Agriculture is the second most important greenhouse gas (GHG: methane (CH4) and nitrous oxide (N2O) emissions)-emitting sector after the energy sector. Agriculture is also recognized as the source and sink of GHGs. Livestock production and feed, nitrogen-rich fertilizers and livestock manure application, crop residue burning, as well as water management in flood-prone cultivation areas are components of agriculture that produce and emit most GHGs. Although agriculture produces 72–89% less GHGs than other sectors, it is believed that reducing GHG emissions in agriculture would considerably lower its share of the global GHG emission records, which may lead to enormous benefits for the environment and food production systems.
- environment
- greenhouse gas
- agriculture
1. Introduction
2. Approaches to Reduce GHG Emissions in Agriculture
2.1. Improving Management of Crop Residues
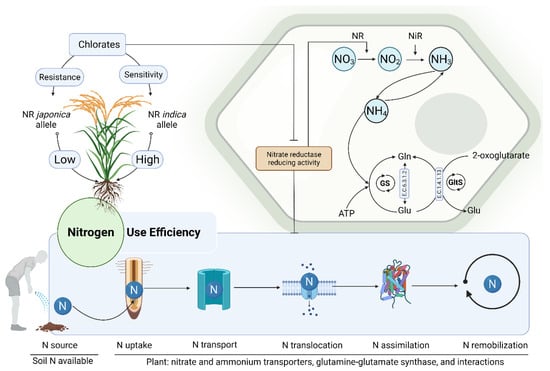
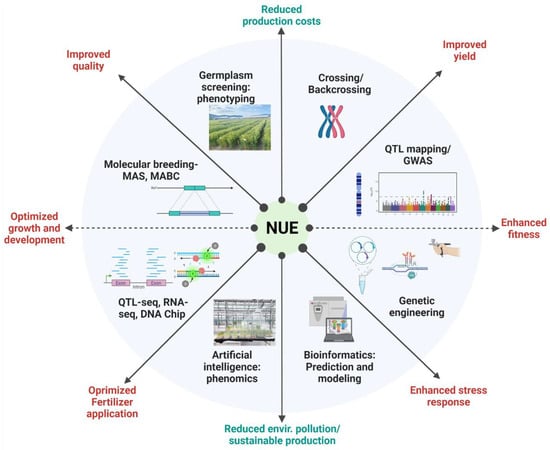
2.2. Enhancing Nitrogen Use Efficiency in Plants
2.3. Improving Abiotic Stress Tolerance in Plants
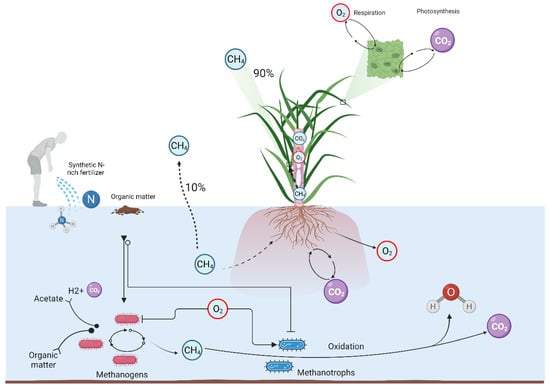
2.4. Exploring Radial Oxygen Loss and Intermittent Drainage
2.5. Biochar Reduces Mineral Fertilizers Use, Improves Soil Properties and Mitigate Ghg Emissions
2.6. Enhancing Sink Strength
2.7. Use of Nitrification Inhibitors or Low GHG-Emitting Crop Cultivars
2.8. Improving Livestock Production and Feeding Efficiency
This entry is adapted from the peer-reviewed paper 10.3390/su152215889
References
- Clapp, J.; Cohen, M.J. The Global Food Crisis: Governance Challenges and Opportunities; Wilfrid Laurier University Press: Waterloo, Canada, 2009.
- Singh, D.; Seager, R.; Cook, B.I.; Cane, M.; Ting, M.; Cook, E.; Davis, M. Climate and the Global Famine of 1876–1878. J. Clim. 2018, 31, 9445–9467.
- Gooding, P. ENSO, IOD, Drought, and Floods in Equatorial Eastern Africa, 1876–1878. In Droughts, Floods, and Global Climatic Anomalies in the Indian Ocean World; Springer: Berlin/Heidelberg, Germany, 2022; pp. 259–287.
- Holden, E.; Linnerud, K.; Banister, D. Sustainable development: Our common future revisited. Glob. Environ. Change 2014, 26, 130–139.
- Sam, S.J.E. The people, planet, prosperity, peace and partnership: Why the sustainable development goals should matter to everyone. Ecodate 2016, 30, 7–12.
- Mpabanga, D.; Sesa, L. Imperatives: The Five P’s: People, Planet, Prosperity, Peace and Partnerships and Sustainable Development Goals-The Need to Transform Public Administration and Management. Afr. J. Public Adm. Manag. 2020, 17, 44–58.
- Papoulis, D.; Kaika, D.; Bampatsou, C.; Zervas, E.J.C. Public perception of climate change in a period of economic crisis. Climate 2015, 3, 715–726.
- Adger, W.N.; Kelly, P.M. Social vulnerability to climate change and the architecture of entitlements. Mitig. Adapt. Strateg. Glob. Change 1999, 4, 253–266.
- Otto, I.M.; Reckien, D.; Reyer, C.P.; Marcus, R.; Le Masson, V.; Jones, L.; Norton, A.; Serdeczny, O. Social vulnerability to climate change: A review of concepts and evidence. Reg. Environ. Change 2017, 17, 1651–1662.
- Hasegawa, T.; Fujimori, S.; Havlík, P.; Valin, H.; Bodirsky, B.L.; Doelman, J.C.; Fellmann, T.; Kyle, P.; Koopman, J.F.; Lotze-Campen, H.; et al. Risk of increased food insecurity under stringent global climate change mitigation policy. Nat. Clim. Change 2018, 8, 699–703.
- Ericksen, P.J.; Thornton, P.K.; Notenbaert, A.M.O.; Cramer, L.; Jones, P.G.; Herrero, M.T. Mapping hotspots of climate change and food insecurity in the global tropics. In CCAFS Report; CCAFS: Copenhagen, Denmark, 2011.
