The overuse and misuse of antibiotics have led to the emergence and spread of multidrug-resistant (MDR), extensively drug-resistant (XDR), and pan-drug-resistant (PDR) bacteria strains, usually associated with poorer patient outcomes and higher costs. In order to preserve the usefulness of these life-saving drugs, it is crucial to use them appropriately, as also recommended by the WHO. Moreover, innovative, safe, and more effective approaches are being investigated, aiming to revise drug treatments to improve their pharmacokinetics and distribution and to reduce the onset of drug resistance. Globally, to reduce the burden of antimicrobial resistance (AMR), guidelines and indications have been developed over time, aimed at narrowing the use and diminishing the environmental spread of these life-saving molecules by optimizing prescriptions, dosage, and times of use, as well as investing resources into obtaining innovative formulations with better pharmacokinetics, pharmacodynamics, and therapeutic results. This has led to the development of new nano-formulations as drug delivery vehicles, characterized by unique structural properties, biocompatible natures, and targeted activities such as state-of-the-art phospholipid particles generally grouped as liposomes, virosomes, and functionalized exosomes, which represent an attractive and innovative delivery approach.
- antimicrobial resistance
- antibiotics
- phospholipids
1. Introduction
1.1. Antibiotics and Chemo-Resistance: A Worrying Phenomenon
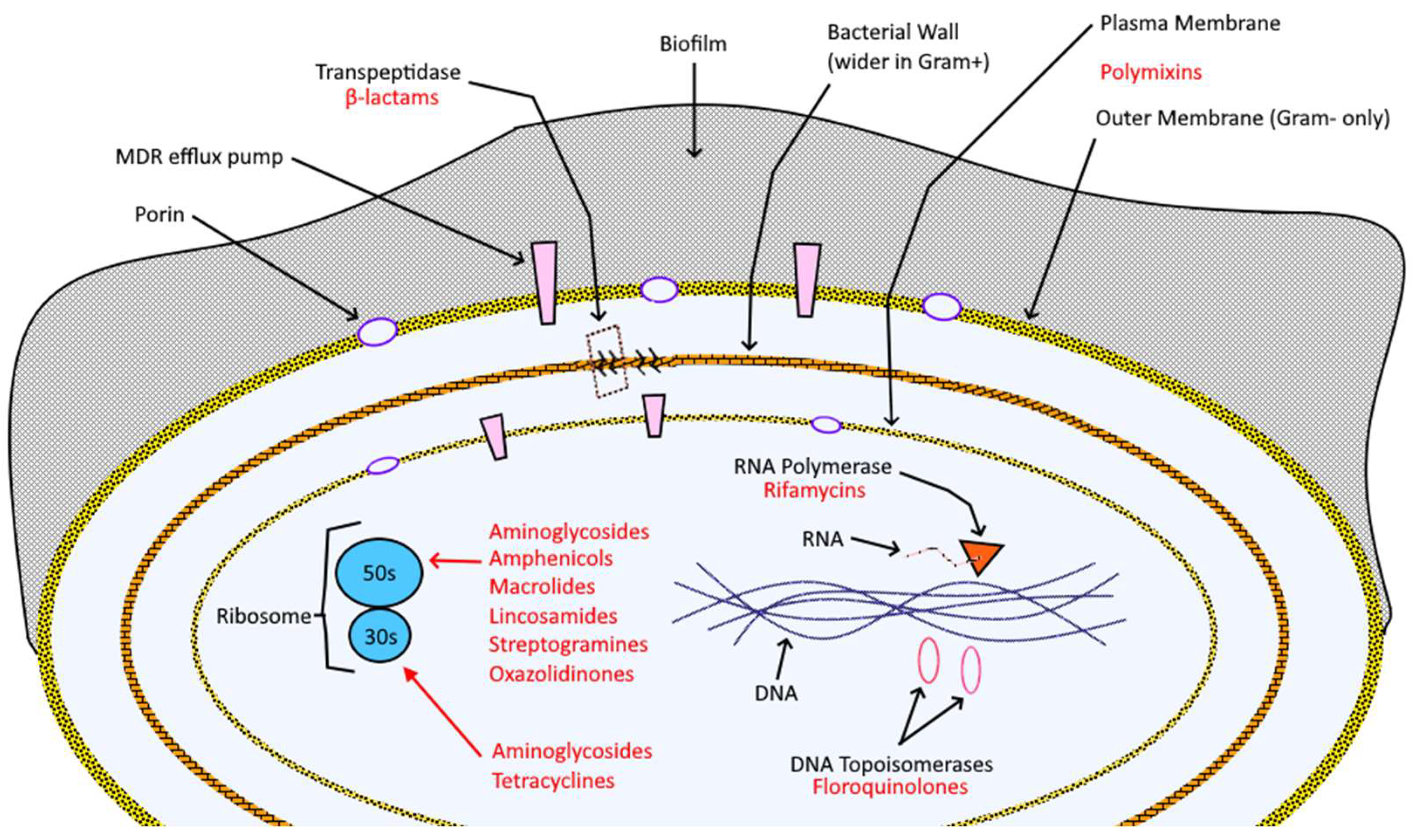
Table 1. Specific mechanism of action (M.O.A.) and chemo-resistance mechanisms (C.M.) per class of antibiotic, with references.
