RNA and single-stranded DNA (ssDNA) phages make up an understudied subset of bacteriophages that have been rapidly expanding in the last decade thanks to advancements in metaviromics. Antibiotics play a crucial role in treating bacterial infections, but their overuse and misuse have led to the emergence of antimicrobial resistance (AMR), resulting in reduced drug efficiency and persistent infections. Given the rapid evolution of AMR and the challenges of developing novel drugs, exploring alternative strategies becomes imperative. Phage therapy, utilizing phage-derived products and both natural and engineered phages for infection treatment, along with infection prevention through vaccines targeting antimicrobial-resistant pathogens, stands out as a promising frontier in the battle against AMR.
1. Phage-Derived Lytic Enzymes as Antibacterial Agents
Phage-derived lysins, also known as “enzybiotics,” present a promising alternative to conventional antibiotics. These enzymes exert their antibacterial effects by lysing host bacteria through the cleavage of peptidoglycan (PG), the primary structural component of the bacterial cell wall. Phage-derived lytic enzymes fall into two categories: virion-associated peptidoglycan hydrolases (VAPGHs) and endolysins. VAPGHs initiate degradation of PG at the onset of the phage infection [
137,
138], whereas endolysins act at the conclusion of the phage infection cycle, facilitating the release of mature phages [
139]. In most cases, dsDNA phages encode distinct VAPGHs and endolysins, whereas dsRNA phages feature a single lytic protein that serves as both VAPGH and endolysin [
140]. In contrast, ssDNA and ssRNA phages employ a single-gene lysis (Sgl) protein—an impactful degradative protein that induces cytolysis without enzymatically breaking down PGs [
141,
142].
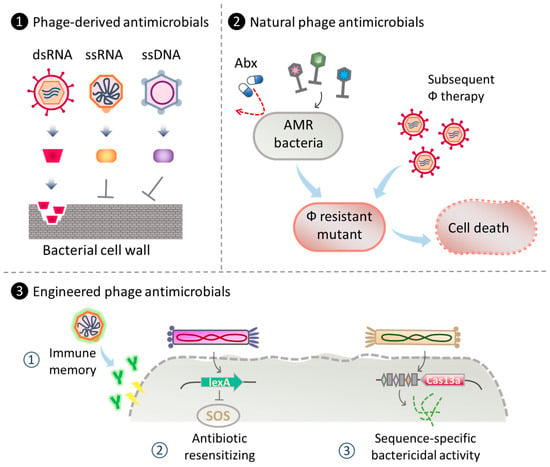
Figure 2. Strategies for phage therapy using ssDNA and RNA phages and their products. (1) Utilization of phage-derived components as antibacterial agents. For instance, the degradation of bacterial cell wall by dsRNA phage lytic enzyme [
143] or inhibition of cell wall synthesis using single-gene lysis (Slg) protein from ssDNA and ssRNA phages [
141,
142,
144]. (2) Phage cocktail employing natural ssDNA and RNA phages as antimicrobial therapeutics against AMR bacteria and phage-resistant mutants [
145,
146]. (3) Engineering ssDNA and RNA phages for antibacterial therapies. Examples include ① engineered phages displaying specific antigens as vaccine against drug-resistant bacteria [
147,
148]; ② engineered phages to enhance antibiotic efficacy in phage-antibiotic combinatory therapy [
149]; ③ engineered antibacterial capsids carrying CRISPR/Cas13a for sequence-specific-based bacterial gene detection, bacterial flora modification, and treatment for AMR bacterial infections [
65]. Abx: antibiotic.
Ply17, a lytic enzyme produced by
Pseudomonas dsRNA phage phiYY, has demonstrated the capability to reduce the number of viable Gram-negative bacteria such as
P. aeruginosa and
E. coli, as well as Gram-positive bacteria including
S. aureus and
S. epidermidis, by approximately 2 logs when treated with an outer membrane permeabilizer such as EDTA [
143]. Notably, recent genome analyses of the animal dsRNA viruses from the
Picobirnaviridae and
Partitiviridae families have unveiled potential antibacterial and antifungal lysis genes [
52]. Upon cloning and expression of the lytic genes from
Picobirnavirus and
Partitivirus in
E. coli DH5a, the observed growth inhibition was comparable to that induced by the
Enterobacteria MS2 phage lysin L [
150].
The bacteriolytic activity of ssRNA
Leviviridae phages varies among different phages. For example, Sgl
M of phiM and Sgl
PP7 of
Pseudomonas phage PP7 exhibit bacteriolytic activity by inhibiting MurJ, the transporter responsible for moving lipid-binding peptidoglycan precursors from the inside to the outside of the plasma membrane [
141,
142]. On the other hand, Sgl
Qβ from Qβ phage exhibits bacteriolytic activity by non-competitively inhibiting MurA, the first enzyme in the PG biosynthetic pathway [
151]. MS2 Sgl
MS2 (lysin L) possesses an N-terminal heat shock-responsive chaperone, DnaJ, but its specific mechanism of action remains unclear. Other known Sgls include Sgl
PRR1 (
Pseudomonas phage PRR1), Sgl
KU1 (Enterobacteria phage KU1), and Sgl
Hgal1 (Enterobacteria phage Hgal1), whose mechanisms of action have not yet been elucidated [
142].
Sgl
φ174X, a bacteriolytic enzyme produced by the ssDNA phage phiX174, inhibits MraY, an enzyme catalyzing the initial step of the lipid cycle reaction in PG biosynthesis. This inhibition results in bacteriolysis [
142,
144]. As the cellular targets and modes of action of many ssRNA and ssDNA phage Slgs are still unidentified, future genome analyses hold great capacity for the discovery of new therapeutic candidates.