- Crutzen, P.J.J.N. Methane’s sinks and sources. Nature 1991, 350, 380–381.
- Reay, D. Greenhouse Gas Sinks; CABI: Wallingford, UK, 2007.
- Reay, D.; Smith, P.; Van Amstel, A. Methane sources and the global methane budget. In Methane and Climate Change; Earthscan: London, UK; Washington, DC, USA, 2010; pp. 1–13.
- Casper, J.K. Agriculture: The Food We Grow and Animals We Raise; Infobase Publishing: New York, NY, USA, 2007.
- Bhuvaneshwari, S.; Hettiarachchi, H.; Meegoda, J.N. Crop residue burning in India: Policy challenges and potential solutions. Int. J. Environ. Res. Public Health 2019, 16, 832.
- Ma, J.; Li, X.L.; Xu, H.; Han, Y.; Cai, Z.C.; Yagi, K. Effects of nitrogen fertiliser and wheat straw application on CH4 and N2O emissions from a paddy rice field. Soil Res. 2007, 45, 359–367.
- Horwath, W. Greenhouse gas emissions from rice cropping systems. In Understanding Greenhouse Gas Emissions from Agricultural Management; ACS Publications: Washington, DC, USA, 2011; pp. 67–89.
- Hoornweg, D.; Bhada-Tata, P. What a Waste: A Global Review of Solid Waste Management; World Bank: Washington, DC, USA, 2012.
- Rakotoson, T.; Dusserre, J.; Letourmy, P.; Frouin, J.; Ratsimiala, I.R.; Rakotoarisoa, N.V.; Vom Brocke, K.; Ramanantsoanirina, A.; Ahmadi, N.; Raboin, L.M. Genome-wide association study of nitrogen use efficiency and agronomic traits in upland rice. Rice Sci. 2021, 28, 379–390.
- Bandyopadhyay, T.; Swarbreck, S.M.; Jaiswal, V.; Maurya, J.; Gupta, R.; Bentley, A.R.; Griffiths, H.; Prasad, M. GWAS identifies genetic loci underlying nitrogen responsiveness in the climate resilient C4 model Setaria italica (L.). J. Adv. Res. 2022, 42, 249–261.
- Akhatar, J.; Goyal, A.; Kaur, N.; Atri, C.; Mittal, M.; Singh, M.P.; Kaur, R.; Rialch, I.; Banga, S.S. Genome wide association analyses to understand genetic basis of flowering and plant height under three levels of nitrogen application in Brassica juncea (L.) Czern & Coss. Sci. Rep. 2021, 11, 4278.
- Nazish, T.; Arshad, M.; Jan, S.U.; Javaid, A.; Khan, M.H.; Naeem, M.A.; Baber, M.; Ali, M. Transporters and transcription factors gene families involved in improving nitrogen use efficiency (NUE) and assimilation in rice (Oryza sativa L.). Transgenic Res. 2021, 31, 23–42.
- Wang, Q.; Su, Q.; Nian, J.; Zhang, J.; Guo, M.; Dong, G.; Hu, J.; Wang, R.; Wei, C.; Li, G.; et al. The Ghd7 transcription factor represses ARE1 expression to enhance nitrogen utilization and grain yield in rice. Mol. Plant 2021, 14, 1012–1023.
- Han, M.L.; Lv, Q.Y.; Zhang, J.; Wang, T.; Zhang, C.X.; Tan, R.J.; Wang, Y.L.; Zhong, L.Y.; Gao, Y.Q.; Chao, Z.F.; et al. Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of Nitrate Reductase 1.2 in rice. Mol. Plant 2022, 15, 167–178.
- Zhang, Z.; Chu, C. Nitrogen-use divergence between indica and japonica rice: Variation at nitrate assimilation. Mol. Plant 2020, 13, 6–7.
- Teng, W.; He, X.; Tong, Y. Genetic Control of Efficient Nitrogen Use for High Yield and Grain Protein Concentration in Wheat: A Review. Plants 2022, 11, 492.
- Kabange, N.R.; Park, S.Y.; Shin, D.; Lee, S.M.; Jo, S.M.; Kwon, Y.; Cha, J.K.; Song, Y.C.; Ko, J.M.; Lee, J.H. Identification of a novel QTL for chlorate resistance in rice (Oryza sativa L.). Agriculture 2020, 10, 360.
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47.
- Li, Q.; Lu, X.; Wang, C.; Shen, L.; Dai, L.; He, J.; Yang, L.; Li, P.; Hong, Y.; Zhang, Q.; et al. Genome-wide association study and transcriptome analysis reveal new QTL and candidate genes for nitrogen-deficiency tolerance in rice. Crop J. 2022, 10, 942–951.
- Fan, X.; Naz, M.; Fan, X.; Xuan, W.; Miller, A.J.; Xu, G. Plant nitrate transporters: From gene function to application. J. Exp. Bot. 2017, 68, 2463–2475.
- Lea, P.J.; Miflin, B.J. Glutamate synthase and the synthesis of glutamate in plants. Plant Physiol. Biochem. 2003, 41, 555–564.
- Prasad, L.R.V.; Mailapalli, D.R. Evaluation of nitrogen fertilization patterns using DSSAT for enhancing grain yield and nitrogen use efficiency in rice. Commun. Soil Sci. Plant Anal. 2018, 49, 1401–1417.
- Nakidakida, T.; Hayashi, H. Nitrogen recovery and nitrogen use efficiency of potatoes in an integrated compost fertilization system in an Andosol soil. In Proceedings of the III International Symposium on Organic Matter Management and Compost Use in Horticulture 1146, Murcia, Spain, 6 April 2015; pp. 41–48.
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: Lessons from Arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488.
- Duan, Y.H.; Zhang, Y.L.; Ye, L.T.; Fan, X.R.; Xu, G.H.; Shen, Q.R. Responses of rice cultivars with different nitrogen use efficiency to partial nitrate nutrition. Ann. Bot. 2007, 99, 1153–1160.