|
Articles |
C.M. |
M.O.A. |
Antibiotic |
|
[12,13] |
β-lactamases (minor for Cephalosporins) |
Inhibition of DD-TP and PBPs |
Penicillins and Cephalosporins |
|
[14–16] |
Carbapenemases, target site mutation and efflux pumps |
Inhibition of DD-TP and PBPs |
Carbapenems, Monobactams and Penems |
|
[17,18] |
Target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation |
Bond to 30S ribosomal subunit (sterical clash on A-site) |
Tetracyclines |
|
[19,20] |
Target site mutation (methylation/de-methylation), membrane non-permeability, efflux pumps, enzymatic degradation |
Bond to 30S–50S ribosomal subunits (link to h44–H69 sites, prevent translocation) |
Aminoglycosides |
|
[21,22] |
Target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation |
Bond to 50S ribosomal subunit (prevent peptide-bond formation on Cam1 (eu) and Cam2 (archea)) |
Amphenicols |
|
[23–25] |
Translation of specific “bottle brush” oligopeptides, bypass synthesis, target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation |
Bond to 50S ribosomal subunit (block ribosomal exit tunnel) |
Macrolides |
|
[26–28] |
Target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation |
Bond to 50S ribosomal subunit (sterical impedance on A-site at peptidyl-transferase center) |
Lincosamides |
|
[29–31] |
Translation of specific “bottle brush” oligopeptides, bypass synthesis, target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation |
Bond to 50S ribosomal subunit (block ribosomal exit tunnel on specific, synergistic sites) |
Streptogramins |
|
[32–34] |
Target site mutation (methylation/de-methylation), membrane non-permeability, enzymatic degradation, PoxtA- and OptrA-mediated resistance |
Bond to 50S ribosomal subunit (bind to A-site, acting as initiation inhibitor) |
Oxazolidinones |
|
[35,36] |
Mutations in rpoB encoding the β subunit of RNAP, inactivation by ADP-ribosylation, and other enzymatic degradations |
Inhibition of DNA-dependent RNA synthesis (binding to prokaryotic RNA polymerases) |
Rifamycins |
|
[37–39] |
Target site mutation (quinolone-resistance-determining regions), efflux pumps |
Inhibition of bacterial DNA synthesis (blockage of two DNA bacterial topoisomerases) |
Fluoroquinolones |
|
[40,41] |
Addition of cationic groups to LPS moieties, two-component signal transduction system enhancement, plasmid-encoded resistance determinant MCR-1 |
Disruption of bacterial membrane, inhibition of bacterial respiration |
Polymixins |
From the engineering and agro-industrial point of view, removing pharmaceutics and antibiotic resistance genes (ARGs) from water, soil, animal feed, and manure would greatly help reduce antibiotic resistance. This feature can be achieved by combining metal-based filtering systems [29][30], biological and biotechnological agents [31][32], and better maintenance and management [33][34]; a remarkable, somewhat different example in the field is brought up by the use of cultivated meat, which not only drastically reduces CO2, biological, and chemical waste with respect to farms but also almost deletes the need for antibiotics [35][36].
Last but not least, awareness campaigns [37][38], surveys [39], and technological integration in work [40][41] and learning [42][43] environments are crucial to stop an otherwise announced disastrous, unstoppable plague: communication is the key to unified, efficacious actions, especially when the enemy to defeat is so protean, diffused, and challenging.
1.2. Innovative Phospholipid Carriers (IPCs)
Lipid nanoparticles are one of the most promising vehicles of the family thanks to their elevated degree of biocompatibility, encapsulation efficiency, and customization [47]. Among them, the two types frequently used for the treatment of microbial infections are solid lipid nanoparticles and nanoliposomes [48][49]; Researchers will discuss the main features and some of the technological evolutions of the latter, namely functionalized liposomes and exosomes, under the label of innovative phospholipid carriers (IPCs).
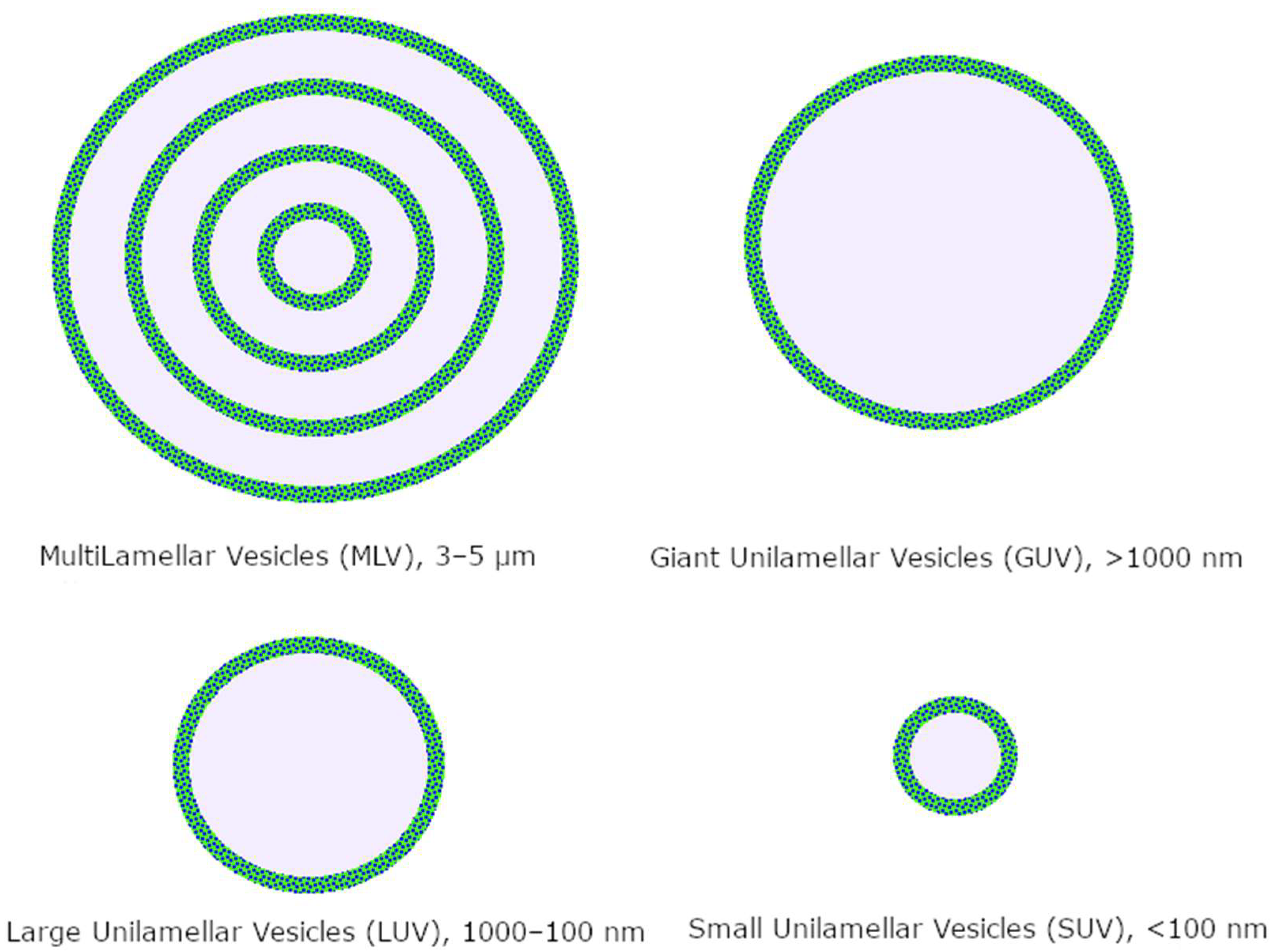
1.2.1. Liposomes
1.2.2. Functionalized Liposomes
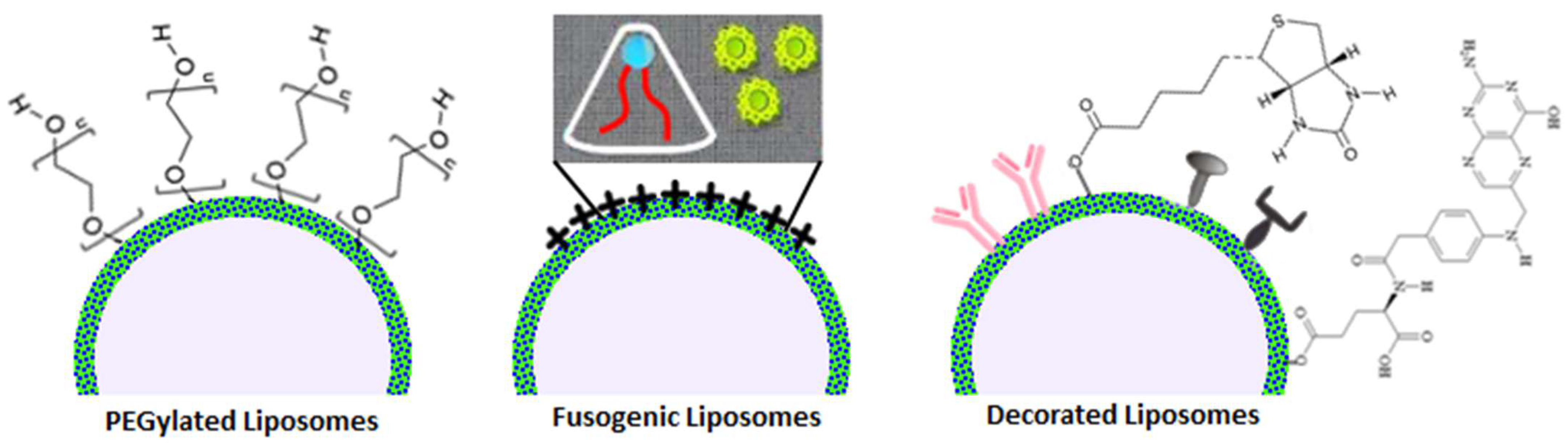
1.2.3. Exosomes
Extracellular vesicles (EVs) are subcellular structures delimited by a lipid bilayer and shed by cells into their surrounding environment. These vesicles are involved in cell-to-cell communication and play important roles in various physiological and pathological processes. They are further categorized in the following three main types of extracellular vesicles:
