Milk is a complex food produced by the mammary gland, providing a baby with essential nutrients for growth along with an immunity kit for short-term survival and long-term epigenetic information influencing the health of the future adult. During lactation, the composition of milk is tuned to the baby’s needs from colostrum at birth up to the end of lactation at weaning [
1]. Milk from different mammals can be used to feed a human baby, suggesting that molecules like miRNAs—known to retain biological activity from flies to mammals—can easily be tested on a relevant animal model for potential preventive use in the nutritional therapy of infants. Many ribo- and desoxyribonucleic acids of complex food are recycled by the digestive system. However, some escape digestion and are used in cell-to-cell signaling or are provided by immune or exfoliated epithelial cells directly to the offspring. A mature miRNA is a single-stranded ribonucleic acid of 19–25 nucleotides in length, which is generated by the RNase-III-type enzyme Dicer from an endogenous transcript that contains a local hairpin structure [
2,
3]. The miRNAs are encoded either within the introns of protein-coding genes or transcribed under the control of their own promoters. After transcription as long primary transcripts, they are trimmed into hairpin intermediates (pre-miRNAs) in the nucleus, subsequently exported to the cytoplasm, then cleaved into mature miRNAs. The hairpins usually code for 3p and 5p mature miRNAs with differential tissular expressions [
4]. The miRNAs function as post-transcriptional repressors of their target genes when bound to specific sites in the 3′ untranslated region (UTR) of the target mRNA. The binding of the miRNAs relies on ‘seed pairing’, i.e., the perfect or near-perfect complementary match of nucleotides 2–8 of the mature miRNA product [
5]. In silico database exploration has predicted binding sites on the promoter 5′-UTR coding domain of mRNA as well as on lnc-RNA. The equivalence of let-7a binding sites in 5′-UTR or 3′-UTR for repression has been demonstrated by transfecting HeLa cells [
6]. miRNAs can be detected in the cytoplasm, nucleus, nucleolus [
7], and mitochondria [
8]. Noncoding ribonucleic acids (small or long ncRNA) are involved in epigenetic regulation. These molecules directly silence or activate chromatin at specific loci or through their integral role in the machinery that drives DNA methylation. Breast milk contains miRNAs in high amounts [
9]; these have been proposed to act as epigenetic regulators [
10,
11,
12,
13].
2. RNA Content and Mature miRNA Diversity
The miRNA composition of breast milk has been explored by RNA sequencing performed with kits designed with an RNA extraction step [
14] or without [
15], as in a recent breast milk cohort [
16]. q-PCR is used for the specific exploration of known miRNAs and confirmation studies, with normalization provided by reference endogenous genes like miRNAs or by spiking samples with xenogenous miRNA like cel-lin4-5p.
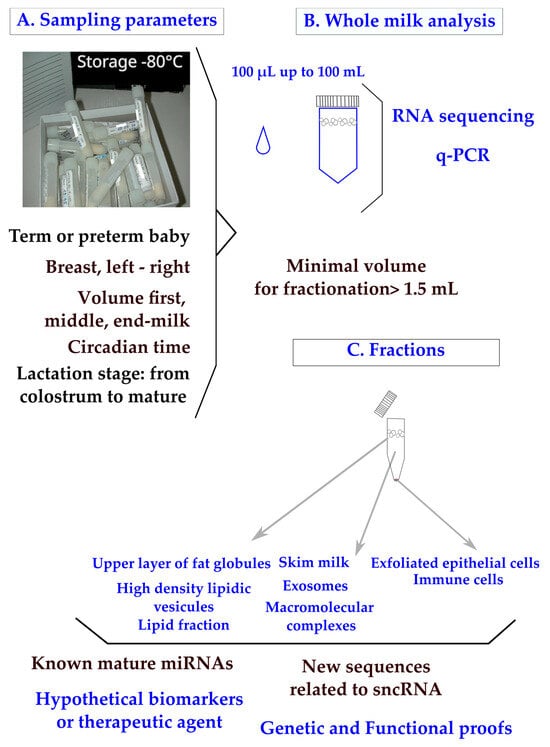
In clinical practice, breast milk is classified as a noninvasive sample available in maternity wards or collected at home on a worldwide scale. The main parameters to ponder when designing a clinical plan are the mother’s delivery (term or preterm), the capacity of mammary glands to deliver the food (difference between breasts; the compositional variation during suckling between fore, middle, and hind milk; and the circadian rhythm of production) (
Figure 1A). To control for the differences between breasts, mothers are sometimes asked to use the same breast for sampling at each time-point. Minimal differences in miRNA content have been found between fore and hind milk [
17]. It has been advised that pre-feed samples should be utilized in order to minimize confounding [
18]. Minimal impacts of freeze–thaw cycles have been found on milk miRNAs [
19,
20], which is not surprising as the milk of domestic animals retains cryoprotective properties. All these parameters have to be taken into account when designing a sampling procedure [
21].
