Carnivorous plants attract animals, trap and kill them, and absorb nutrients from the digested bodies. This unusual (for autotrophs) type of nutrient acquisition evolved through the conversion of photosynthetically active leaves into specialised organs commonly called traps. The genus Nepenthes (pitcher plants) consists of approximately 169 species belonging to the group of carnivorous plants. Pitcher plants are characterised by specialised passive traps filled with a digestive fluid. The digestion that occurs inside the traps of carnivorous plants depends on the activities of many enzymes. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) also participate in the digestive process.
- carnivorous plants
- Nepenthes
- reactive nitrogen species
- reactive oxygen species
1. Carnivorous Plants
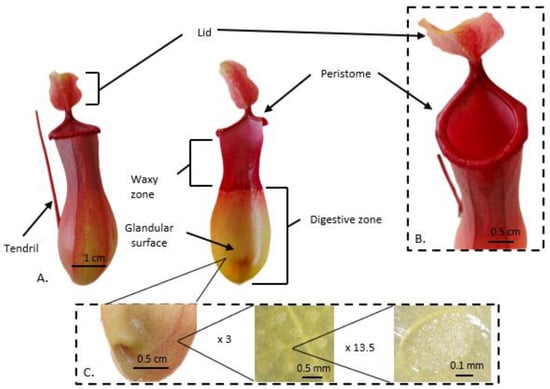
2. Carnivorous Pitcher Plants
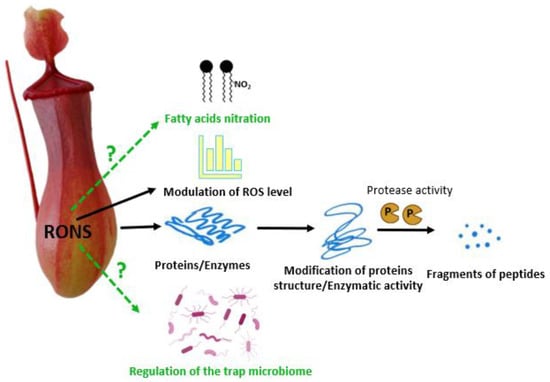
3. The Role of Reactive Oxygen Species in External Digestion by Carnivorous Plants
This entry is adapted from the peer-reviewed paper 10.3390/biology12101356
References
- Melzig, M.F.; Pertz, H.H.; Krenn, L. Anti-Inflammatory and Spasmolytic Activity of Extracts from Droserae herba. Phytomedicine 2001, 8, 225–229.
- Mithöfer, A. Carnivorous Pitcher Plants: Insights in an Old Topic. Phytochemistry 2011, 72, 1678–1682.
- Morohoshi, T.; Oikawa, M.; Sato, S.; Kikuchi, N.; Kato, N.; Ikeda, T. Isolation and Characterization of Novel Lipases from a Metagenomic Library of the Microbial Community in the Pitcher Fluid of the Carnivorous Plant Nepenthes hybrida. J. Biosci. Bioeng. 2011, 112, 315–320.
- Darwin, C. Insectivorous Plants; D. Appleton and Company: New York, NY, USA, 1875.
- Ellison, A.M.; Gotelli, N.J. Energetics and the Evolution of Carnivorous Plants—Darwin’s ‘Most Wonderful Plants in the World’. J. Exp. Bot. 2009, 60, 19–42.
- Armstrong, W. Aeration in Higher Plants. Adv. Bot. Res. 1979, 7, 225–332.
- Jürgens, A.; El-Sayed, A.M.; Suckling, D.M. Do Carnivorous Plants Use Volatiles for Attracting Prey Insects? Funct. Ecol. 2009, 23, 875–887.
- Adlassnig, W.; Steinhauser, G.; Peroutka, M.; Musilek, A.; Sterba, J.H.; Lichtscheidl, I.K.; Bichler, M. Expanding the Menu for Carnivorous Plants: Uptake of Potassium, Iron and Manganese by Carnivorous Pitcher Plants. Appl. Radiat. Isot. 2009, 67, 2117–2122.
- Król, E.; Płachno, B.J.; Adamec, L.; Stolarz, M.; Dziubińska, H.; Trębacz, K. Quite a Few Reasons for Calling Carnivores ‘the Most Wonderful Plants in the World’. Ann. Bot. 2012, 109, 47–64.