-
Exosomes are the smallest type of EVs, typically ranging in size from 30 to 200 nanometers. They originate from the endocytic pathway within the cell. Exosomes contain a diverse array of biomolecules, including proteins, nucleic acids (such as RNA and DNA), and lipids.
-
Microvesicles are larger than exosomes, typically ranging from 100 to 1000 nanometers in size. They are formed by the outward budding of the cell's plasma membrane and contain a similar assortment of biomolecules, serving as carriers for intercellular communication.
-
Apoptotic bodies are the largest of the extracellular vesicles, typically exceeding 1000 nanometers in size. They are produced during the process of cell apoptosis (programmed cell death) and contain cellular debris and organelles from the dying cell.
These three subtypes vary in size, function, biological origin, and other attributes.
Exosomes, which are the smaller extracellular vesicles (EVs), are accumulated into multi-vesicular bodies (MVBs) before secretion. Exosome are shed by a wide array of cell types, including dendritic cells (DCs), lymphocytes, epithelial cells, endothelial cells, neurons, etc. They are detectable in an extensive range of bodily fluids, including blood, urine, saliva, amniotic fluid, breast milk, hydrothoracic fluid, ascitic fluid, and the culture medium of most cell types [64]. Various factors can induce the release of exosomes: microbial agents, external stimuli, and various stressors can elicit these responses [65]. Exosomes encompass sugars, lipids, proteins, nucleic acids, and bioactive substances within the extracellular matrix. Initially, their role was believed to be the removal of metabolic waste [66].
The composition of exosomes differs depending on their source cell (Figure 1). So far, research has identified nearly 350,000 proteins, 40,000 nucleic acids, and 600 lipids in various exosomes [67]. This extensive flexibility offers numerous possibilities for practical clinical diagnosis and treatment scenarios involving exosomes, since the contain a wide range of proteins, including transmembrane proteins, lipid-anchored membrane proteins, peripherally adsorbed membrane proteins, and soluble proteins within the exosome lumen [68]. The most commonly found proteins in exosomes include membrane transporters and fusion proteins (e.g., GTPases, annexins, and flotillin), heat shock proteins (e.g., HSC70), tetraspanins (e.g., CD9, CD63, and CD81, which are typically considered exosome markers), proteins involved in multi-vesicular body (MVB) biogenesis (e.g., Alix and TSG101), and lipid-related proteins and phospholipases . Some proteins are recognized as specific markers of exosomes, with CD63 and CD81 tetraspanins being the most commonly used ones. Noteworthy, exosomes are also rich in lipids, primarily cholesterol, sphingolipids, phospholipids, and bisphosphates [69].
Exosomes are flexible enough to carry a variety of nucleic acids such as RNAs (mRNA, microRNA, and other non-coding RNAs) and DNAs (mitochondrial DNA, double-stranded DNA, single-stranded DNA, and viral DNA), suggesting that exosomes could act as carriers of genetic information [70]. Despite many of the RNAs in exosomes being degraded fragments with lengths of less than 200 nucleotides, some full-length RNAs may be present and delivered to recipient cells through endocytosis, potentially influencing protein production in those cells. In this context, exosomal miRNAs are also associated with specific diseases [71].
Recent attention has focused on exosomes as an auspicious drug delivery system. Their inherent biocompatibility, efficient delivery mechanisms, and minimal immunogenicity have elevated their standing. Extensive research has revealed the role of exosomes in mediating intercellular communication and participating in various physiological and pathological processes in the body. Their functions span a wide spectrum, encompassing immune responses, antigen presentation, cell migration, differentiation, angiogenesis, inflammation induction, apoptosis, atherosclerosis, tumor development, invasion, metastasis, and drug resistance. The capacity of exosomes to transport bioactive substances holds great potential for deciphering an enigma of diseases such as cancer, neurological disorders, cardiovascular conditions, and metabolic disorders, as well as for disease diagnosis using biomarkers [65][72].