In clinical trials, available volumes varying between 100 µL and 100 mL have been reported (
Figure 1B). Although q-PCR is a sensitive method, the design of a breast milk analysis using 100 µL (down to 50 µL [
20]) is challenging, but it is a prerequisite when exploring milk samples from the same mother during a nychthemeron or when sharing samples between multiple analyses (transcriptomic, lipidomic, and metabolomic [
22]). We have to consider that the sampling of a single drop of breast milk clearly has less informative value than a volume of over 5 mL. Multiplexed techniques to allow bulk extractions from low volumes of crude milk are needed in the field.
The RNA content of whole breast milk after classical phenol/chloroform extraction is wide; for instance, 90 to 1000 ng/µL from 8 parturients [
23]. Breast milk contains a high amount of small or very small noncoding RNAs [
24] along with transfer RNA, messenger RNA, and long noncoding RNA [
25,
26].
Figure 1. Human breast milk composition: sampling parameters, analyses of whole milk, and fractions. Sampling parameters of whole milk (
A) were explored using RNA sequencing or q-PCR. (
B) Clinical samples pose a major problem of legal pressure to reduce the sample volume. Milk fractions have been explored down to 1 mL [
27] and 1.5 mL of whole milk [
19]. (
C) Recommendations have been made to centrifuge breast milk immediately after sampling to remove exfoliated cells, then to store them at −80 °C or, preferably, under liquid nitrogen. A step at −20 °C is frequently applied when mothers are sampled at home.
In the lipid fraction of breast milk (
Figure 1C), the variations in known and newly described miRNAs have been explored using mothers with normal diets [
18,
28,
29] or comparatively with mothers consuming high-fat diets [
30]. Many short RNAs are undescribed and, beyond as an interest as biomarkers, new regulatory processes may be discovered when exploring the total content of whole milk. The classification of miRNAs is designed on biogenesis [
2,
31] with four criteria: (1) the confirmation of expression by hybridization to a size-fractionated RNA sample; (2) the small RNA sequence has to be present in one arm of the hairpin precursor, which lacks large internal loops or bulges; (3) the small RNA sequence has to be phylogenetically conserved and the sequence conservation has to be seen in the precursor hairpin, but to a lesser extent than in the mature miRNA segment; and (4) the evidence is stronger if the precursor accumulates in the presence of a reduced Dicer function. More recently, the definition has been reassessed to distinguish the function from the biogenesis of miRNAs from other classes of RNA by dissociating the three notions of an miRNA gene (Mir), an miRNA precursor (pre-MiRNA), and a mature miRNA product (MiRNA). Under that assumption, miRNA genes produce mature miRNAs, but some miRNAs may originate from genes transcribed into other types of noncoding RNAs [
32]. These definitions are of importance when performing RNA sequencing with the aim of discovering new miRNAs.
The recovery of exosomes is influenced by the procedure of sampling (
Figure 1C) and a systematic centrifugation step of the milk before storage has been proposed in order to remove cells and debris [
33]. A minimal volume of 1.5 mL whole milk has been used to purify exosomes [
19], which is a technical lock, up to higher volumes like 20 mL [
34].
The diversity of miRNAs has been related to the stage of lactation. A strikingly different composition of miRNAs between colostrum and mid-lactation from 18 parturients has been found (seven miRNAs were confirmed by q-PCR: miR-511-3p, 429, 29c-3p, 885-5p, 30b-5p, 183-5p, and 623 [
35]). Previously, a different composition of miRNAs comprising one colostrum against five breast milk has been reported. miR-518c-3p miRNAs is uniquely detected in breast milk [
9] and is also expressed in the placenta with a putative use as a preeclampsia biomarker [
36]. Seven miRNAs (miR-148a-3p, 22-3p, 26a-5p, 21-5p, 7b-5p, 7g-5p, and 24-3p) have been found to be common to nipple aspirate, serum, plasma, breast tissue, and breast milk [
37]. Surprisingly, the lists have no common miRNAs; this is notable in the let-7 family, which is highly expressed in tissues [
37] and used as reference gene [
20]. let-7g miRNAs are considered to be a reliable molecular method for body-fluid identification markers at crime scenes [
38]. It should be underlined that the levels of microRNAs of the let-7 family in exosomes from mothers suffering from type 1 diabetes are widely divergent from normal levels [
39].
hsa-miR-26a-5p MIMAT0000082, UUCAAGUAAUCCAGGAUAGGCU, is highly fsimilar to 26b-5p, just like hsa-miR-26a-1-3p MIMAT0004499, 5′-CCUAUUCUUGGUUACUUGCACG-3, is highly similar to 26a-2-3p and 26b-3p. The family is widely expressed in mammals [
50].