- Djaman, K.; Mel, V.C.; Ametonou, F.Y.; El-Namaky, R.; Diallo, M.D.; Koudahe, K. Effect of nitrogen fertilizer dose and application timing on yield and nitrogen use efficiency of irrigated hybrid rice under semi-arid conditions. J. Agric. Sci. Food Res. 2018, 9, 223.
- Gallais, A.; Hirel, B. An approach to the genetics of nitrogen use efficiency in maize. J. Exp. Bot. 2004, 55, 295–306.
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J. Genet. Genom. 2021, 49, 394–404.
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving nitrogen use efficiency in crops for sustainable agriculture. Sustainability 2011, 3, 1452–1485.
- Yamaya, T.; Obara, M.; Nakajima, H.; Sasaki, S.; Hayakawa, T.; Sato, T. Genetic manipulation and quantitative-trait loci mapping for nitrogen recycling in rice. J. Exp. Bot. 2002, 53, 917–925.
- Zhang, S.; Zhu, L.; Shen, C.; Ji, Z.; Zhang, H.; Zhang, T.; Li, Y.; Yu, J.; Yang, N.; He, Y.; et al. Natural allelic variation in a modulator of auxin homeostasis improves grain yield and nitrogen use efficiency in rice. Plant Cell 2021, 33, 566–580.
- Kiba, T.; Kudo, T.; Kojima, M.; Sakakibara, H. Hormonal control of nitrogen acquisition: Roles of auxin, abscisic acid, and cytokinin. J. Exp. Bot. 2011, 62, 1399–1409.
- Séguéla, M.; Briat, J.F.; Vert, G.; Curie, C. Cytokinins negatively regulate the root iron uptake machinery in Arabidopsis through a growth-dependent pathway. Plant J. 2008, 55, 289–300.
- Maruyama-Nakashita, A.; Nakamura, Y.; Yamaya, T.; Takahashi, H. A novel regulatory pathway of sulfate uptake in Arabidopsis roots: Implication of CRE1/WOL/AHK4-mediated cytokinin-dependent regulation. Plant J. 2004, 38, 779–789.
- Wagner, B.M.; Beck, E. Cytokinins in the perennial herb Urtica dioica L. as influenced by its nitrogen status. Planta 1993, 190, 511–518.
- Franco-Zorrilla, J.M.; Martin, A.C.; Solano, R.; Rubio, V.; Leyva, A.; Paz-Ares, J. Mutations at CRE1 impair cytokinin-induced repression of phosphate starvation responses in Arabidopsis. Plant J. 2002, 32, 353–360.
- Nacry, P.; Canivenc, G.; Muller, B.; Azmi, A.; Van Onckelen, H.; Rossignol, M.; Doumas, P. A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. Plant Physiol. 2005, 138, 2061–2074.
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482.
- Chen, K.-E.; Chen, H.-Y.; Tseng, C.-S.; Tsay, Y.-F. Improving nitrogen use efficiency by manipulating nitrate remobilization in plants. Nat. Plants 2020, 6, 1126–1135.
- Ueda, Y.; Yanagisawa, S. Transcription factor-based genetic engineering to increase nitrogen use efficiency. In Engineering Nitrogen Utilization in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–55.
- Wu, J.; Zhang, Z.S.; Xia, J.Q.; Alfatih, A.; Song, Y.; Huang, Y.J.; Wan, G.Y.; Sun, L.Q.; Tang, H.; Liu, Y. Rice NIN-LIKE PROTEIN 4 plays a pivotal role in nitrogen use efficiency. Plant Biotechnol. J. 2021, 19, 448–461.
- Li, Q.; Ding, G.; Yang, N.; White, P.J.; Ye, X.; Cai, H.; Lu, J.; Shi, L.; Xu, F. Comparative genome and transcriptome analysis unravels key factors of nitrogen use efficiency in Brassica napus L. Plant Cell Environ. 2020, 43, 712–731.
- Heuermann, D.; Hahn, H.; Von Wirén, N. Seed yield and nitrogen efficiency in oilseed rape after ammonium nitrate or urea fertilization. Front. Plant Sci. 2021, 11, 2197.
- Ruffel, S.; Krouk, G.; Ristova, D.; Shasha, D.; Birnbaum, K.D.; Coruzzi, G.M. Nitrogen economics of root foraging: Transitive closure of the nitrate–cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc. Natl. Acad. Sci. USA 2011, 108, 18524–18529.
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. Plant Sci. 2018, 274, 320–331.
- Azarakhsh, M.; Lebedeva, M.A.; Lutova, L.A. Identification and expression analysis of Medicago truncatula isopentenyl transferase genes (IPTs) involved in local and systemic control of nodulation. Front. Plant Sci. 2018, 9, 304.
- Lin, J.; Roswanjaya, Y.P.; Kohlen, W.; Stougaard, J.; Reid, D. Nitrate restricts nodule organogenesis through inhibition of cytokinin biosynthesis in Lotus japonicus. Nat. Commun. 2021, 12, 6544.
- Sasaki, T.; Suzaki, T.; Soyano, T.; Kojima, M.; Sakakibara, H.; Kawaguchi, M. Shoot-derived cytokinins systemically regulate root nodulation. Nat. Commun. 2014, 5, 4983.
- Singh, B.N.; Dwivedi, P.; Sarma, B.K.; Singh, G.S.; Singh, H.B. A novel function of N-signaling in plants with special reference to Trichoderma interaction influencing plant growth, nitrogen use efficiency, and cross talk with plant hormones. 3 Biotech 2019, 9, 109.
- Mundim, F.M.; Pringle, E.G. Whole-plant metabolic allocation under water stress. Front. Plant Sci. 2018, 9, 852.
- Rolly, N.K.; Yun, B.-W. Regulation of Nitrate (NO3) Transporters and Glutamate Synthase-Encoding Genes under Drought Stress in Arabidopsis: The Regulatory Role of AtbZIP62 Transcription Factor. Plants 2021, 10, 2149.
- Rolly, N.K.; Mun, B.-G.; Yun, B.-W. Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. Genes 2021, 12, 298.