2. Innovative Phospholipid Carriers versus Antimicrobial Resistance
3. Conclusions and Future Perspectives
The aim is to underline the important synergy between non-/conventional antibiotics and IPCs in order to fight chemo-resistance phenomena properly, to achieve better targeting, and to reduce a plethora of annoying side effects; even if this research field is thriving, with some notable examples of pharmaceutical formulations which successfully hit the market, the necessity of carrying on more studies and experimentations on IPCs specifically tailored to carry antibiotics and other essential antimicrobial molecules is clearly inferable from the scientific evidence collected in this work. However, this alone obviously cannot be the solution to a complex problem such as AMR, which indeed needs to be approached in a multi-strategical and multidisciplinary way: starting from the conventional therapies, multi-therapy protocols should be properly adopted in conjunction with precise antibiograms, pondered dosages of antibiotics, and molecular diversification to reduce, at minimum, the risk of resistance derived by nosocomial and community misusage. The employment of antimicrobial peptides and other molecules (i.e., oligonucleotides, metals, etc.) that naturally do not trigger chemo-resistance mechanisms can definitely help reduce the burden on conventional antibiotics; other carriers like solid lipid nanoparticles (SLN) and nanostructured lipid carriers (NLC), and innovative biocarriers such as bacteriophages can offer additional solutions to formulation, chemical stability, and delivery problems; last but not least, the strong need for more and more social awareness in communities, hospitals, and industries regarding correct handling of antibiotics and the strong impact that multidrug-resistant superbugs (MSRBs) are already exerting on our health and environment should be communicated at local, national, and international levels.
To sum up, this research wants to be an additional voice in the already loud, worried chorus about the rampage of microbial resistance and its dangerous outcomes: if common action is not taken properly starting from the present, the world will experience pandemics much worse than the recent COVID-19 outbreak, as claimed by the World Health Organization (WHO), which predicts 5.2 million due to AMR deaths in the Western Pacific alone by 2030. In this scenario, IPCs are a well-known, precious weapon against AMR that should be implemented more, given the promising results it produced over time.
This entry is adapted from the peer-reviewed paper 10.3390/ijms242115934
References
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial Resistance: A Global Emerging Threat to Public Health Systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2857–2876.
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6, 521–547.
- Subramaniam, G.; Girish, M. Antibiotic Resistance—A Cause for Reemergence of Infections. Indian J. Pediatr. 2020, 87, 937–944.
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Antimicrobial Resistance Collaborators; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655.
- Ukuhor, H.O. The Interrelationships between Antimicrobial Resistance, COVID-19, Past, and Future Pandemics. J. Infect. Public Health 2021, 14, 53–60.
- Rizvi, S.G.; Ahammad, S.Z. COVID-19 and Antimicrobial Resistance: A Cross-Study. Sci. Total Environ. 2022, 807, 150873.
- Neu, H.C.; Gootz, T.D. Antimicrobial Chemotherapy. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 978-0-9631172-1-2.
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular Mechanisms of Antibiotic Resistance Revisited. Nat. Rev. Microbiol. 2023, 21, 280–295.
- Stogios, P.J.; Savchenko, A. Molecular Mechanisms of Vancomycin Resistance. Protein Sci. Publ. Protein Soc. 2020, 29, 654–669.
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-Lactam Antibiotics: An Overview from a Medicinal Chemistry Perspective. Eur. J. Med. Chem. 2020, 208, 112829.
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance—The Need for Global Solutions. Lancet Infect. Dis. 2013, 13, 1057–1098.
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Mol. J. Synth. Chem. Nat. Prod. Chem. 2018, 23, 795.
- Nguyen, A.Q.; Vu, H.P.; Nguyen, L.N.; Wang, Q.; Djordjevic, S.P.; Donner, E.; Yin, H.; Nghiem, L.D. Monitoring Antibiotic Resistance Genes in Wastewater Treatment: Current Strategies and Future Challenges. Sci. Total Environ. 2021, 783, 146964.
- Junaid, M.; Liu, S.; Liao, H.; Liu, X.; Wu, Y.; Wang, J. Wastewater Plastisphere Enhances Antibiotic Resistant Elements, Bacterial Pathogens, and Toxicological Impacts in the Environment. Sci. Total Environ. 2022, 841, 156805.
- Farr, B.M.; Salgado, C.D.; Karchmer, T.B.; Sherertz, R.J. Can Antibiotic-Resistant Nosocomial Infections Be Controlled? Lancet Infect. Dis. 2001, 1, 38–45.
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, e00001-19.
- Yang, L.; Luo, X.; Sun, J.; Ma, X.; Ren, Q.; Wang, Y.; Wang, W.; He, Y.; Li, Q.; Han, B.; et al. The Antimicrobial Potential and Aquaculture Wastewater Treatment Ability of Penaeidins 3a Transgenic Duckweed. Plants 2023, 12, 1715.
- Zhang, L.-M.; Yang, M.; Zhou, S.-W.; Zhang, H.; Feng, Y.; Shi, L.; Li, D.-S.; Lu, Q.-M.; Zhang, Z.-H.; Zhao, M. Blapstin, a Diapause-Specific Peptide-Like Peptide from the Chinese Medicinal Beetle Blaps rhynchopetera, Has Antifungal Function. Microbiol. Spectr. 2023, 11, e0308922.
- Shang, C.; Ye, T.; Zhou, Q.; Chen, P.; Li, X.; Li, W.; Chen, S.; Hu, Z.; Zhang, W. Genome-Wide Identification and Bioinformatics Analyses of Host Defense Peptides Snakin/GASA in Mangrove Plants. Genes 2023, 14, 923.
- Lima, R.M.; Rathod, B.B.; Tiricz, H.; Howan, D.H.O.; Al Bouni, M.A.; Jenei, S.; Tímár, E.; Endre, G.; Tóth, G.K.; Kondorosi, É. Legume Plant Peptides as Sources of Novel Antimicrobial Molecules Against Human Pathogens. Front. Mol. Biosci. 2022, 9, 870460.
- Lima, W.G.; de Lima, M.E. Therapeutic Prospection of Animal Venoms-Derived Antimicrobial Peptides against Infections by Multidrug-Resistant Acinetobacter Baumannii: A Systematic Review of Pre-Clinical Studies. Toxins 2023, 15, 268.
- Blaskovich, M.A.T.; Hansford, K.A.; Gong, Y.; Butler, M.S.; Muldoon, C.; Huang, J.X.; Ramu, S.; Silva, A.B.; Cheng, M.; Kavanagh, A.M.; et al. Protein-Inspired Antibiotics Active against Vancomycin- and Daptomycin-Resistant Bacteria. Nat. Commun. 2018, 9, 22.
- Nguyen, T.H.T.; Nguyen, H.D.; Le, M.H.; Nguyen, T.T.H.; Nguyen, T.D.; Nguyen, D.L.; Nguyen, Q.H.; Nguyen, T.K.O.; Michalet, S.; Dijoux-Franca, M.-G.; et al. Efflux Pump Inhibitors in Controlling Antibiotic Resistance: Outlook under a Heavy Metal Contamination Context. Molecules 2023, 28, 2912.
- Masoudi-Sobhanzadeh, Y.; Pourseif, M.M.; Khalili-Sani, A.; Jafari, B.; Salemi, A.; Omidi, Y. Deciphering Anti-Biofilm Property of Arthrospira Platensis-Origin Peptides against Staphylococcusaureus aureus. Comput. Biol. Med. 2023, 160, 106975.