Up to now, the number of studies on the composition of noncoding RNA is relatively modest according to the population of women of reproductive age, ethnic diversity, and the diets in use on a world scale. The consequence is that discrepancies between studies cannot properly be explained according to the huge diversity of environmental factors like food or mother’s ethnicity. Due to the high connectivity of miRNAs, new integrative strategies between omics and the clinical parameters of the mother have to be designed to evidence existing logical links. The studies on miR-26 and miR-320 families illustrate that more work under the redundancy hypothesis of action of near-identical molecules or paralogs are needed for proper therapeutic use. The next section presents the data available on the conditions of miRNA bioavailability in the digestive tract.
3. Bioavailability of Mature miRNAs in the Digestive System
The systemic RNA interference–deficient transporter (sidt1) is the main receptor of dietary and orally administered miRNAs in the digestive system [
53], indicating that miRNAs present in breast milk can transfer to offspring. However, miR-375-3p cannot cross the digestive tract of mice [
54]; even when using the breast milk of a mouse engineered to produce a high amount of miR-30b, a demonstration of plasma loading was impossible [
55].
As a consequence, the bioavailability of miRNA depends on sheltering these molecules from the molecular environment. It determines the capacity of miRNA molecules to reach target cells with a concentration high enough to trigger a biological response, a crucial step in digestive fluids highly loaded with RNases. In vitro data on cell cultures have shown that a ratio of 100 miRNA molecules delivered to the target cell cytoplasm triggers a measurable physiological response [
56]. In vivo, after a rough estimation of the total cells of a rat gastric mucosa to adapt to a concentration of miR-320-3p or miR-375-3p, we were able to confirm that this ratio could be used in an oral gavage [
57,
58].
Milk is rich in exosomes, which can serve as natural cargo for miRNAs [
59]. However, these molecules are frequently widely present in all milk fractions. Thus, fluorescently labeled miRNAs like miR-375-3p must be transfected in exosomes in order to show accumulations in the liver, spleen, and brain after suckling or oral gavages. A demonstration was conducted using mice and by transfecting bovine exosomes with fluorescent miRNA administered to mice [
60]. Another approach is to take advantage of the milk composition related to a certain pathology of the mother. Human milk exosomes from gestational diabetes mellitus (GDM) and healthy parturients showed distinct regulatory bioactivities in both HepG2 cell cultures and, in vivo, in the liver of Balb/c mice. The profile of GDM exosomes has been related to natural loading by miR-101-3p [
34]. Previous studies confirmed that the target gene of miR-101-3p is mTOR. miR-101-3p suppresses mTOR by binding in the 3′-UTR regions of mRNAs [
61].
Works on milk exosomes are a very active field of investigation because (1) a food source of a single miRNA species to supplement diets with a crude product does not exist; (2) exosomes can protect RNA from digestive enzymes; and (3) they can be tailored by the genetic engineering of cell lines to load miRNAs with molecular data, addressing relevant cells or tissues [
62]. Some natural miRNAs can be addressed to exosomes [
63] with a step of loading the miRNA onto Argonaute proteins before loading into exosomes [
64].
However, miRNA obtained by chemical synthesis can also be loaded into artificial vectors like lipoaminoglycoside Dioleyl-Succinyl Paromomycin [
65]. The loading of miR-375-3p has been measured using a transgenic rat model on the enteroendocrine cell lineage [
58]. Taking this specific miRNA as an example, miR-375-3p has been proposed as a key regulator in malignant breast cancer [
66]. In cohorts of breast-fed infants, the consumption of miR-375-3p was associated with protection from atopy [
67], opening the consideration of the prevention of allergic diseases through breastfeeding. The levels of miR-375-3p were found to be upregulated in the breast milk of mothers treated with probiotics, although the study concluded that this miRNA and others could not be considered to convey protection against allergic diseases to the baby [
68]. These divergent properties illustrate the problem caused by supplying miRNA in an animal model; most of the time, the site of delivery impacted different cellular phenotypes. Another level of complexity is added if one considers that the immune cells involved in allergy may be also stimulated by miRNAs from plants present in breast milk [
23]. As the chemistry of plant miRNAs is different from eucaryotes, complicating the analysis of breast milk, the presence of these xenogenous molecules has to be taken into account in future allergy studies [
69].
The oral administration of miRNAs is a current problem in targeted nutritional therapy and requires further studies, both to design new prokaryotic or eukaryotic vectors [
70,
71] and to test miRNA cocktails. A specific problem with models of rodent babies is to ensure that the stomach is empty by separating the pups from their mother in one hour before gavage [
57,
58]. The biological relevance of miRNAs in low amounts is counterintuitive as a high abundance frequently correlates with a high bioactivity [
72]. But, the continued uptake of milk-derived exosomes that carry dnmt-targeting miRNAs may promote diabetes, allergies [
69], neurodegenerative diseases, and cancer later in life [
11].