- Sun, L.; Di, D.-W.; Li, G.; Li, Y.; Kronzucker, H.J.; Shi, W. Transcriptome analysis of rice (Oryza sativa L.) in response to ammonium resupply reveals the involvement of phytohormone signaling and the transcription factor OsJAZ9 in reprogramming of nitrogen uptake and metabolism. J. Plant Physiol. 2020, 246, 153137.
- Zhong, L.; Chen, D.; Min, D.; Li, W.; Xu, Z.; Zhou, Y.; Li, L.; Chen, M.; Ma, Y. AtTGA4, a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 457, 433–439.
- Gupta, K.J. Plant Respiration and Internal Oxygen: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2017.
- Armstrong, W.; Armstrong, J. Plant internal oxygen transport (diffusion and convection) and measuring and modelling oxygen gradients. In Low-Oxygen Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2014; pp. 267–297.
- Kosmacz, M.; Weits, D.A. Oxygen perception in plants. In Low-Oxygen Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–17.
- Guan, B.; Lin, Z.; Liu, D.; Li, C.; Zhou, Z.; Mei, F.; Li, J.; Deng, X. Effect of waterlogging-induced autophagy on programmed cell death in Arabidopsis roots. Front. Plant Sci. 2019, 10, 468.
- Armstrong, W. Advances in Botanical Research, Volume 7. Aeration in Higher Plants; Woolhouse, H.W., Ed.; Academic Press: Cambridge, MA, USA, 1979.
- Sifton, H.B. Air-space tissue in plants. Bot. Rev. 1945, 11, 108–143.
- Roland, J.C. Cell wall differentiation and stages involved with intercellular gas space opening. J. Cell Sci. 1978, 32, 325–336.
- Jeffree, C.; Dale, J.; Fry, S.C. The genesis of intercellular spaces in developing leaves of Phaseolus vulgaris L. Protoplasma 1986, 132, 90–98.
- Raven, J.A. Into the voids: The distribution, function, development and maintenance of gas spaces in plants. Ann. Bot. 1996, 78, 137–142.
- Wegner, L.H. Oxygen transport in waterlogged plants. In Waterlogging Signalling and Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3–22.
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36.
- Lai, W.-L.; Zhang, Y.; Chen, Z.-H. Radial oxygen loss, photosynthesis, and nutrient removal of 35 wetland plants. Ecol. Eng. 2012, 39, 24–30.
- Mohammed, U.; Caine, R.; Atkinson, J.; Harrison, E.; Wells, D.; Chater, C.; Gray, J.; Swarup, R.; Murchie, E.H. Rice plants overexpressing OsEPF1 show reduced stomatal density and increased root cortical aerenchyma formation. Sci. Rep. 2019, 9, 5584.
- Scheehle, E.A.; Kruger, D. Global anthropogenic methane and nitrous oxide emissions. Energy J. 2006.
- Denmead, O.; Freney, J.; Simpson, J.R. Nitrous oxide emission during denitrification in a flooded field. Soil Sci. Soc. Am. J. 1979, 43, 716–718.
- Bodelier, P.L.; Pérez, G.; Veraart, A.J.; Krause, S. Methanotroph ecology, environmental distribution and functioning. In Methanotrophs; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–38.
- Reim, A.; Lüke, C.; Krause, S.; Pratscher, J.; Frenzel, P. One millimetre makes the difference: High-resolution analysis of methane-oxidizing bacteria and their specific activity at the oxic–anoxic interface in a flooded paddy soil. ISME J. 2012, 6, 2128–2139.
- Trotsenko, Y.A.; Murrell, J.C. Metabolic aspects of aerobic obligate methanotrophy⋆. Adv. Appl. Microbiol. 2008, 63, 183–229.
- Dalal, R.C.; Allen, D.E.; Livesley, S.J.; Richards, G. Magnitude and biophysical regulators of methane emission and consumption in the Australian agricultural, forest, and submerged landscapes: A review. Plant Soil 2008, 309, 43–76.
- Saggar, S.; Tate, K.; Giltrap, D.; Singh, J. Soil-atmosphere exchange of nitrous oxide and methane in New Zealand terrestrial ecosystems and their mitigation options: A review. Plant Soil 2008, 309, 25–42.
- Schütz, H.; Seiler, W.; Rennenberg, H. Soil and land use related sources and sinks of methane (CH4) in the context of the global methane budget. In Soils and the Greenhouse Effect; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 1990; pp. 269–285.
- Walkiewicz, A.; Brzezińska, M.; Bieganowski, A. Methanotrophs are favored under hypoxia in ammonium-fertilized soils. Biol. Fertil. Soils 2018, 54, 861–870.
- Colmer, T.D.; Kotula, L.; Malik, A.I.; Takahashi, H.; Konnerup, D.; Nakazono, M.; Pedersen, O. Rice acclimation to soil flooding: Low concentrations of organic acids can trigger a barrier to radial oxygen loss in roots. Plant Cell Environ. 2019, 42, 2183–2197.
- Ejiri, M.; Shiono, K. Prevention of radial oxygen loss is associated with exodermal suberin along adventitious roots of annual wild species of Echinochloa. Front. Plant Sci. 2019, 10, 254.
- Abiko, T.; Kotula, L.; Shiono, K.; Malik, A.I.; Colmer, T.D.; Nakazono, M. Enhanced formation of aerenchyma and induction of a barrier to radial oxygen loss in adventitious roots of Zea nicaraguensis contribute to its waterlogging tolerance as compared with maize (Zea mays ssp. mays). Plant Cell Environ. 2012, 35, 1618–1630.
- Ejiri, M.; Fukao, T.; Miyashita, T.; Shiono, K. A barrier to radial oxygen loss helps the root system cope with waterlogging-induced hypoxia. Breed. Sci. 2021, 71, 40–50.
- Peralta Ogorek, L.L.; Pellegrini, E.; Pedersen, O. Novel functions of the root barrier to radial oxygen loss–radial diffusion resistance to H2 and water vapour. New Phytol. 2021, 231, 1365–1376.