- Haque, M.A.; Marathakam, A.; Rana, R.; Almehmadi, S.J.; Tambe, V.B.; Charde, M.S.; Islam, F.; Siddiqui, F.A.; Culletta, G.; Almerico, A.M.; et al. Fighting Antibiotic Resistance: New Pyrimidine-Clubbed Benzimidazole Derivatives as Potential DHFR Inhibitors. Molecules 2023, 28, 501.
- Frei, A.; Verderosa, A.D.; Elliott, A.G.; Zuegg, J.; Blaskovich, M.A.T. Metals to Combat Antimicrobial Resistance. Nat. Rev. Chem. 2023, 7, 202–224.
- Raj, R.; Bhattu, M.; Verma, M.; Acevedo, R.; Duc, N.D.; Singh, J. Biogenic Silver Based Nanostructures: Synthesis, Mechanistic Approach and Biological Applications. Environ. Res. 2023, 231, 116045.
- Zafar, N.; Uzair, B.; Menaa, F.; Khan, B.A.; Niazi, M.B.K.; Alaryani, F.S.; Majrashi, K.A.; Sajjad, S. Moringa Concanensis-Mediated Synthesis and Characterizations of Ciprofloxacin Encapsulated into Ag/TiO2/Fe2O3/CS Nanocomposite: A Therapeutic Solution against Multidrug Resistant E. Coli Strains of Livestock Infectious Diseases. Pharmaceutics 2022, 14, 1719.
- Li, X.; Zheng, S.; Li, Y.; Ding, J.; Qin, W. Effectively Facilitating the Degradation of Chloramphenicol by the Synergism of Shewanella oneidensis MR-1 and the Metal-Organic Framework. J. Hazard. Mater. 2023, 454, 131545.
- Jiang, H.; Zhang, L.; Wang, X.; Gu, J.; Song, Z.; Wei, S.; Guo, H.; Xu, L.; Qian, X. Reductions in Abundances of Intracellular and Extracellular Antibiotic Resistance Genes by SiO2 Nanoparticles during Composting Driven by Mobile Genetic Elements. J. Environ. Manag. 2023, 341, 118071.
- Zhao, S.; Chang, Y.; Liu, J.; Sangeetha, T.; Feng, Y.; Liu, D.; Xu, C. Removal of Antibiotic Resistance Genes and Mobile Genetic Elements in a Three-Stage Pig Manure Management System: The Implications of Microbial Community Structure. J. Environ. Manag. 2022, 323, 116185.
- Yang, S.; Lu, C.; Qin, C.; Lu, C.; Pan, Z.; Zhao, L.; Bai, M.; Li, X.; Sun, Y.; Weng, L.; et al. Mitigation Effects and Microbial Mechanism of Two Ecological Earthworms on the Uptake of Chlortetracycline and Antibiotic Resistance Genes in Lettuce. Sci. Total Environ. 2023, 885, 163907.
- Mthiyane, Z.L.; Makhubela, N.; Nyoni, H.; Madikizela, L.M.; Maseko, B.R.; Ncube, S. Determination of Antibiotics during Treatment of Hospital Wastewater Using Automated Solid-Phase Extraction Followed by UHPLC-MS: Occurrence, Removal and Environmental Risks. Environ. Technol. 2023, 2023, 2209741.
- Gajdoš, S.; Zuzáková, J.; Pacholská, T.; Kužel, V.; Karpíšek, I.; Karmann, C.; Šturmová, R.; Bindzar, J.; Smrčková, Š.; Sýkorová, Z.; et al. Synergistic Removal of Pharmaceuticals and Antibiotic Resistance from Ultrafiltered WWTP Effluent: Free-Floating ARGs Exceptionally Susceptible to Degradation. J. Environ. Manag. 2023, 340, 117861.
- McNamara, E.; Bomkamp, C. Cultivated Meat as a Tool for Fighting Antimicrobial Resistance. Nat. Food 2022, 3, 791–794.
- Saied, A.A.; Chandran, D.; Chopra, H.; Dey, A.; Emran, T.B.; Dhama, K. Cultivated Meat Could Aid in Reducing Global Antimicrobial Resistance Burden—Producing Meat without Antibiotics as a Safer Food System for the Future. Int. J. Surg. 2023, 109, 189–190.
- Ghiga, I.; Sidorchuk, A.; Pitchforth, E.; Stålsby Lundborg, C.; Machowska, A. ‘If You Want to Go Far, Go Together’—Community-Based Behaviour Change Interventions to Improve Antibiotic Use: A Systematic Review of Quantitative and Qualitative Evidence. J. Antimicrob. Chemother. 2023, 78, 1344–1353.
- Allerton, F.; Jamieson, C.; Aggarwal, R.; Barker, A.; Work, M.; Cooper, D.; Ramsey, I. An Antibiotic Amnesty Can Be a One Health Tool to Tackle Antimicrobial Resistance. Nat. Med. 2023, 29, 1046–1047.
- McCracken, C.M.; Tucker, K.J.; Tallman, G.B.; Holmer, H.K.; Noble, B.N.; McGregor, J.C. General Perceptions and Knowledge of Antibiotic Resistance and Antibiotic Use Behavior: A Cross-Sectional Survey of US Adults. Antibiotics 2023, 12, 672.
- Rodríguez-González, A.; Zanin, M.; Menasalvas-Ruiz, E. Public Health and Epidemiology Informatics: Can Artificial Intelligence Help Future Global Challenges? An Overview of Antimicrobial Resistance and Impact of Climate Change in Disease Epidemiology. Yearb. Med. Inform. 2019, 28, 224–231.
- Kula, C.; Arga, K.Y. Systems Biomarkers, Artificial Intelligence, and One Health Vision Can Help Fight Antimicrobial Resistance. Omics J. Integr. Biol. 2023, 27, 191–192.
- Lau, H.J.; Lim, C.H.; Foo, S.C.; Tan, H.S. The Role of Artificial Intelligence in the Battle against Antimicrobial-Resistant Bacteria. Curr. Genet. 2021, 67, 421–429.
- Pineros, N.; Tenaillon, K.; Marin, J.; Berry, V.; Jaureguy, F.; Ghelfenstein-Ferreira, T.; Carbonnelle, E.; Lescat, M. Using Gamification to Improve Engagement and Learning Outcomes in Medical Microbiology: The Case Study of “BacteriaGame”. FEMS Microbiol. Lett. 2023, 370, fnad034.
- Mehrabi, M.R.; Soltani, M.; Chiani, M.; Raahemifar, K.; Farhangi, A. Nanomedicine: New Frontiers in Fighting Microbial Infections. Nanomaterials 2023, 13, 483.
- Yi, W.; Yan, D.; Wang, D.; Li, Y. Smart Drug Delivery Systems to Overcome Drug Resistance in Cancer Immunotherapy. Cancer Biol. Med. 2023, 20, 248–267.
