1. Two-Dimensional (2D) OS Cell Models
Two-dimensional OS cell culture models, frequently used in vitro, have long been a conventional method for studying tumorigenesis, cancer biology, and drug discovery [
17]. These basic models not only aid in understanding the molecular and phenotypic characteristics of cells, but also facilitate hypothesis testing for translational research and the creation of genome–drug response correlations [
18]. The popularity of established cell lines is attributed to their practicality, cost-effectiveness, and speed in delivering experimental results.
Pioneering research from Mohseny’s laboratory has identified OS cell lines that exhibit key features of tumorigenesis, such as immune attraction (U2OS), angiogenesis (IOR/OS-14 and HOS-143B), the invasion of adjacent tissues (MHM), in vivo differentiation (IOR/OS9), and metastasis (HOS-143B) [
19]. These OS cell lines offer a broad range of tumorigenesis attributes, thus accelerating the drug discovery process [
19]. For instance, drug response assays with SAOS-2, U2OS, SJSA-1, HOS, and MNNG human OS cell lines have been instrumental in uncovering the therapeutic potential of compounds like afatinib [
20]. Afatinib was observed to inhibit OS cell viability, motility, and migration by suppressing the activation of the ErbB pathway [
20].
Further research has enriched the variety of available OS cell lines. Thanindratarn and colleagues unveiled a novel recurrent OS cell line, OSA 1777, which provided novel insights into the mechanisms of OS recurrence and metastasis [
21]. Similarly, VanCleave and team introduced a unique, enduring human cancer cell line, COS-33, which precisely mirrors the original tumor’s histopathology, cytogenetic intricacy, osteoblastic activity, and drug sensitivity [
22]. Notably, VanCleave’s research revealed that this cell line has a particular dependency on the mTOR pathway, a critical regulator of cell growth and proliferation [
22]. Such dependency is of high clinical relevance as there are already clinically approved drugs targeting this pathway [
22]. Consequently, COS-33 could serve as a new or complementary tool for drug screening, and for further elucidating OS dependencies on key signaling pathways like the mTOR pathway [
22].
Proteomic analysis reveals that established OS cell lines can partially depict primary tumors, demonstrating their significant value in illustrating tumor biology [
23]. However, these cell lines often exhibit systemic proteomic differences compared to the original tumors, reflecting variations in tumor stroma, extrinsic signaling, and growth conditions [
24]. Despite their easy manipulation, adaptability for global studies, and suitability for high-throughput applications, their questionable accuracy in reflecting clinical samples is a persistent concern [
25,
26].
The 2D cell lines bear inherent limitations, which include genetic homogeneity from in vitro selection, gene drift upon successive passaging, and a deficiency in authentically mimicking interactions between cancer cells and their microenvironment or reproducing patient treatment responses [
27,
28]. Furthermore, these models fall short in fully capturing the intricacy and pathophysiology of in vivo tumors [
29,
30,
31]. Despite these, 2D models remain essential. Their rich data have propelled the evolution of more advanced in vitro preclinical models and have corroborated previous findings in clinically relevant models.
2. Three-Dimensional (3D) OS Cell Models
Advancements in tissue engineering have led to the development of 3D constructs, such as spheroids and organoids, designed to more accurately replicate the complex intracellular dynamics and microenvironments of OS [
32,
33]. Spheroids are cellular aggregates embedded with collagen type I, with outer cells adhering to and invading into the matrix [
34]. These compact, globular structures can mimic diverse microenvironments within tumors, including anoxic, hypoxic, and oxic niches [
35]. Organoids are self-organized three-dimensional structures derived in vitro from pluripotent or adult stem cells [
9]. They create a microanatomy that closely resembles native tissue with differentiated cell types and organ-specific compartmentalization [
36,
37]. These 3D models, with their advanced tissue mimicry, present a promising platform for the advancement of personalized medicine. They can be expanded in vitro and subjected to various drug treatments to determine the most effective therapy for each individual patient. Based on the chosen preparation method, 3D models can be crudely classified into three categories: (i) scaffold-free sphere models, (ii) scaffold-based sphere models, and (iii) organoid models [
38,
39,
40,
41] (
Table 2 and
Figure 1).
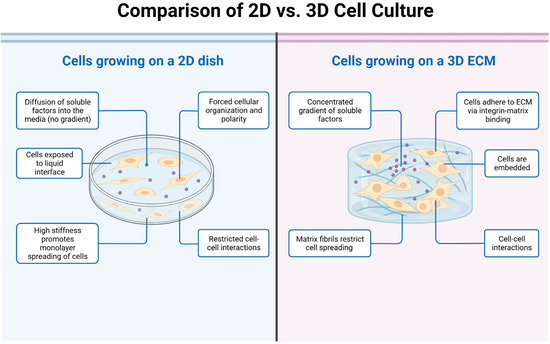
Figure 1. The picture illustrates the key differences between 2D and 3D cell cultures. In the 2D cell culture model, cells are grown and adhered to a flat surface, such as a petri dish or a culture flask. The cells form a monolayer and spread out in a single plane. In the 3D cell culture model, cells are grown in a three-dimensional environment that better mimics the natural tissue architecture. Cells can be encapsulated within hydrogels or scaffolds, allowing them to grow and interact in a more physiologically relevant manner.