- Duyen, D.V.; Kwon, Y.; Kabange, N.R.; Lee, J.Y.; Lee, S.M.; Kang, J.W.; Park, H.; Cha, J.K.; Cho, J.H.; Shin, D.; et al. Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide. Plants 2022, 11, 788.
- Foyer, C.H.; Noctor, G.; Hodges, M. Respiration and nitrogen assimilation: Targeting mitochondria-associated metabolism as a means to enhance nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1467–1482.
- Maier, R.J. Nitrogen fixation and respiration: Two processes linked by the energetic demands of Nitrogenase. In Respiration in Archaea and Bacteria; Springer: Berlin/Heidelberg, Germany, 2004; pp. 101–120.
- Weger, H.G.; Turpin, D.H. Mitochondrial respiration Can support NO3− and NO2− reduction during photosynthesis: Interactions between photosynthesis, respiration, and N assimilation in the N-limited green alga Selenastrum minutum. Plant Physiol. 1989, 89, 409–415.
- De Laulanié, H. Intensive rice farming in Madagascar. Tropicultura 2011, 29, 183–187.
- Sinclair, T.R. Agronomic UFOs waste valuable scientific resources. Rice Today 2004, 3, 43.
- Pereira-Mora, L.; Terra, J.A.; Fernández-Scavino, A. Methanogenic community linked to organic acids fermentation from root exudates are affected by rice intensification in rotational soil systems. Appl. Soil Ecol. 2022, 176, 104498.
- Xu, Z.; Chan, K. Biochar: Nutrient properties and their enhancement. Biochar Environ. Manag. 2009, 1, 67–84.
- Lehmann, J.; Cowie, A.; Masiello, C.A.; Kammann, C.; Woolf, D.; Amonette, J.E.; Cayuela, M.L.; Camps-Arbestain, M.; Whitman, T. Biochar in climate change mitigation. Nat. Geosci. 2021, 14, 883–892.
- Roe, S.; Streck, C.; Beach, R.; Busch, J.; Chapman, M.; Daioglou, V.; Deppermann, A.; Doelman, J.; Emmet-Booth, J.; Engelmann, J.; et al. Land-based measures to mitigate climate change: Potential and feasibility by country. Glob. Change Biol. 2021, 27, 6025–6058.
- Yang, W.; Feng, G.; Miles, D.; Gao, L.; Jia, Y.; Li, C.; Qu, Z. Impact of biochar on greenhouse gas emissions and soil carbon sequestration in corn grown under drip irrigation with mulching. Sci. Total Environ. 2020, 729, 138752.
- Lyu, H.; Zhang, H.; Chu, M.; Zhang, C.; Tang, J.; Chang, S.X.; Mašek, O.; Ok, Y.S. Biochar affects greenhouse gas emissions in various environments: A critical review. Land Degrad. Dev. 2022, 33, 3327–3342.
- Lin, X.; Xie, Z.; Zheng, J.; Liu, Q.; Bei, Q.; Zhu, J.G. Effects of biochar application on greenhouse gas emissions, carbon sequestration and crop growth in coastal saline soil. Eur. J. Soil Sci. 2015, 66, 329–338.
- Lehmann, J.; Joseph, S. Biochar for environmental management: An introduction. In Biochar for Environmental Management; Routledge: London, UK, 2015; p. 1.
- Lehmann, J.; Czimczik, C.; Laird, D.; Sohi, S. Stability of biochar in soil. In Biochar for Environmental Management; Routledge: London, UK, 2009; pp. 183–206.
- Major, J.; Steiner, C.; Downie, A.; Lehmann, J. Biochar effects on nutrient leaching. In Biochar for Environmental Management; Routledge: London, UK, 2009; pp. 303–320.
- Turunen, M.; Hyväluoma, J.; Heikkinen, J.; Keskinen, R.; Kaseva, J.; Hannula, M.; Rasa, K. Quantifying the pore structure of different biochars and their impacts on the water retention properties of Sphagnum moss growing media. Biosyst. Eng. 2020, 191, 96–106.
- Gaunt, J.; Cowie, A. Biochar, greenhouse gas accounting and emissions trading. In Biochar for Environmental Management: Science and Technology; Routledge: London, UK, 2009; pp. 317–340.
- Smernik, R.J. Biochar and sorption of organic compounds. In Biochar for Environmental Management; Routledge: London, UK, 2009; pp. 321–332.
- Thies, J.E.; Rillig, M.C. Characteristics of biochar: Biological properties. In Biochar for Environmental Management; Routledge: London, UK, 2012; pp. 117–138.
- DeLuca, T.H.; Gundale, M.J.; MacKenzie, M.D.; Jones, D.L. Biochar effects on soil nutrient transformations. In Biochar for Environmental Management: Science, Technology and Implementation; Routledge: London, UK, 2015; Volume 2, pp. 421–454.
- Rondon, M.A.; Molina, D.; Hurtado, M.; Ramirez, J.; Lehmann, J.; Major, J.; Amezquita, E. Enhancing the productivity of crops and grasses while reducing greenhouse gas emissions through bio-char amendments to unfertile tropical soils. In Proceedings of the 18th World Congress of Soil Science, Philadelphia, PA, USA, 9–15 July 2006; pp. 9–15.
- Yanai, Y.; Toyota, K.; Okazaki, M. Effects of charcoal addition on N2O emissions from soil resulting from rewetting air-dried soil in short-term laboratory experiments. Soil Sci. Plant Nutr. 2007, 53, 181–188.
- Joseph, S.; Cowie, A.L.; Van Zwieten, L.; Bolan, N.; Budai, A.; Buss, W.; Cayuela, M.L.; Graber, E.R.; Ippolito, J.A.; Kuzyakov, Y.; et al. How biochar works, and when it doesn’t: A review of mechanisms controlling soil and plant responses to biochar. Gcb Bioenergy 2021, 13, 1731–1764.
- Joseph, S.; Peacocke, C.; Lehmann, J.; Munroe, P. Developing a biochar classification and test methods. In Biochar for Environmental Management; Routledge: London, UK, 2009; Volume 1, pp. 139–158.
