Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Infectious Diseases
Staphylococcus aureus (S. aureus) is a common pathogen involved in community- and hospital-acquired infections. Its biofilm formation ability predisposes it to device-related infections. Methicillin-resistant S. aureus (MRSA) strains are associated with more serious infections and higher mortality rates and are more complex in terms of antibiotic resistance.
- MRSA
- MSSA
- biofilms
- PIA (polysaccharide intercellular adhesin)
1. Differences in Biofilm Formation by MRSA and MSSA
PIA (Polysaccharide intercellular adhesin), also called PNAG (poly-N-acetylglucosamine), is a glycan with repeats of β-1,6-linked 2-acetamido-2-deoxy-D-glucopyranosyl residues, involved in intercellular adhesion, aggregation, and attachment to abiotic surfaces for S. aureus. PIA synthesis depends on the ica(intercellular adhesin)-ADBC operon, whose expression products are IcaA, IcaD, IcaB, and IcaC [33]. IcaA is a transmembrane N-acetyl-glucosaminyltransferase that, together with IcaD, synthesizes N-acetyl-glucosamine oligomers of more than 20 residues. Then, IcaC translocates these polysaccharides out of the cell surface. Finally, IcaB deacetylates the poly-N-acetylglucosamine molecules, which is important for the adherence of these polymers to the outer cell surface and biofilm formation (Figure 1) [33]. The role of PIA and the ica operon in biofilm formation was first described in Staphylococcus epidermidis (S. epidermidis) [34]. Later, Crampton et al. showed for the first time the presence of the ica operon in all the strains of S. aureus analyzed. Also, the mutation of the ica locus in the S. aureus strain ATCC 35556 resulted in reduced biofilm formation [35]. Since then, PIA has been considered the main extracellular matrix component involved in biofilm formation by S. aureus. However, it is worth noting that not all the strains analyzed by Crampton were able to form biofilm in vitro [35]. Likewise, it was tested the ability to form biofilm by 128 S. aureus isolates, and only 57.1% showed a biofilm-positive phenotype, even though all of them were icaA-positive [36]. We now know that icaADBC genes are present in most of the S. aureus clinical isolates, and that the expression of those genes is tightly regulated in vitro; for example, CO2 levels, anaerobicity, glucose, and osmotic stress have been shown to influence ica operon expression and biofilm formation [19,37]. This may explain, at least in part, why not all the S. aureus isolates containing those genes can form biofilms.
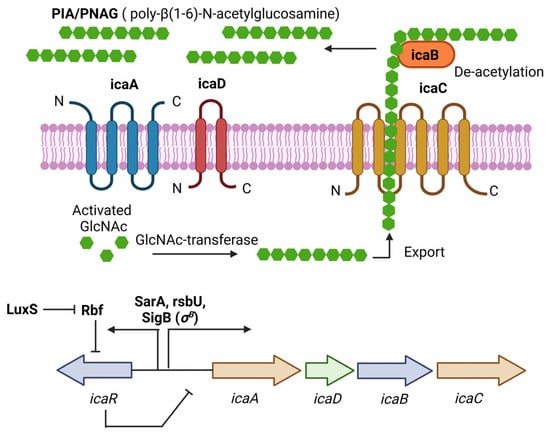
Figure 1. PIA (polysaccharide intercellular adhesin) is produced by the icaADBC operon in S. aureus. PIA-dependent biofilm formation has been described mainly for MSSA isolates under osmotic stress conditions. The ica operon is regulated by SarA and SigB. IcaR is part of the operon and acts as a negative regulator of PIA production.