Table 2. Three-dimensional in vitro models for osteosarcoma and drug discovery research.
3. Murine Models
OS murine models include xenografts and genetically engineered models. Xenograft models are characterized by the implantation of patient-derived OS cell lines into immunodeficient mice [
8]. This model maintains the heterogeneity of human tumors, providing an advantageous platform for the evaluation of therapeutic efficacy and the study of tumor–host interactions [
8]. Conversely, genetically engineered models, often utilizing specific oncogene alterations, present an ideal system for studying OS pathogenesis and progression [
81,
82]. Xenograft and transgenic mouse models have emerged as indispensable experimental systems, demonstrating exceptional proficiency in accurately replicating the intricate characteristics of OS in vivo [
83,
84] (
Figure 3).
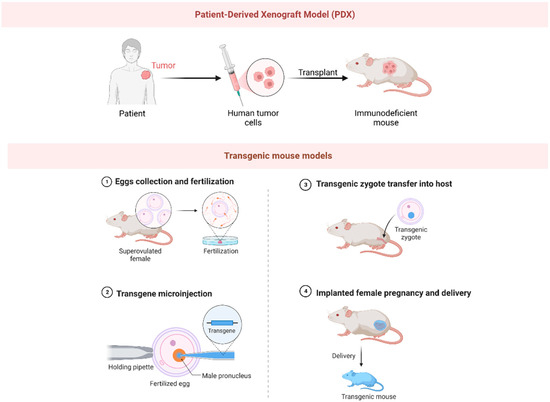
Figure 3. The picture illustrates the preparation methods for PDX (patient-derived xenograft) model and transgenic mouse model.
3.1. Xenograft Mouse Models
Xenograft models are predominantly categorized into two types: direct xenograft models (DXM) and cell-line-derived xenograft models [
85]. Currently, the PDX model, a specific type of DXM, is more extensively utilized in the investigation of OS therapeutics [
86]. Historically, the optimization of standard chemotherapeutic drugs, such as cisplatin, DOX, ifosfomide, and methotrexate, has been achieved through the utilization of PDX OS models [
87]. In recent years, an array of over 100 compounds has been subjected to rigorous screening via PDX models to ascertain their therapeutic efficacy against OS [
88]. One exemplary example is anticarin-β, a naturally derived coumarin compound extracted from the bark of Antiaris toxicaria Lesch [
89]. Utilizing tumor tissues procured from OS patients, researchers successfully established PDX models via subcutaneous transplantation into immunodeficient mice [
89]. The clinical potential of anticarin-β was subsequently evaluated utilizing these mouse PDX models [
89]. Remarkably, anticarin-β demonstrated potent inhibitory impacts across diverse stages of OS, notably including lung metastasis, in the PDX models [
89]. These promising outcomes suggest that anticarin-β may offer a viable therapeutic strategy for the management of OS, particularly in the context of advanced or metastatic cases [
89].
Despite bearing identical genomic modifications to their corresponding human tumors, PDX models inherently present certain constraints. In particular, the therapeutic response observed within these models does not invariably imply successful clinical trial efficacy [
83]. For instance, glembatumumab vedotin, an antibody–drug conjugate (ADC), and eribulin, a microtubule inhibitor, showed potential against OS in PDX models [
90]. However, their actual effectiveness in patients suffering from recurrent OS was found to be decidedly limited [
90,
91]. In the case of eribulin, the observed discrepancy likely stems from a failure to adequately consider the pharmacokinetic variations between mice and humans [
92]. One significant limitation is that PDX tumors must be implanted in immunodeficient mice, which results in these models falling short of reproducing the immunological intricacies of cancers and their treatments. This limitation is particularly noticeable when assessing the effectiveness of immunotherapies. Determining how activity levels in PDX models translate into clinical efficacy presents another challenge. The evaluation could be based on either the percentage of models demonstrating a response, or the intensity of the response within an individual model. Evaluating the predictive value of these preclinical models is complicated, particularly with the scarcity of novel agents that exhibit clinical activity, thereby constraining the derivation of reliable insights from these models.