- Borchard, N.; Schirrmann, M.; Cayuela, M.L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizábal, T.; Sigua, G.; Spokas, K.; Ippolito, J.A.; et al. Biochar, soil and land-use interactions that reduce nitrate leaching and N2O emissions: A meta-analysis. Sci. Total Environ. 2019, 651, 2354–2364.
- Riley, W.; Ortiz-Monasterio, I.; Matson, P.A. Nitrogen leaching and soil nitrate, nitrite, and ammonium levels under irrigated wheat in Northern Mexico. Nutr. Cycl. Agroecosystems 2001, 61, 223–236.
- Song, X.; Pan, G.; Zhang, C.; Zhang, L.; Wang, H. Effects of biochar application on fluxes of three biogenic greenhouse gases: A meta-analysis. Ecosyst. Health Sustain. 2016, 2, e01202.
- He, Y.; Zhou, X.; Jiang, L.; Li, M.; Du, Z.; Zhou, G.; Shao, J.; Wang, X.; Xu, Z.; Hosseini Bai, S.; et al. Effects of biochar application on soil greenhouse gas fluxes: A meta-analysis. Gcb Bioenergy 2017, 9, 743–755.
- Kalu, S.; Kulmala, L.; Zrim, J.; Peltokangas, K.; Tammeorg, P.; Rasa, K.; Kitzler, B.; Pihlatie, M.; Karhu, K. Potential of biochar to reduce greenhouse gas emissions and increase nitrogen use efficiency in boreal arable soils in the long-term. Front. Environ. Sci. 2022, 10, 914766.
- Sass, R.L.; Cicerone, R.J. Photosynthate allocations in rice plants: Food production or atmospheric methane? Proc. Natl. Acad. Sci. USA 2002, 99, 11993–11995.
- van Der Gon, H.D.; Kropff, M.; Van Breemen, N.; Wassmann, R.; Lantin, R.; Aduna, E.; Corton, T.; Van Laar, H.H. Optimizing grain yields reduces CH4 emissions from rice paddy fields. Proc. Natl. Acad. Sci. USA 2002, 99, 12021–12024.
- Su, J.; Hu, C.; Yan, X.; Jin, Y.; Chen, Z.; Guan, Q.; Wang, Y.; Zhong, D.; Jansson, C.; Wang, F.; et al. Expression of barley SUSIBA2 transcription factor yields high-starch low-methane rice. Nature 2015, 523, 602–606.
- Stein, L.Y.; Klotz, M.G. Nitrifying and denitrifying pathways of methanotrophic bacteria. Biochem. Soc. Trans. 2011, 39, 1826–1831.
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli. Front. Plant Sci. 2019, 10, 157.
- Rougier, M. Secretory activity of the root cap. In Plant Carbohydrates II: Extracellular Carbohydrates; Springer: Berlin/Heidelberg, Germany, 1981; pp. 542–574.
- Abbott, L.K.; Murphy, D.V. Soil Biological Fertility: A Key to Sustainable Land Use in Agriculture; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003.
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root exudation and rhizosphere biology. Plant Physiol. 2003, 132, 44–51.
- Nardi, S.; Concheri, G.; Pizzeghello, D.; Sturaro, A.; Rella, R.; Parvoli, G. Soil organic matter mobilization by root exudates. Chemosphere 2000, 41, 653–658.
- Vives-Peris, V.; De Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17.
- De-la-Pena, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brandao, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M. Root secretion of defense-related proteins is development-dependent and correlated with flowering time. J. Biol. Chem. 2010, 285, 30654–30665.
- Chai, Y.N.; Schachtman, D.P. Root exudates impact plant performance under abiotic stress. Trends Plant Sci. 2022, 27, 80–91.
- Salem, M.A.; Wang, J.Y.; Al-Babili, S. Metabolomics of plant root exudates: From sample preparation to data analysis. Front. Plant Sci. 2022, 13, 5035.
- Narula, N.; Kothe, E.; Behl, R.K. Role of root exudates in plant-microbe interactions. J. Appl. Bot. Food Qual. 2012, 82, 122–130.
- Kumar, R.; Bhatia, R.; Kukreja, K.; Behl, R.K.; Dudeja, S.S.; Narula, N. Establishment of Azotobacter on plant roots: Chemotactic response, development and analysis of root exudates of cotton (Gossypium hirsutum L.) and wheat (Triticum aestivum L.). J. Basic Microbiol. 2007, 47, 436–439.
- Lu, Y.; Wassmann, R.; Neue, H.; Huang, C.; Bueno, C.S. Methanogenic responses to exogenous substrates in anaerobic rice soils. Soil Biol. Biochem. 2000, 32, 1683–1690.
- Moscôso, J.S.C.; Silva, L.S.d.; Pujol, S.B.; Giacomini, S.J.; Severo, F.F.; Marzari, L.B.; Molin, G.D. Methane emission induced by short-chain organic acids in lowland soil. Rev. Bras. Ciência Solo 2019, 43.
- Aulakh, M.S.; Wassmann, R.; Bueno, C.; Rennenberg, H. Impact of root exudates of different cultivars and plant development stages of rice (Oryza sativa L.) on methane production in a paddy soil. Plant Soil 2001, 230, 77–86.
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. Front. Plant Sci. 2018, 9, 1889.
- Andersen, C.P. Source–sink balance and carbon allocation below ground in plants exposed to ozone. New Phytol. 2003, 157, 213–228.
- Wingler, A.; Einig, W.; Schaeffer, C.; Wallenda, T.; Hampp, R.; Wallander, H.; Nylund, J.E. Influence of different nutrient regimes on the regulation of carbon metabolism in Norway spruce seedlings. New Phytol. 1994, 128, 323–330.
- Koch, K.E. Carbohydrate-modulated gene expression in plants. Annu. Rev. Plant Biol. 1996, 47, 509–540.
- Mathur, J.; Hülskamp, M. Microtubules and microfilaments in cell morphogenesis in higher plants. Curr. Biol. 2002, 12, R669–R676.