- Płachno, B.J.; Adamec, L.; Huet, H. Mineral Nutrient Uptake from Prey and Glandular Phosphatase Activity as a Dual Test of Carnivory in Semi-Desert Plants with Glandular Leaves Suspected of Carnivory. Ann. Bot. 2009, 104, 649–654.
- Cross, A.T.; Krueger, T.A.; Gonella, P.M.; Robinson, A.S.; Fleischmann, A.S. Conservation of Carnivorous Plants in the Age of Extinction. Glob. Ecol. Conserv. 2020, 24, e01272.
- Millett, J.; Svensson, B.M.; Newton, J.; Rydin, H. Reliance on Prey-Derived Nitrogen by the Carnivorous Plant Drosera rotundifolia Decreases with Increasing Nitrogen Deposition. New Phytol. 2012, 195, 182–188.
- Adlassnig, W.; Peroutka, M.; Lambers, H.; Lichtscheidl, I.K. The Roots of Carnivorous Plants. Plant Soil. 2005, 274, 127–140.
- Lloyd, F.E. The Carnivorous Plants; Chronica Botanica Company: Waltham, MA, USA, 1942.
- Albert, V.A.; Williams, S.E.; Chase, M.W. Carnivorous Plants: Phylogeny and Structural Evolution. Science (1979) 1992, 257, 1491–1495.
- Clarke, C.; Moran, J.A.; Lee, C.C. Nepenthes baramensis (Nepenthaceae)—A New Species from North-Western Borneo. Blumea Biodivers. Evol. Biogeogr. Plants 2011, 56, 229–233.
- Meimberg, H.; Wistuba, A.; Dittrich, P.; Heubl, G. Molecular Phylogeny of Nepenthaceae Based on Cladistic Analysis of Plastid TrnK Intron Sequence Data. Plant Biol. 2001, 3, 164–175.
- Meimberg, H.; Heubl, G. Introduction of a Nuclear Marker for Phylogenetic Analysis of Nepenthaceae. Plant Biol. 2006, 8, 831–840.
- Bauer, U.; Grafe, T.U.; Federle, W. Evidence for Alternative Trapping Strategies in Two Forms of the Pitcher Plant. Nepenthes rafflesiana. J. Exp. Bot. 2011, 62, 3683–3692.
- Clarke, C. Nepenthes of Borneo; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 1997; ISBN 9789838120159.
- Dančák, M.; Majeský, Ľ.; Čermák, V.; Golos, M.R.; Płachno, B.J.; Tjiasmanto, W. First Record of Functional Underground Traps in a Pitcher Plant: Nepenthes pudica (Nepenthaceae), a New Species from North Kalimantan, Borneo. PhytoKeys 2022, 201, 77–97.
- Adlassnig, W.; Peroutka, M.; Lendl, T. Traps of Carnivorous Pitcher Plants as a Habitat: Composition of the Fluid, Biodiversity and Mutualistic Activities. Ann. Bot. 2011, 107, 181–194.
- Gaume, L.; Di Giusto, B. Adaptive Significance and Ontogenetic Variability of the Waxy Zone in Nepenthes rafflesiana. Ann. Bot. 2009, 104, 1281–1291.
- Gaume, L.; Gorb, S.; Rowe, N. Function of Epidermal Surfaces in the Trapping Efficiency of Nepenthes alata Pitchers. New Phytol. 2002, 156, 479–489.
- Gaume, L.; Perret, P.; Gorb, E.; Gorb, S.; Labat, J.J.; Rowe, N. How Do Plant Waxes Cause Flies to Slide? Experimental Tests of Wax-Based Trapping Mechanisms in Three Pitfall Carnivorous Plants. Arthropod Struct. Dev. 2004, 33, 103–111.
- Wang, L.; Tao, D.; Dong, S.; Li, S.; Tian, Y. Contributions of Lunate Cells and Wax Crystals to the Surface Anisotropy of Nepenthes Slippery Zone. R. Soc. Open Sci. 2018, 5, 180766.
- Gorb, E.; Haas, K.; Henrich, A.; Enders, S.; Barbakadze, N.; Gorb, S. Composite Structure of the Crystalline Epicuticular Wax Layer of the Slippery Zone in the Pitchers of the Carnivorous Plant Nepenthes alata and Its Effect on Insect Attachment. J. Exp. Biol. 2005, 208, 4651–4662.
- Moran, J.A.; Clarke, C.M.; Hawkins, B.J. From Carnivore to Detritivore? Isotopic Evidence for Leaf Litter Utilization by the Tropical Pitcher Plant Nepenthes ampullaria. Int. J. Plant Sci. 2003, 164, 635–639.