3.2. Transgenic Mouse Models
Besides PDX models, various transgenic OS models have been developed, and yet their application in drug discovery remains notably infrequent [
8]. For example, Nannan et al. crafted a unique transgenic mouse model, wherein tumor protein p53 was specifically inactivated in osteoblasts [
93]. The study’s results revealed that inactivating p53 within osteoblasts led to an increase in local bone formation [
93]. This suggested a previously unexplored role for p53 within these cells, positioning it as a potential regulator of bone metabolism [
93]. The authors’ novel findings have critical implications for devising therapies for diseases with abnormal bone activity, such as osteoporosis and OS [
93]. Wang et al. delved into the intricate relationship between the S-phase kinase-associated protein 2 (SKP2) and cyclin-dependent kinase inhibitor 1B (p27) [
94]. Their groundbreaking study used a mouse model with Rb1 and Trp53 double knockouts within osteoblastic lineage cells [
94]. This investigation highlighted the profound effect of the SKP2-p27 interaction on OS’s progression and stemness [
94]. Their discovery suggests potential novel targets for therapeutic intervention, thereby expanding our understanding of OS’s complex molecular pathways [
94]. In a pivotal study, Ferrena et al. utilized mouse models deficient in Retinoblastoma 1 (Rb1) and Tumor Protein p53—two key genes in OS—to examine the effects of SKP2 knockout [
95]. Their results revealed that SKP2 deficiency induced significant immune infiltration within the tumor microenvironment, suggesting a potential immune response against OS [
95]. Further, the SKP2 knockout triggered a transcriptional program associated with a favorable prognosis [
95]. This crucial work, leveraging interactions within the tumor microenvironment, paves the way for novel osteosarcoma treatment strategies [
95]. These transgenic models provide the opportunity to assess OS within their native microenvironment, thereby addressing certain limitations associated with PDX models. Nonetheless, due to the dissimilarities between murine and human immune systems, transgenic models may not fully replicate immune reactions to OS in patients. To overcome this limitation, researchers have begun developing ‘humanized’ mouse models—PDX models of OS in immunocompromised mice reconstituted with human immune cells [
96]. However, this research field remains in its infancy, with relatively few models currently available. Nonetheless, it represents a promising frontier for osteosarcoma research, with the potential to revolutionize our understanding of the disease and our approach to its treatment.
4. Canine Models
Dogs represent a highly relevant model for studying human OS due to several compelling parallels. Just as in humans, OS is the most prevalent bone cancer in dogs, frequently manifesting in the long bones—a clinical feature consistently observed in both species [
97]. Furthermore, the clinical intervention process for osteosarcoma, which encompasses preoperative to postoperative procedures, exhibits a striking resemblance between canines and humans. This parallelism highlights the significance of the canine model in enhancing the comprehension of osteosarcoma, and in the development of therapeutic approaches [
97]. A unique aspect that highlights the relevance of the canine model is that, apart from humans, dogs are the only mammals known to spontaneously develop OS within the context of an intact immune system [
98]. These marked similarities not only highlight the dog as a powerful model for understanding the biology and clinical progression of OS, but also emphasize its potential in advancing novel therapeutic approaches for OS.
Recent investigations employing the canine model have opened promising pathways for the development of innovative pharmaceutical treatments in OS. Canine OS cell lines have proven to be a vital resource in the field of drug discovery. In their research, Chirio et al. used these cell lines to evaluate how well DOX-loaded, calcium phosphate-coated lipid nanoparticles worked. Their laboratory results highlighted the promise of these particles in overcoming drug resistance and boosting the effects of chemotherapy [
99]. Similarly, Yang et al. investigated the synergistic effects of sorafenib and DOX in both human and canine OS cell lines [
100]. Their findings revealed that the combination of these two drugs exhibited enhanced efficacy in inhibiting cell proliferation, reducing migration and invasion abilities, and inducing cell cycle arrest [
100]. The in vivo canine OS model provides a valuable tool for studying drug behavior within a complex physiological context. A study by Regan et al. investigated the efficacy of losartan, a drug commonly used to treat hypertension, in combination with the kinase inhibitor toceranib, in the treatment of metastatic OS in 28 dogs [
101]. They demonstrated that losartan effectively blocked the recruitment of monocytes elicited by OS, and, when combined with toceranib, resulted in significant clinical benefits in dogs with metastatic OS [
101]. These results hold significant implications for OS drug development, suggesting a potential therapeutic strategy that could improve treatment outcomes for both human and canine patients.
However, it is imperative to acknowledge certain limitations associated with using the canine OS model for drug development. Firstly, significant differences might exist between canines and humans in the pharmacokinetic and pharmacodynamic profiles of drugs due to species-specific metabolic processes [
102]. This could potentially create discrepancies in drug efficacy and safety assessments [
102]. Additionally, ethical considerations concerning animal welfare in experimental settings must be strictly addressed, which may limit the scope and application of certain investigational procedures [
103]. Thus, while the canine model provides crucial insights for OS drug development, it is essential to balance its use with complementary models and strategies to ensure comprehensive and accurate results.
This entry is adapted from the peer-reviewed paper 10.3390/biom13091362