- Yang, P.; Ren, J.; Yang, L. Nanoparticles in the New Era of Cardiovascular Therapeutics: Challenges and Opportunities. Int. J. Mol. Sci. 2023, 24, 5205.
- Maddiboyina, B.; Ramaiah, R.; Nakkala, R.K.; Roy, H. Perspectives on Cutting-Edge Nanoparticulate Drug Delivery Technologies Based on Lipids and Their Applications. Chem. Biol. Drug Des. 2023, 102, 377–394.
- Vairo, C.; Villar Vidal, M.; Maria Hernandez, R.; Igartua, M.; Villullas, S. Colistin- and Amikacin-Loaded Lipid-Based Drug Delivery Systems for Resistant Gram-Negative Lung and Wound Bacterial Infections. Int. J. Pharm. 2023, 635, 122739.
- Kiymaci, M.E.; Topal, G.R.; Esim, O.; Bacanli, M.; Ozkan, C.K.; Erdem, O.; Savaser, A.; Ozkan, Y. Evaluation of Bacterial Uptake, Antibacterial Efficacy against Escherichia coli, and Cytotoxic Effects of Moxifloxacin-Loaded Solid Lipid Nanoparticles. Arh. Hig. Rada Toksikol. 2022, 73, 260–269.
- Russo, S.; Torrisi, C.; Cardullo, N.; Muccilli, V.; La Mantia, A.; Castelli, F.; Acquaviva, R.; Sarpietro, M.G. Ethyl Protocatechuate Encapsulation in Solid Lipid Nanoparticles: Assessment of Pharmacotechnical Parameters and Preliminary In Vitro Evaluation for Colorectal Cancer Treatment. Pharmaceutics 2023, 15, 394.
- Magnifico, I.; Petronio Petronio, G.; Venditti, N.; Cutuli, M.A.; Pietrangelo, L.; Vergalito, F.; Mangano, K.; Zella, D.; Di Marco, R. Atopic Dermatitis as a Multifactorial Skin Disorder. Can the Analysis of Pathophysiological Targets Represent the Winning Therapeutic Strategy? Pharmaceuticals 2020, 13, 411.
- Maja, L.; Željko, K.; Mateja, P. Sustainable Technologies for Liposome Preparation. J. Supercrit. Fluids 2020, 165, 104984.
- Rasti, B.; Jinap, S.; Mozafari, M.R.; Abd-Manap, M.Y. Optimization on Preparation Condition of Polyunsaturated Fatty Acids Nanoliposome Prepared by Mozafari Method. J. Liposome Res. 2014, 24, 99–105.
- Zhang, H. Thin-Film Hydration Followed by Extrusion Method for Liposome Preparation. Methods Mol. Biol. 2017, 1522, 17–22.
- Nosova, A.S.; Koloskova, O.O.; Nikonova, A.A.; Simonova, V.A.; Smirnov, V.V.; Kudlay, D.; Khaitov, M.R. Diversity of PEGylation Methods of Liposomes and Their Influence on RNA Delivery. MedChemComm 2019, 10, 369–377.
- Suzuki, T.; Suzuki, Y.; Hihara, T.; Kubara, K.; Kondo, K.; Hyodo, K.; Yamazaki, K.; Ishida, T.; Ishihara, H. PEG Shedding-Rate-Dependent Blood Clearance of PEGylated Lipid Nanoparticles in Mice: Faster PEG Shedding Attenuates Anti-PEG IgM Production. Int. J. Pharm. 2020, 588, 119792.
- Kolašinac, R.; Kleusch, C.; Braun, T.; Merkel, R.; Csiszár, A. Deciphering the Functional Composition of Fusogenic Liposomes. Int. J. Mol. Sci. 2018, 19, 346.
- Kato, Y.; Ozawa, S.; Miyamoto, C.; Maehata, Y.; Suzuki, A.; Maeda, T.; Baba, Y. Acidic Extracellular Microenvironment and Cancer. Cancer Cell Int. 2013, 13, 89.
- Kromer, C.; Schwibbert, K.; Gadicherla, A.K.; Thiele, D.; Nirmalananthan-Budau, N.; Laux, P.; Resch-Genger, U.; Luch, A.; Tschiche, H.R. Monitoring and Imaging pH in Biofilms Utilizing a Fluorescent Polymeric Nanosensor. Sci. Rep. 2022, 12, 9823.
- Lee, R.J.; Low, P.S. Folate-Mediated Tumor Cell Targeting of Liposome-Entrapped Doxorubicin in Vitro. Biochim. Biophys. Acta BBA Biomembr. 1995, 1233, 134–144.
- Tang, B.; Peng, Y.; Yue, Q.; Pu, Y.; Li, R.; Zhao, Y.; Hai, L.; Guo, L.; Wu, Y. Design, Preparation and Evaluation of Different Branched Biotin Modified Liposomes for Targeting Breast Cancer. Eur. J. Med. Chem. 2020, 193, 112204.
- Ye, Z.; Gastfriend, B.D.; Umlauf, B.J.; Lynn, D.M.; Shusta, E.V. Antibody-Targeted Liposomes for Enhanced Targeting of the Blood-Brain Barrier. Pharm. Res. 2022, 39, 1523–1534.
- Kumar, V.; Kumar, R.; Jain, V.K.; Nagpal, S. Preparation and Characterization of Nanocurcumin Based Hybrid Virosomes as a Drug Delivery Vehicle with Enhanced Anticancerous Activity and Reduced Toxicity. Sci. Rep. 2021, 11, 368.
- Batrakova, E.V.; Kim, M.S. Using Exosomes, Naturally-Equipped Nanocarriers, for Drug Delivery. J. Control. Release 2015, 219, 396–405.
- Si, Q.; Wu, L.; Pang, D.; Jiang, P. Exosomes in Brain Diseases: Pathogenesis and Therapeutic Targets. MedComm 2023, 4, e287.
- Chen, J.; Li, P.; Zhang, T.; Xu, Z.; Huang, X.; Wang, R.; Du, L. Review on Strategies and Technologies for Exosome Isolation and Purification. Front. Bioeng. Biotechnol. 2021, 9, 811971.
- Vesiclepedia: Home: Extracellular Vesicles Database. Available online: http://microvesicles.org/ (accessed on 26 August 2023).
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514.
- Fagone, P.; Mangano, K.; Di Marco, R.; Reyes-Castillo, Z.; Muñoz-Valle, J.F.; Nicoletti, F. Altered Expression of TSPAN32 during B Cell Activation and Systemic Lupus Erythematosus. Genes 2021, 12, 931.