- Yao, Z.; Zhou, Z.; Zheng, X.; Xie, B.; Mei, B.; Wang, R.; Butterbach-Bahl, K.; Zhu, J. Effects of organic matter incorporation on nitrous oxide emissions from rice-wheat rotation ecosystems in China. Plant Soil 2010, 327, 315–330.
- Zou, J.; Huang, Y.; Jiang, J.; Zheng, X.; Sass, R.L. A 3-year field measurement of methane and nitrous oxide emissions from rice paddies in China: Effects of water regime, crop residue, and fertilizer application. Glob. Biogeochem. Cycles 2005, 19.
- Eagle, A.J.; Bird, J.A.; Horwath, W.R.; Linquist, B.A.; Brouder, S.M.; Hill, J.E.; van Kessel, C. Rice yield and nitrogen utilization efficiency under alternative straw management practices. Agron. J. 2000, 92, 1096–1103.
- Li, X.; Zhang, G.; Xu, H.; Cai, Z.; Yagi, K. Effect of timing of joint application of hydroquinone and dicyandiamide on nitrous oxide emission from irrigated lowland rice paddy field. Chemosphere 2009, 75, 1417–1422.
- Akiyama, H.; Yan, X.; Yagi, K. Evaluation of effectiveness of enhanced-efficiency fertilizers as mitigation options for N2O and NO emissions from agricultural soils: Meta-analysis. Glob. Change Biol. 2010, 16, 1837–1846.
- Majumdar, D. Methane and nitrous oxide emission from irrigated rice fields: Proposed mitigation strategies. Curr. Sci. 2003, 84, 1317–1326.
- Mahesh, M.; Mohini, M. Crop residues for sustainable livestock production. J. Adv. Dairy Res. 2014, 2, e108.
- Ritchie, H.; Rosado, P.; Roser, M. Meat and Dairy Production. Our World Data 2017. Available online: https://ourworldindata.org/meat-production (accessed on 18 October 2023).
- Williams, P. Nutritional composition of red meat. Nutr. Diet. 2007, 64, S113–S119.
- Porter, J.W.G. Milk as a source of lactose, vitamins and minerals. Proc. Nutr. Soc. 1978, 37, 225–230.
- Shaheen, N.; Ahmed, M.K.; Islam, M.S.; Habibullah-Al-Mamun, M.; Tukun, A.B.; Islam, S.; MA Rahim, A.T. Health risk assessment of trace elements via dietary intake of ‘non-piscine protein source’foodstuffs (meat, milk and egg) in Bangladesh. Environ. Sci. Pollut. Res. 2016, 23, 7794–7806.
- McAfee, A.J.; McSorley, E.M.; Cuskelly, G.J.; Moss, B.W.; Wallace, J.M.; Bonham, M.P.; Fearon, A.M. Red meat consumption: An overview of the risks and benefits. Meat Sci. 2010, 84, 1–13.
- Willett, W.C.; Ludwig, D.S. Milk and health. N. Engl. J. Med. 2020, 382, 644–654.
- Tricarico, J.; de Haas, Y.; Hristov, A.; Kebreab, E.; Kurt, T.; Mitloehner, F.; Pitta, D. Symposium review: Development of a funding program to support research on enteric methane mitigation from ruminants. J. Dairy Sci. 2022, 105, 8535–8542.
- United Nations Environment Programme. Emissions Gap Report 2022: The Closing Window—Climate Crisis Calls for Rapid Transformation of Societies; UN: New York, NY, USA, 2022.
- Frank, S.; Beach, R.; Havlík, P.; Valin, H.; Herrero, M.; Mosnier, A.; Hasegawa, T.; Creason, J.; Ragnauth, S.; Obersteiner, M. Structural change as a key component for agricultural non-CO2 mitigation efforts. Nat. Commun. 2018, 9, 1060.
- Arndt, C.; Hristov, A.N.; Price, W.J.; McClelland, S.C.; Pelaez, A.M.; Cueva, S.F.; Oh, J.; Dijkstra, J.; Bannink, A.; Bayat, A.R.; et al. Full adoption of the most effective strategies to mitigate methane emissions by ruminants can help meet the 1.5 C target by 2030 but not 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2111294119.
- Zhao, Y.; Zhao, G. Decreasing ruminal methane production through enhancing the sulfate reduction pathway. Anim. Nutr. 2022, 9, 320–326.
- Haque, M.N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 15.
- Newbold, C.; El Hassan, S.; Wang, J.; Ortega, M.; Wallace, R.J. Influence of foliage from African multipurpose trees on activity of rumen protozoa and bacteria. Br. J. Nutr. 1997, 78, 237–249.
- Bodas, R.; Prieto, N.; García-González, R.; Andrés, S.; Giráldez, F.J.; López, S. Manipulation of rumen fermentation and methane production with plant secondary metabolites. Anim. Feed. Sci. Technol. 2012, 176, 78–93.
- Patra, A.K.; Saxena, J. Dietary phytochemicals as rumen modifiers: A review of the effects on microbial populations. Antonie Van Leeuwenhoek 2009, 96, 363–375.
- Patra, A.; Saxena, J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr. Res. Rev. 2009, 22, 204–219.
- Milich, L. The role of methane in global warming: Where might mitigation strategies be focused? Glob. Environ. Change 1999, 9, 179–201.
- Benchaar, C.; Pomar, C.; Chiquette, J. Evaluation of dietary strategies to reduce methane production in ruminants: A modelling approach. Can. J. Anim. Sci. 2001, 81, 563–574.
- Boadi, D.; Wittenberg, K.M. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. Can. J. Anim. Sci. 2002, 82, 201–206.
- Beauchemin, K.; Kreuzer, M.; O’mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27.
- Tamminga, S.; Bannink, A.; Dijkstra, J.; Zom, R. Feeding Strategies to Reduce Methane Loss in Cattle; Animal Sciences Group: Wageningen, The Netherlands, 2007; pp. 1570–8616.
- O’mara, F.; Fitzgerald, J.; Murphy, J.; Rath, M. The effect on milk production of replacing grass silage with maize silage in the diet of dairy cows. Livest. Prod. Sci. 1998, 55, 79–87.