Fitzpatrick et al. showed differences in biofilm formation by MRSA and MSSA clinical isolates using BHI medium alone or supplemented with 1% glucose or 4% NaCl. A higher percentage of MSSA isolates were able to form a biofilm in a NaCl medium in comparison with MRSA, while the proportion was similar in BHI glucose. It was suggested a possible association between methicillin susceptibility and biofilm formation depending on the specific environmental conditions [37]. It is worth mentioning that the association of biofilm production, methicillin susceptibility, and mecA expression was first analyzed in S. epidermidis [38,39]. Also, the transcription of the ica operon and biofilm formation induced by NaCl was first reported in S. epidermidis [40,41]. It is now well accepted that the ica operon is expressed under osmotic stress conditions, and this correlates with biofilm formation in MSSA strains [42]. In case of MRSA, it was shown an increase in biofilm formation by four ica-positive MRSA strains in a BHI medium supplemented with glucose but not in a BHI or BHI-NaCl medium [37]. Also, only in one MRSA strain was the expression of icaA induced in BHI-NaCl, but the deletion of the ica operon in this strain did not affect the biofilm formation in BHI-glucose. The expression of ica operon at the transcriptional level was not dependent on the presence of NaCl for the others three MRSA strains. The authors concluded that the ability to form biofilm only in BHI-glucose by these MRSA strains was independent of the ica-ADBC operon [37].
Later, the ability of MRSA and MSSA strains isolated from device-related infections to form biofilms was tested. Again, it was found that biofilm formation in BHI-NaCl was more frequent in MSSA strains, in comparison with MRSA, which preferentially formed biofilms in a medium containing glucose. For most of the MSSA strains tested, as expected, the addition of NaCl to the BHI medium was associated with the expression of icaA, PIA production, and biofilm formation. Even though the icaA expression was also increased in most MRSA strains in the presence of NaCl, PIA production was not observed [19], suggesting post-transcriptional regulation of PIA synthesis. Furthermore, deletion of the ica operon in the MSSA isolates affected their ability to form biofilms in BHI NaCl, while deletion of the ica operon in the MRSA isolates did not affect their ability to form biofilms in BHI glucose. Interestingly, glucose-induced MRSA biofilms were susceptible to treatment with proteinase K, while NaCl-induced MSSA biofilms were susceptible to sodium periodate, a reagent used to destabilize β-1,6-linked polysaccharides [19]. Thus, the chemical composition of the biofilms generated by both types of strains under those conditions seems to be different.
2. Biofilm Formation Independent of the icaADBC Operon
In addition to the aforementioned cases of biofilm formation independent of the ica operon in MRSA strains in a medium containing glucose, one of the first reports of an ica-independent biofilm formation in S. aureus was from a clinical isolate named UAMS-1 (MSSA). It was shown that a mutation in the ica locus had little impact on its biofilm formation capacity in vitro and in vivo [43]. Also, Lim et al. identified using mutagenesis analysis a gene called rbf involved in biofilm formation. The sequence suggested that it was a transcriptional regulator, and it did not affect the expression of the ica operon. Thus, it was considered an ica-independent mechanism [44]. Upstream of the ica operon is the icaR gene; Conlon et al. showed that icaR is a repressor of PIA production and biofilm formation by inhibiting the expression of the ica operon in S. epidermidis [40]. It was later shown that Rbf promotes biofilm formation by repressing the transcription of icaR in S. aureus [45].
Also found using mutagenesis analysis was a gene called bap (biofilm-associated protein) involved in attachment to inert surfaces, intercellular adhesion, and biofilm formation. Bap was described as a surface protein consisting of 2276 amino acids. However, it was only found in a small fraction of the S. aureus isolates from bovine mastitis lesions (5%), while it was absent in the clinical S. aureus isolates analyzed from humans [46]. In the S. aureus strain SA113, whose biofilm formation ability has been associated with the ica operon, complementation with the bap gene in the mutant strain SA113 Δica resulted in biofilm formation, suggesting the role of bap in biofilm formation independent of the ica operon [46]. In fact, bap is a surface protein containing the LPXTG motif. Since biofilms from MRSA strains in BHI-glucose were susceptible to proteinase K treatment, it is reasonable to believe that surface proteins such as bap must be responsible for the ica-independent biofilm formation in these strains. In this regard, a genomic analysis from MRSA strains revealed the presence of several LPXTG surface proteins presumed to be involved in the adherence to host tissues such as clumping factor A and B (Clf A/B), fibronectin binding protein A and B (Fnb A/B), collagen adhesin (Cna), protein A (Spa), methicillin resistance surface protein (Pls), SdrC/D/E, and several cell wall surface anchor proteins (Sas A/B/C/D/E/F/G/H/I/J/K). There were also found other non-LPXTG surface proteins such as autolysin (Atl), elastin binding protein (Ebp), and the fibrinogen binding proteins Fib and Efb [47].