2. Natural ssDNA and RNA Phages as Antibacterial Agents
Pseudomonas phages phiYY [
42] and phiZ98 [
152], which belong to the dsRNA
Cystoviridae family, utilize the lipopolysaccharide (LPS) core oligosaccharide of
P. aeruginosa as the binding receptor. Smooth-colony-type
P. aeruginosa strains exhibit resistance to phiYY due to the presence of the
galU gene, which confers O-antigen to the LPS core oligosaccharide [
145]. However, LPS-deficient
P. aeruginosa, a strain that emerged during dsDNA phage therapy and lacks the
galU gene [
153], exposes its LPS core oligosaccharide on the cell surface, making it susceptible to phiYY [
145]. Phage therapy employing phiYY has been reported to reduce bacterial load and alleviate clinical symptoms in patients diagnosed with interstitial lung disease (ILD) and chronic lung infections [
146]. Moreover, a phage cocktail, including phiYY among five phages, has demonstrated effectiveness against a broad spectrum of
P. aeruginosa clinical isolates and significantly impedes the emergence of phage-resistant mutants [
145].
dsRNA phages have also been effectively applied as plant therapeutics.
Pseudomonas phage phi6 has been employed to manage various plant diseases, including controlling
P. syringae pv. phaseolicola (Pph), the causal agent of halo disease in common bean (
Phaseolus vulgaris) [
154]. Additionally, phi6 has been utilized against
P. syringae pv. actinidiae (Psa), responsible for kiwifruit psyllid [
155], and
P. syringae pv. syringae (Pss), which causes early leaf symptoms of bean brown spot [
154]. Other
Pseudomonas phages, including phi8, phi12, phi13, phi2954, phiNN, and phiYY, have also shown efficacy against Pph [
156]. Collectively, the application of dsRNA phages holds significant promise for complementing and enhancing phage therapy for both human and plant diseases.
3. Engineering ssDNA and RNA Phages for Antimicrobial Therapy
RNA phages exhibit significant potential as antigen carriers in the development of vaccines targeting drug-resistant pathogens. Huo and colleagues pioneered the development of Qβ phage presenting Qβ-glycan 1, a synthetic tetrasaccharide from
Salmonella enteritidis. This construct successfully induced robust IgG antibody responses in both mice and rabbits [
147]. Notably, carbohydrate-based antigens, such as tetrasaccharide, do not directly interact with helper T cells, leading to limited immune memory. However, the conjugation of glycan antigens to a potent carrier such as Qβ enables engagement with both T-helper and B cells, resulting in a vigorous antibody response. Mice treated with post-immunization serum recovered from rabbits vaccinated with Qβ-glycan 1 demonstrated increased survival rates following administration of lethal doses of
S. enteritidis. In another study, Rashidijahanabad and colleagues reported the production of Qβ-OSP, a Qβ phage presenting O-specific polysaccharide (OSP) 1 of
Vibrio cholerae. Employed in mouse immunization, the Qβ-OSP conjugate elicited strong IgG antibody responses against
V. cholerae O1 Inaba, reaching sufficient IgG antibody levels after just two administrations. Remarkably, the titer of IgG antibodies remained detectable up to day 265 [
148].
Phagemid vectors derived from the filamentous ssDNA M13 phage serve as a highly efficient engineering platform for diverse therapeutic applications, including engineering phages for enhanced antibacterial activity. The delivery of antibacterial therapeutics can be achieved either through surface display on the virion or incorporated into the M13 phagemid itself. These displayed therapeutics may target vital cell components, virulent factors, or entire bacterial pathogens. Notably, several potential candidates have been identified for combating
S. aureus,
P. aeruginosa,
H. pylori, and other pathogens. In another example, the dual display of two functional peptides enables M13 to (1) undergo endocytosis by eukaryotic cells and (2) impede infection caused by the intracellular pathogen
Chlamydia trachomatis [
157]. For an in-depth exploration of anti-infective development using phage display, refer to [
158].
Antibacterial efficacy can also be achieved through cargoes loaded onto M13 phagemid. Lu and Collins cleverly designed an indirect antibacterial method using M13 phages overexpressing
lexA3, a repressor of the SOS DNA repair system [
149]. The authors demonstrated that suppressing the SOS network in
E. coli with engineered
lexA3-M13 significantly enhanced quinolone killing and increased the survival of infected mice. The engineered phage holds promise for targeting antibiotic-resistant bacteria and biofilm cells and modulating the bacterial population resistant to antibiotics post-treatment. In a parallel approach, Prokopczuk and colleagues employed a filamentous phage Pf for an indirect strategy to interfere with
P. aeruginosa pathogenesis at burn wound sites [
159]. Given that
P. aeruginosa is a major cause of burn-related infections and sepsis, often exhibiting multi-drug resistance, the authors engineered a superinfective Pf phage (eSI-Pf). Administration of eSI-Pf effectively attenuated
P. aeruginosa virulence, reduced bacterial load at the wound site, minimized bacterial dissemination from the burn site to internal organs, alleviated septicemia symptoms, and ultimately improved mouse survival.
Direct bactericidal effects have been achieved through several successful therapies, including the delivery of programmed CRISPR/Cas by recombinant M13 phages. In a proof-of-concept study on phage therapy for strain-specific depletion and genomic deletions in the gut microbiome, Lam and colleagues engineered M13 to deliver
gfp-targeting CRISPR/Cas9 to bacteria residing in the gastrointestinal tract [
160]. The engineered phage showed efficient sequence-specific targeting of GFP-expressing
E. coli in the gut, confirming the capability of CRISPR/Cas9 to induce genomic deletions at the designated target site.
This entry is adapted from the peer-reviewed paper 10.3390/ijms242317029