- Bohn, H.F.; Federle, W. Insect Aquaplaning: Nepenthes Pitcher Plants Capture Prey with the Peristome, a Fully Wettable Water-Lubricated Anisotropic Surface. Proc. Natl. Acad. Sci. USA 2004, 101, 14138–14143.
- Moran, J.A.; Clarke, C.M. The Carnivorous Syndrome in Nepenthes Pitcher Plants. Plant Signal Behav. 2010, 5, 644–648.
- Kang, V.; Isermann, H.; Sharma, S.; Wilson, D.I.; Federle, W. How a Sticky Fluid Facilitates Prey Retention in a Carnivorous Pitcher Plant (Nepenthes rafflesiana). Acta Biomater. 2021, 128, 357–369.
- Hatano, N.; Hamada, T. Proteomic Analysis of Secreted Protein Induced by a Component of Prey in Pitcher Fluid of the Carnivorous Plant Nepenthes alata. J. Proteom. 2012, 75, 4844–4852.
- Eilenberg, H.; Pnini-Cohen, S.; Rahamim, Y.; Sionov, E.; Segal, E.; Carmeli, S.; Zilberstein, A. Induced Production of Antifungal Naphthoquinones in the Pitchers of the Carnivorous Plant Nepenthes khasiana. J. Exp. Bot. 2010, 61, 911–922.
- Raj, G.; Kurup, R.; Hussain, A.A.; Baby, S. Distribution of Naphthoquinones, Plumbagin, Droserone, and 5-O-Methyl Droserone in Chitin-Induced and Uninduced Nepenthes khasiana: Molecular Events in Prey Capture. J. Exp. Bot. 2011, 62, 5429–5436.
- Aung, H.H.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Ahmed, A.A.; Pare, P.W.; Mabry, T.J. Phenolic Constituents from the Leaves of the Carnivorous Plant Nepenthes gracilis. Fitoterapia 2002, 73, 445–447.
- Clarke, C.M.; Bauer, U.; Lee, C.C.; Tuen, A.A.; Rembold, K.; Moran, J.A. Tree Shrew Lavatories: A Novel Nitrogen Sequestration Strategy in a Tropical Pitcher Plant. Biol. Lett. 2009, 5, 632–635.
- Grafe, T.U.; Schöner, C.R.; Kerth, G.; Junaidi, A.; Schöner, M.G. A Novel Resource-Service Mutualism between Bats and Pitcher Plants. Biol. Lett. 2011, 7, 436–439.
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19.
- Ciacka, K.; Krasuska, U.; Staszek, P.; Wal, A.; Zak, J.; Gniazdowska, A. Effect of Nitrogen Reactive Compounds on Aging in Seed. Front. Plant Sci. 2020, 11, 1011.
- Horn, A.; Jaiswal, J.K. Cellular Mechanisms and Signals That Coordinate Plasma Membrane Repair. Cell. Mol. Life Sci. 2018, 75, 3751–3770.
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Production Sites of Reactive Oxygen Species (ROS) in Organelles from Plant Cells. In Reactive Oxygen Species and Oxidative Damage in Plants under Stress; Corpas, F.J., Gupta, D.K., Palma, J.M., Eds.; Springer: Cham, Switzerland, 2015; pp. 1–22.
- Thiel, J.; Rolletschek, H.; Friedel, S.; Lunn, J.E.; Nguyen, T.H.; Feil, R.; Tschiersch, H.; Müller, M.; Borisjuk, L. Seed-Specific Elevation of Non-Symbiotic Hemoglobin AtHb1: Beneficial Effects and Underlying Molecular Networks in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 48.
- Møller, I.M.; Rogowska-Wrzesinska, A.; Rao, R.S.P. Protein Carbonylation and Metal-Catalyzed Protein Oxidation in a Cellular Perspective. J. Proteomics 2011, 74, 2228–2242.
- Kumar, J.S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed Birth to Death: Dual Functions of Reactive Oxygen Species in Seed Physiology. Ann. Bot. 2015, 116, 663–668.
- Tamura, M.; Mutoh, M.; Fujii, G.; Matsui, H. Involvement of Mitochondrial Reactive Oxygen Species in Gastric Carcinogenesis. J. Gastrointest. Dig. Syst. 2013, 3, 150.