Likewise, a mutation of sortase A (SrtA), an enzyme that catalyzes the cell wall anchoring of LPXTG proteins, in an S. aureus strain with an ica-independent biofilm phenotype was deficient in biofilm formation [48,49]. It was also shown using mass spectrometry that SdrD and protein A (Spa) were key components of the cell wall of an S. aureus strain able to form ica-independent biofilms, and deletion of the spa gene resulted in reduced biofilm formation [48]. SasC and SasG were also shown to promote biofilm formation in S. aureus [50,51]. Interestingly, it was found that MRSA strains in a BHI medium with glucose lowered the pH to mildly acidic conditions, and this was associated with the expression of fnbA and fnbB, and biofilm formation [49]. Mutations of fnbA and fnbB in several MRSA strains reduced biofilm formation. However, these mutations did not affect the biofilm formation in MSSA strains with a PIA-dependent biofilm phenotype. It was proposed that the effect on biofilm formation of fnbA and fnbB was due to intercellular aggregation rather than initial attachment [49]. Furthermore, carriage of both the fnbA and fnbB genes correlated with higher biofilm formation in MRSA strains in comparison with those strains carrying only one gene. It is worth mentioning that the proportion of bacteria expressing both genes was even higher in the MSSA clinical isolates, 69% (68/99) for MSSA, and 33% (40/118) for MRSA [52]. Thus, LPXTG proteins seem to be important in the proteinaceous biofilms formed by MRSA strains.
Also described was extracellular DNA (eDNA) as an important component of the S. aureus biofilms, as addition of DNase I to the medium or to the preformed biofilms resulted in their inhibition or destabilization, respectively [53,54]. CidA, a hydrolase involved in cell lysis, was shown to be important for DNA release and biofilm regulation in S. aureus [54]. Later, the major autolysin of S. aureus called Atl was found to mediate cell lysis and eDNA release, contributing to biofilm formation [55]. It was also shown that Atl was important for attachment and biofilm formation in MRSA strains with a FnbA/B biofilm phenotype, but not for MSSA strains with a PIA-dependent biofilm phenotype [56]. Furthermore, mutation of the protease ClpP in the S. aureus Newman strain resulted in an increase in biofilm formation, and it was a PIA-independent but protein- and eDNA-dependent biofilm [57]. Transcription of the agrA and agrC genes was reduced in the clpP-mutant strain, and it was associated with biofilm formation and decreased levels of protease activity. Interestingly, an increase in the expression of the hydrolase sle1 was found in the clpP-mutant strain, together with an increase in cell lysis and eDNA release [57]; however, it was not clear how the Agr system was affected in the clpP-mutant strain. Altogether, these results indicate the relevance of hydrolases in eDNA release and their contribution to biofilm formation by S. aureus. It is now well accepted that the biofilm composition in MRSA strains depends more on surface proteins and eDNA in comparison with the ica-dependent biofilm phenotype in MSSA strains [42]. Most of the studies have used a medium with NaCl for MSSA strains and a medium with glucose for MRSA strains, favoring the mentioned biofilm phenotypes. However, it is worth noting that a high proportion of MSSA isolates can also form biofilms in mediums containing glucose [19,20]. Thus, it is reasonable that depending on the environment, MSSA strains could form biofilms with a PIA-independent phenotype as well. Indeed, the first experiments analyzing the content of eDNA in the biofilms of S. aureus used UAMS-1 and SH1000, two MSSA strains [53,54,55].