- Hassanat, F.; Gervais, R.; Julien, C.; Massé, D.; Lettat, A.; Chouinard, P.; Petit, H.; Benchaar, C. Replacing alfalfa silage with corn silage in dairy cow diets: Effects on enteric methane production, ruminal fermentation, digestion, N balance, and milk production. J. Dairy Sci. 2013, 96, 4553–4567.
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365.
- Ferris, C.; Gordon, F.; Patterson, D.; Porter, M.; Yan, T. The effect of genetic merit and concentrate proportion in the diet on nutrient utilization by lactating dairy cows. J. Agric. Sci. 1999, 132, 483–490.
- Lovett, D.; Lovell, S.; Stack, L.; Callan, J.; Finlay, M.; Conolly, J.; O’Mara, F.P. Effect of forage/concentrate ratio and dietary coconut oil level on methane output and performance of finishing beef heifers. Livest. Prod. Sci. 2003, 84, 135–146.
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492.
- Murphy, M.; Baldwin, R.; Koong, L.J. Estimation of stoichiometric parameters for rumen fermentation of roughage and concentrate diets. J. Anim. Sci. 1982, 55, 411–421.
- Hindrichsen, I.; Wettstein, H.; Machmüller, A.; Jörg, B.; Kreuzer, M. Effect of the carbohydrate composition of feed concentratates on methane emission from dairy cows and their slurry. Environ. Monit. Assess. 2005, 107, 329–350.
- Hindrichsen, I.; Kreuzer, M. High methanogenic potential of sucrose compared with starch at high ruminal pH. J. Anim. Physiol. Anim. Nutr. 2009, 93, 61–65.
- Castillo, C.; Benedito, J.; Méndez, J.; Pereira, V.; Lopez-Alonso, M.; Miranda, M.; Hernández, J. Organic acids as a substitute for monensin in diets for beef cattle. Anim. Feed. Sci. Technol. 2004, 115, 101–116.
- McAllister, T.; Newbold, C.J. Redirecting rumen fermentation to reduce methanogenesis. Aust. J. Exp. Agric. 2008, 48, 7–13.
- Beauchemin, K.; McGinn, S.M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006, 84, 1489–1496.
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253.
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.; Colombatto, D.; McAllister, T.A.; Beauchemin, K.A. A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed. Sci. Technol. 2008, 145, 209–228.
- Benchaar, C.; Greathead, H. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim. Feed. Sci. Technol. 2011, 166, 338–355.
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. In Annales de Zootechnie; EDP Sciences: Les Ulis, France, 2000; pp. 231–253.
- Lopez, S.; McIntosh, F.; Wallace, R.; Newbold, C.J. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Anim. Feed. Sci. Technol. 1999, 78, 1–9.
- McAllister, T.; Beauchemin, K.; Alazzeh, A.; Baah, J.; Teather, R.; Stanford, K. The use of direct fed microbials to mitigate pathogens and enhance production in cattle. Can. J. Anim. Sci. 2011, 91, 193–211.
- Newbold, C.J.; Rode, L. Dietary additives to control methanogenesis in the rumen. In International Congress Series; Elsevier: Amsterdam, The Netherlands, 2006; pp. 138–147.
- Beauchemin, K.; Colombatto, D.; Morgavi, D.; Yang, W.Z. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J. Anim. Sci. 2003, 81, E37–E47.
- Eun, J.-S.; Beauchemin, K.A. Assessment of the efficacy of varying experimental exogenous fibrolytic enzymes using in vitro fermentation characteristics. Anim. Feed. Sci. Technol. 2007, 132, 298–315.
- Hook, S.E.; Northwood, K.S.; Wright, A.D.; McBride, B.W. Long-term monensin supplementation does not significantly affect the quantity or diversity of methanogens in the rumen of the lactating dairy cow. Appl. Environ. Microbiol. 2009, 75, 374–380.
- Patra, A.K. Enteric methane mitigation technologies for ruminant livestock: A synthesis of current research and future directions. Environ. Monit. Assess. 2012, 184, 1929–1952.
- McGinn, S.; Beauchemin, K.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356.
- Guan, H.; Wittenberg, K.; Ominski, K.; Krause, D.O. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 2006, 84, 1896–1906.
- Clark, H.; Pinares-Patiño, C.; De Klein, C. Methane and nitrous oxide emissions from grazed grasslands. Grassland. A Glob. Resour. 2005, 279, 293.
- Madsen, J.; Lassen, J.; Hvelplund, T.; Weisbjerg, M.R. A fast, easy, reliable and cheap method to measure the methane production from ruminants. Eaap Publ. 2010, 127, 121–122.
- Eckard, R.; Grainger, C.; De Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livest. Sci. 2010, 130, 47–56.
- Sherman, R. Large-Scale Organic Materials Composting. North Carolina Cooperative Extension Service. 2005; North Carolina State Extension Publications: Raleigh, NC, USA, 2011.
- Kelly, F.; Michael, L. Storing Manure on Small Horse and Livestock Farms; The State University of New Jersey: Rutgers, NJ, USA, 2014.
- Ghaly, A.; MacDonald, K.N. An effective passive solar dryer for thin layer drying of poultry manure. Am. J. Eng. Appl. Sci. 2012, 5, 136–150.
- Endres, M.I.; Janni, K.A. Compost bedded pack barns for dairy cows. Univ. Neb.-Linc. 2008, 1, 1–9.
- Bewley, J.; Taraba, J.; Day, G.; Black, R.; Damasceno, F.A. Compost Bedded Pack Barn Design Features and Management Considerations; Cooperative Extension Publ. ID-206, Cooperative Extension Service; University of Kentucky College of Agriculture: Lexington, KY, USA, 2012.
- Aguirre-Villegas, H.; Larson, R.A.; Ruark, M.D. Solid-Liquid Separation of Manure and Effects on Greenhouse Gas and Ammonia Emissions; University of Wisconsin-Extension, Cooperative Extension: Madison, WI, USA, 2017; Available online: https://learningstore.extension.wisc.edu/products/solid-liquid-separation-of-manure-and-effects-on-greenhouse-gas-and-ammonia-emissions-p1844 (accessed on 23 October 2023).