- Toyokuni, S.; Mori, T.; Hiai, H.; Dizdaroglu, M. Treatment of Wistar Rats with a Renal Carcinogen, Ferric Nitrilotriacetate, Causes DNA-protein Cross-linking between Thymine and Tyrosine in Their Renal Chromatin. Int. J. Cancer 1995, 62, 309–313.
- Kasai, H.; Nishimura, S. Hydroxylation of Deoxyguanosine at the C-8 Position by Ascorbic Acid and Other Reducing Agents. Nucleic Acids Res. 1984, 12, 2137–2145.
- Ciacka, K.; Tymiński, M.; Gniazdowska, A.; Krasuska, U. Carbonylation of Proteins—An Element of Plant Ageing. Planta 2020, 252, 12.
- Morscher, R.J.; Aminzadeh-Gohari, S.; Feichtinger, R.G.; Mayr, J.A.; Lang, R.; Neureiter, D.; Sperl, W.; Kofler, B. Inhibition of Neuroblastoma Tumor Growth by Ketogenic Diet and/or Calorie Restriction in a CD1-Nu Mouse Model. PLoS ONE 2015, 10, e0129802.
- Kranner, I.; Birtić, S.; Anderson, K.M.; Pritchard, H.W. Glutathione Half-Cell Reduction Potential: A Universal Stress Marker and Modulator of Programmed Cell Death? Free Radic. Biol. Med. 2006, 40, 2155–2165.
- Landriscina, M.; Maddalena, F.; Laudiero, G.; Esposito, F. Adaptation to Oxidative Stress, Chemoresistance, and Cell Survival. Antioxid. Redox Signal. 2009, 11, 2701–2716.
- Babior, B.M. Oxidants from Phagocytes: Agents of Defense and Destruction. Blood 1984, 64, 959–966.
- Takaki, A.; Kawano, S.; Uchida, D.; Takahara, M.; Hiraoka, S.; Okada, H. Paradoxical Roles of Oxidative Stress Response in the Digestive System before and after Carcinogenesis. Cancers 2019, 11, 213.
- Wetscher, G.J.; Hinder, P.R.; Bagchi, D.; Perdikis, G.; Redmond, E.J.; Glaser, K.; Adrian, T.E.; Hinder, R.A. Free Radical Scavengers Prevent Reflux Esophagitis in Rats. Dig. Dis. Sci. 1995, 40, 1292–1296.
- Meining, A.; Classen, M. The Role of Diet and Lifestyle Measures in the Pathogenesis and Treatment of Gastroesophageal Reflux Disease. Am. J. Gastroenterol. 2000, 95, 2692–2697.
- Van Hecke, T.; Van Camp, J.; De Smet, S. Oxidation during Digestion of Meat: Interactions with the Diet and Helicobacter Pylori Gastritis, and Implications on Human Health. Compr. Rev. Food Sci. Food Saf. 2017, 16, 214–233.
- Davies, K.J. Protein Damage and Degradation by Oxygen Radicals. I. General. Aspects. J. Biol. Chem. 1987, 262, 9895–9901.
- Davies, K.J.A.; Lin, S.W. Degradation of Oxidatively Denatured Proteins in Escherichia Coli. Free Radic. Biol. Med. 1988, 5, 215–223.
- An, C.I.; Fukusaki, E.I.; Kobayashi, A. Aspartic Proteinases Are Expressed in Pitchers of the Carnivorous Plant Nepenthes alata Blanco. Planta 2002, 214, 661–667.
- Basset, G.; Raymond, P.; Malek, L.; Brouquisse, R. Changes in the Expression and the Enzymic Properties of the 20S Proteasome in Sugar-Starved Maize Roots. Evidence for an in Vivo Oxidation of the Proteasome. Plant Physiol. 2002, 128, 1149–1162.
- Krasuska, U.; Ciacka, K.; Dębska, K.; Bogatek, R.; Gniazdowska, A. Dormancy Alleviation by NO or HCN Leading to Decline of Protein Carbonylation Levels in Apple (Malus domestica Borkh.). Embryos. J. Plant Physiol. 2014, 171, 1132–1141.
- Chia, T.F.; Aung, H.H.; Osipov, A.N.; Goh, N.K.; Chia, L.S. Carnivorous Pitcher Plant Uses Free Radicals in the Digestion of Prey. Redox Report 2004, 9, 255–261.
- Wal, A.; Staszek, P.; Pakula, B.; Paradowska, M.; Krasuska, U. ROS and RNS Alterations in the Digestive Fluid of Nepenthes × Ventrata Trap at Different Developmental Stages. Plants 2022, 11, 3304.