Interestingly, a MRSA clinical isolate capable of switching from a PIA-dependent to a PIA-independent biofilm was found, depending on the medium composition. For example, in a TSB medium with glucose, it formed a PIA-independent biofilm, but when NaCl was added to the medium, detection of PIA and biofilm formation was observed. Deletion of ica in this S. aureus strain did not affect its capacity to form a biofilm in a medium with glucose, but it completely lost the capacity of biofilm formation in a medium with NaCl [58]. Sortase A (srtA) mutation revealed that LPXTG proteins were important for biofilm formation in a medium with glucose while they did not affect the generation of a PIA-dependent biofilm in a medium with NaCl. Genomic analysis revealed a unique repertoire of 20 LPXTG proteins in this strain, and deletion of fnbA and fnbB was associated with a complete loss of the PIA-independent biofilm. Also, fnbB was induced at the transcriptional level in the medium with glucose in comparison with the medium containing NaCl, but the icaC mRNA levels were unaffected in both types of media, suggesting again that there must exist post-transcriptional regulatory mechanisms in PIA expression [58].
3. Main Regulatory Systems in Biofilm Formation by S. aureus
The accessory gene regulator (agr) operon consists of the agrBDCA genes and is part of a quorum-sensing regulatory system in S. aureus. The expression of many virulence factors in S. aureus is controlled by the Agr system, which consists of two transcripts whose expression are controlled by promoters P2 and P3. The first transcript is an operon that encodes the four agrBDCA genes, and the second is RNAIII, the true effector molecule that regulates the expression of all the genes regulated by the Agr system. The gene encoding the δ-toxin (hld) is in the effector RNAIII regulatory molecule. This system is activated during the transition from the exponential growth phase to the stationary growth phase [59]. The Agr system responds to the extracellular levels of AIP (autoinducing peptide), an eight-residue peptide, with the last five residues forming a thiolactone cyclic ring. AIPs are formed via the action of AgrB, a membrane-bound peptidase that proteolytically processes AgrD, the peptide precursor of AIP. AgrC is a membrane-bound histidine kinase sensor of AIPs that is autophosphorylated, and this signal passes to the response regulator AgrA, which in turn binds to the promoters P2 and P3 to start their transcription (Figure 2) [60].
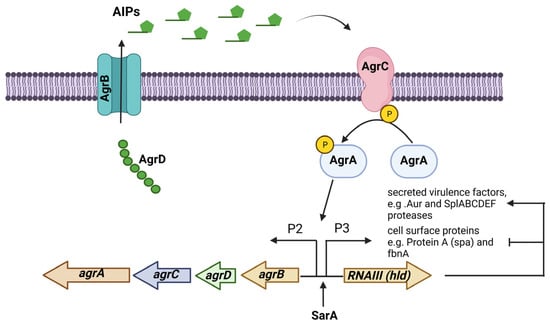
Figure 2. The Agr (accessory gene regulator) system is an operon that regulates the expression of several virulence factors in S. aureus. The agrBDCA operon is activated by SarA. In some S. aureus strains, the Agr system regulates PIA-independent biofilm formation by negatively regulating the expression of surface proteins while positively regulating the expression of extracellular proteases.
Analyzing the agr phenotype in terms of δ-toxin production (the product of the RNAIII transcript in the agr operon) and the biofilm formation capacity, it was demonstrated that 78% of the agr-negative S. aureus strains formed a biofilm in comparison with only 6% of the agr-positive strains [59]. It was also shown that agr mutation in S. aureus strains enhanced biofilm formation in comparison with the wild-type strains. Interestingly, the expression of PIA was not regulated by the Agr system [59]. It has been suggested that agr is repressed in a medium containing glucose and it correlates with biofilm formation [42]. Indeed, it was shown that expression of RNAIII was repressed in a medium containing glucose [61]. It was also found that reactivation of the Agr system via AIP addition or glucose depletion triggered detachment, destabilizing biofilms, only in agr-competent strains [61]. It has been shown that the activation of the Agr system results in upregulation of the Aur metalloprotease and the SpIABCDEF serine proteases, together with a downregulation of several surface proteins [62]. In fact, the inhibition of serine proteases and an S. aureus aur mutant, or a double mutant strain deficient in the aur gene and SpI genes, showed reduced detachment and a stable biofilm after the addition of AIPs, indicating that the Agr system destabilizes biofilms via the action of extracellular proteases.
This entry is adapted from the peer-reviewed paper 10.3390/diseases11040160
This entry is offline, you can click here to edit this entry!