- Avgoulas, D.I.; Tasioulis, K.S.; Papi, R.M.; Pantazaki, A.A. Therapeutic and Diagnostic Potential of Exosomes as Drug Delivery Systems in Brain Cancer. Pharmaceutics 2023, 15, 1439.
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.-M.; Xu, Y.-M.; Huang, L.-F.; Wang, X.-Z. Exosomes: Novel Biomarkers for Clinical Diagnosis. Sci. World J. 2015, 2015, 657086.
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying Extracellular Vesicles Based Therapeutics in Clinical Trials—An ISEV Position Paper. J. Extracell. Vesicles 2015, 4, 30087.
- Lagacé, J.; Dubreuil, M.; Montplaisir, S. Liposome-Encapsulated Antibiotics: Preparation, Drug Release and Antimicrobial Activity against Pseudomonas aeruginosa. J. Microencapsul. 2008, 8, 53–61.
- Hsu, C.-Y.; Yang, S.-C.; Sung, C.T.; Weng, Y.-H.; Fang, J.-Y. Anti-MRSA Malleable Liposomes Carrying Chloramphenicol for Ameliorating Hair Follicle Targeting. Int. J. Nanomed. 2017, 12, 8227–8238.
- Bapolisi, A.M.; Nkanga, C.I.; Walker, R.B.; Krause, R.W.M. Simultaneous Liposomal Encapsulation of Antibiotics and Proteins: Co-Loading and Characterization of Rifampicin and Human Serum Albumin in Soy-Liposomes. J. Drug Deliv. Sci. Technol. 2020, 58, 101751.
- Halwani, M.; Mugabe, C.; Azghani, A.O.; Lafrenie, R.M.; Kumar, A.; Omri, A. Bactericidal Efficacy of Liposomal Aminoglycosides against Burkholderia cenocepacia. J. Antimicrob. Chemother. 2007, 60, 760–769.
- Hamblin, K.A.; Armstrong, S.J.; Barnes, K.B.; Davies, C.; Laws, T.; Blanchard, J.D.; Harding, S.V.; Atkins, H.S. Inhaled Liposomal Ciprofloxacin Protects against a Lethal Infection in a Murine Model of Pneumonic Plague. Front. Microbiol. 2017, 8, 91.
- d’Angelo, I.; Conte, C.; La Rotonda, M.I.; Miro, A.; Quaglia, F.; Ungaro, F. Improving the Efficacy of Inhaled Drugs in Cystic Fibrosis: Challenges and Emerging Drug Delivery Strategies. Adv. Drug Deliv. Rev. 2014, 75, 92–111.
- Vairo, C.; Villar Vidal, M.; Maria Hernandez, R.; Igartua, M.; Villullas, S. Colistin- and Amikacin-Loaded Lipid-Based Drug Delivery Systems for Resistant Gram-Negative Lung and Wound Bacterial Infections. Int. J. Pharm. 2023, 635, 122739.
- Bilton, D.; Pressler, T.; Fajac, I.; Clancy, J.P.; Sands, D.; Minic, P.; Cipolli, M.; Galeva, I.; Solé, A.; Quittner, A.L.; et al. Amikacin Liposome Inhalation Suspension for Chronic Pseudomonas aeruginosa Infection in Cystic Fibrosis. J. Cyst. Fibros. 2020, 19, 284–291.
- Pires, D.; Mandal, M.; Pinho, J.; Catalão, M.J.; Almeida, A.J.; Azevedo-Pereira, J.M.; Gaspar, M.M.; Anes, E. Liposomal Delivery of Saquinavir to Macrophages Overcomes Cathepsin Blockade by Mycobacterium tuberculosis and Helps Control the Phagosomal Replicative Niches. Int. J. Mol. Sci. 2023, 24, 1142.
- Khameneh, B.; Iranshahy, M.; Ghandadi, M.; Ghoochi Atashbeyk, D.; Fazly Bazzaz, B.S.; Iranshahi, M. Investigation of the Antibacterial Activity and Efflux Pump Inhibitory Effect of Co-Loaded Piperine and Gentamicin Nanoliposomes in Methicillin-Resistant Staphylococcus aureus. Drug Dev. Ind. Pharm. 2015, 41, 989–994.
- Hedges, J.F.; Snyder, D.T.; Robison, A.; Thompson, M.A.; Aspelin, K.; Plewa, J.; Baldridge, J.; Jutila, M.A. A TLR4 Agonist Liposome Formulation Effectively Stimulates Innate Immunity and Enhances Protection from Bacterial Infection. Innate Immun. 2023, 29, 45–57.
- Giannaccare, G.; Comis, S.; Jannuzzi, V.; Camposampiero, D.; Ponzin, D.; Cambria, S.; Santocono, M.; Pallozzi Lavorante, N.; Del Noce, C.; Scorcia, V.; et al. Effect of Liposomal-Lactoferrin-Based Eye Drops on the Conjunctival Microflora of Patients Undergoing Cataract Surgery. Ophthalmol. Ther. 2023, 12, 1315–1326.
- Karpuz, M.; Temel, A.; Ozgenc, E.; Tekintas, Y.; Erel-Akbaba, G.; Senyigit, Z.; Atlihan-Gundogdu, E. 99mTc-Labeled, Colistin Encapsulated, Theranostic Liposomes for Pseudomonas aeruginosa Infection. AAPS PharmSciTech 2023, 24, 77.
- Guo, R.; Liu, Y.; Li, K.; Tian, B.; Li, W.; Niu, S.; Hong, W. Direct Interactions between Cationic Liposomes and Bacterial Cells Ameliorate the Systemic Treatment of Invasive Multidrug-Resistant Staphylococcus aureus Infections. Nanomed. Nanotechnol. Biol. Med. 2021, 34, 102382.
- Patil, R.; Torris, A.; Bhat, S.; Patil, S. Mapping Fusogenicity of Ciprofloxacin-Loaded Liposomes with Bacterial Cells. AAPS PharmSciTech 2019, 20, 180.
- Nicolosi, D.; Scalia, M.; Nicolosi, V.M.; Pignatello, R. Encapsulation in Fusogenic Liposomes Broadens the Spectrum of Action of Vancomycin against Gram-Negative Bacteria. Int. J. Antimicrob. Agents 2010, 35, 553–558.
- Darvishi, M.; Farahani, S.; Haeri, A. Moxifloxacin-Loaded Lipidic Nanoparticles for Antimicrobial Efficacy. Curr. Pharm. Des. 2021, 27, 135–140.
- Longo, J.P.F.; Leal, S.C.; Simioni, A.R.; de Fátima Menezes Almeida-Santos, M.; Tedesco, A.C.; Azevedo, R.B. Photodynamic Therapy Disinfection of Carious Tissue Mediated by Aluminum-Chloride-Phthalocyanine Entrapped in Cationic Liposomes: An in Vitro and Clinical Study. Lasers Med. Sci. 2012, 27, 575–584.
