. Plant Perceptions of plant growth-promoting Pseudomonas. Phil. Trans. R. Soc. Lond. B 2004, 359, 907–918.
- Bunker, S.T.; Bates, T.C.; Oliver, J.D. Effects of temperature on detection of plasmid or chromosomally encoded gfp and lux labeled Pseudomonas fluorescens in Soil. Environ. Biosaf. Res. 2004, 3, 83–90.
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Efficacy of Pseudomonas fluorescens for control of mucor rot of apple during commercial storage and potential modes of action. Can. J. Microbiol. 2018, 64, 420–431.
- Van Der Ent, S.; Van Wees, S.C.M.; Pieterse, C.M.J. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 2009, 70, 1581–1588.
- Wang, Z.; Zhong, T.; Chen, K.; Du, M.; Chen, G.; Chen, X.; Wang, K.; Zalán, Z.; Takács, K.; Kan, J. Antifungal activity of volatile organic compounds produced by Pseudomonas fluorescens ZX and potential biocontrol of blue mold decay on postharvest citrus. Food Control 2021, 120, 107499.
- Ramos-González, M.I.; Ramos-Díaz, M.A.; Ramos, J.L. Chromosomal gene capture mediated by the Pseudomonas putida TOL catabolic plasmid. J. Bacteriol. 1994, 176, 4635–4641.
- Espinosa-Urgel, M.; Kolter, R.; Ramos, J.-L. Root colonization by Pseudomonas putida: Love at first sight. Microbiology 2002, 148, 341–343.
- Matilla, M.A.; Ramos, J.L.; Bakker, P.A.H.M.; Doornbos, R.; Badri, D.V.; Vivanco, J.M.; Ramos-González, M.I. Pseudomonas putida KT2440 causes induced systemic resistance and changes in Arabidopsis root exudation. Environ. Microbiol. Rep. 2010, 2, 381–388.
- Molina-Romero, D.; Morales-García, Y.-E.; Hernández-Tenorio, A.-L.; Netzahuatl-Muñoz, A.-R. Pseudomonas putida estimula el crecimiento de maíz en función de la temperatura. Rev. Iberoam. Cienc. 2017, 4, 80–88.
- Morales-García, Y.E.; Baez, A.; Quintero-Hernández, V.; Molina-Romero, D.; Rivera-Urbalejo, A.P.; Pazos-Rojas, L.A.; Muñoz-Rojas, J. Bacterial Mixtures, the future generation of inoculants for sustainable crop production. In Field Crops: Sustainable Management by PGPR, Sustainable Development and Biodiversity; Springer Nature: Cham, Switzerland, 2019; Volume 23, pp. 11–44. ISBN 978-3-030-30925-1.
- Daranas, N.; Bonaterra, A.; Francés, J.; Cabrefiga, J.; Montesinos, E.; Badosa, E. Monitoring Viable cells of the biological control agent Lactobacillus plantarum PM411 in aerial plant surfaces by means of a strain-specific viability quantitative PCR method. Appl. Environ. Microbiol. 2018, 84, e00107-18.
- Pazos-Rojas, L.A.; Marín-Cevada, V.; García, Y.E.M.; Baez, A.; Villalobos-López, M.A.; Pérez-Santos, M. Uso de microorganismos benéficos para reducir los daños causados por la revolución verde. Rev. Iberoam. Cienc. 2016, 3, 72–85.
- Meighen, E.A. Molecular biology of bacterial bioluminescence. Microbiol. Rev. 1991, 55, 123–142.
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the Hawaiian Bobtail squid, Euprymna scolopes berry (Mollusca: Cephalopoda). Mar. Biol. 2004, 144, 1151–1155.
- Lee, K.-H.; Ruby, E.G. Effect of the squid host on the abundance and distribution of symbiotic Vibrio fischeri in Nature. Appl. Environ. Microbiol. 1994, 60, 1565–1571.
- Ding, L.; Hirose, T.; Yokota, A. Four novel Arthrobacter species Isolated from filtration substrate. Int. J. Syst. Evol. Microbiol. 2009, 59, 856–862.
- Soomro, A.H.; Masud, T.; Anwaar, K. Role of Lactic Acid Bacteria (LAB) in food preservation and human health—A Review. Pak. J. Nutr. 2001, 1, 20–24.
- Mousavi Khaneghah, A.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in Hosts and foods: A review. Trends Food Sci. Technol. 2020, 95, 205–218.
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Eshuis-de Ruiter, T.; Booij-Veurink, J.; De Vries, Y.P.; Ali, F. Evaluation of genetic and phenotypic consistency of Bacillus coagulans MTCC 5856: A commercial probiotic strain. World J. Microbiol. Biotechnol. 2016, 32, 60.
- Saw, C.-Y.; Chang, T.-J.; Chen, P.-Y.; Dai, F.-J.; Lau, Y.-Q.; Chen, T.-Y.; Chau, C.-F. Presence of Bacillus coagulans spores and vegetative cells in rat intestine and feces and their physiological effects. Biosci. Biotechnol. Biochem. 2019, 83, 2327–2333.
- Gupta, R.K.; Srivastava, R. Resuscitation Promoting Factors: A Family of microbial proteins in survival and resuscitation of dormant mycobacteria. Indian J. Microbiol. 2012, 52, 114–121.
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and its probiotic and food potentialities. Probiotics Antimicro. Prot. 2017, 9, 111–122.
- Todorov, S.D.; Franco, B.D.G.D.M. Lactobacillus plantarum: Characterization of the species and application in food production. Food Rev. Int. 2010, 26, 205–229.
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.V.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.-M.; Haertlé, T.; Franco, B.D.G.M. Characterization of a two-peptide plantaricin produced by Lactobacillus plantarum MBSa4 isolated from brazilian salami. Food Control 2016, 60, 103–112.
- Surono, I.S.; Collado, M.C.; Salminen, S.; Meriluoto, J. Effect of glucose and incubation temperature on metabolically active Lactobacillus plantarum from Dadih in removing microcystin-LR. Food Chem. Toxicol. 2008, 46, 502–507.
- Tonon, T.; Lonvaud-Funel, A. Metabolism of arginine and its positive effect on growth and revival of Oenococcus oeni: Degradation of arginine by Oenococcus oeni. J. Appl. Microbiol. 2000, 89, 526–531.
- Bech-Terkilsen, S.; Westman, J.O.; Swiegers, J.H.; Siegumfeldt, H. Oenococcus oeni, a species born and moulded in wine: A critical review of the stress impacts of wine and the physiological responses. Aust. J. Grape Wine Res. 2020, 26, 188–206.
This entry is offline, you can click here to edit this entry!