- Ferro, S.; Ricchelli, F.; Monti, D.; Mancini, G.; Jori, G. Efficient Photoinactivation of Methicillin-Resistant Staphylococcus aureus by a Novel Porphyrin Incorporated into a Poly-Cationic Liposome. Int. J. Biochem. Cell Biol. 2007, 39, 1026–1034.
- Bartomeu Garcia, C.; Shi, D.; Webster, T.J. Tat-Functionalized Liposomes for the Treatment of Meningitis: An in Vitro Study. Int. J. Nanomed. 2017, 12, 3009–3021.
- Hu, Z.; Tang, Y.; Jiang, B.; Xu, Y.; Liu, S.; Huang, C. Functional Liposome Loaded Curcumin for the Treatment of Streptococcus mutans Biofilm. Front. Chem. 2023, 11, 1160521.
- Vanamala, K.; Bhise, K.; Sanchez, H.; Kebriaei, R.; Luong, D.; Sau, S.; Abdelhady, H.; Rybak, M.J.; Andes, D.; Iyer, A.K. Folate Functionalized Lipid Nanoparticles for Targeted Therapy of Methicillin-Resistant Staphylococcus aureus. Pharmaceutics 2021, 13, 1791.
- Rani, N.N.; Chen, X.Y.; Al-Zubaidi, Z.M.; Azhari, H.; Khaitir, T.M.N.; Ng, P.Y.; Buang, F.; Tan, G.C.; Wong, Y.P.; Said, M.M.; et al. Surface-Engineered Liposomes for Dual-Drug Delivery Targeting Strategy against Methicillin-Resistant Staphylococcus aureus (MRSA). Asian J. Pharm. Sci. 2022, 17, 102–119.
- Uhl, P.; Sauter, M.; Hertlein, T.; Witzigmann, D.; Laffleur, F.; Hofhaus, G.; Fidelj, V.; Tursch, A.; Özbek, S.; Hopke, E.; et al. Overcoming the Mucosal Barrier: Tetraether Lipid-Stabilized Liposomal Nanocarriers Decorated with Cell-Penetrating Peptides Enable Oral Delivery of Vancomycin. Adv. Ther. 2021, 4, 2000247.
- Singla, A.; Simbassa, S.B.; Chirra, B.; Gairola, A.; Southerland, M.R.; Shah, K.N.; Rose, R.E.; Chen, Q.; Basharat, A.; Baeza, J.; et al. Hetero-Multivalent Targeted Liposomal Drug Delivery to Treat Pseudomonas aeruginosa Infections. ACS Appl. Mater. Interfaces 2022, 14, 40724–40737.
- Wang, R.; Song, C.; Gao, A.; Liu, Q.; Guan, W.; Mei, J.; Ma, L.; Cui, D. Antibody-Conjugated Liposomes Loaded with Indocyanine Green for Oral Targeted Photoacoustic Imaging-Guided Sonodynamic Therapy of Helicobacter pylori Infection. Acta Biomater. 2022, 143, 418–427.
- Chen, R.; Ji, Y.; Li, T.; Zhao, B.; Guo, H.; Wang, Z.; Yao, H.; Zhang, Z.; Liu, C.; Du, M. Anti-Porphyromonas gingivalis Nanotherapy for Maintaining Bacterial Homeostasis in Periodontitis. Int. J. Antimicrob. Agents 2023, 61, 106801.
- Chen, X.; Yang, H.; Li, C.; Hu, W.; Cui, H.; Lin, L. Enhancing the Targeting Performance and Prolonging the Antibacterial Effects of Clove Essential Oil Liposomes to Campylobacter Jejuni by Antibody Modification. Food Res. Int. 2023, 167, 112736.
- Krivić, H.; Himbert, S.; Sun, R.; Feigis, M.; Rheinstädter, M.C. Erythro-PmBs: A Selective Polymyxin B Delivery System Using Antibody-Conjugated Hybrid Erythrocyte Liposomes. ACS Infect. Dis. 2022, 8, 2059–2072.
- Moreira, L.; Guimarães, N.M.; Pereira, S.; Santos, R.S.; Loureiro, J.A.; Ferreira, R.M.; Figueiredo, C.; Pereira, M.C.; Azevedo, N.F. Engineered Liposomes to Deliver Nucleic Acid Mimics in Escherichia coli. J. Control. Release 2023, 355, 489–500.
- Shi, Y.; Feng, X.; Lin, L.; Wang, J.; Chi, J.; Wu, B.; Zhou, G.; Yu, F.; Xu, Q.; Liu, D.; et al. Virus-Inspired Surface-Nanoengineered Antimicrobial Liposome: A Potential System to Simultaneously Achieve High Activity and Selectivity. Bioact. Mater. 2021, 6, 3207–3217.
- Noguchi, K.; Obuki, M.; Sumi, H.; Klußmann, M.; Morimoto, K.; Nakai, S.; Hashimoto, T.; Fujiwara, D.; Fujii, I.; Yuba, E.; et al. Macropinocytosis-Inducible Extracellular Vesicles Modified with Antimicrobial Protein CAP18-Derived Cell-Penetrating Peptides for Efficient Intracellular Delivery. Mol. Pharm. 2021, 18, 3290–3301.
- Celebi, D.; Celebi, O.; Baser, S.; Taghizadehghalehjoughi, A. Evaluation of Antimicrobial and Antibiofilm Efficacy of Bee Venom and Exosome Against Escherichia coli K99 Strain. Kafkas Üniv. Vet. Fak. Derg. 2023, 29, 239–246.
- Cheng, Y.; Schorey, J.S. Exosomes Carrying Mycobacterial Antigens Can Protect Mice against Mycobacterium tuberculosis Infection. Eur. J. Immunol. 2013, 43, 3279–3290.
- Hazrati, A.; Soudi, S.; Hashemi, S.M. Wharton’s Jelly Mesenchymal Stem Cells-Derived Exosomes and Imipenem in Combination Reduce Apoptosis and Inflammatory Responses in E. coli-Infected HepG2 Cells. Iran. J. Allergy Asthma Immunol. 2022, 21, 273.
- Yang, X.; Xie, B.; Peng, H.; Shi, G.; Sreenivas, B.; Guo, J.; Wang, C.; He, Y. Eradicating Intracellular MRSA via Targeted Delivery of Lysostaphin and Vancomycin with Mannose-Modified Exosomes. J. Control. Release 2021, 329, 454–467.
- Yang, X.; Shi, G.; Guo, J.; Wang, C.; He, Y. Exosome-Encapsulated Antibiotic against Intracellular Infections of Methicillin-Resistant Staphylococcus aureus. Int. J. Nanomed. 2018, 13, 8095–8104.