When research on osteogenic differentiation in dental follicle cells (DFCs) began, projects focused on bone morphogenetic protein (BMP) signaling. The BMP pathway induces the transcription factor DLX3, whichh in turn induces the BMP signaling pathway via a positive feedback mechanism. However, this BMP2/DLX3 signaling pathway only seems to support the early phase of osteogenic differentiation, since simultaneous induction of BMP2 or DLX3 does not further promote differentiation. Data showed that inhibition of classical protein kinase C (PKCs) supports the mineralization of DFCs and that osteogenic differentiation is sensitive to changes in signaling pathways, such as protein kinase B (PKB), also known as AKT. Small changes in the lipidome seem to confirm the participation of AKT and PKC in osteogenic differentiation.
1. Introduction
Regenerative dentistry is an emerging area of dental research, and in recent years, numerous articles have appeared dealing with tooth development, tissue regeneration, organoids, and stem cell differentiation [
1,
2,
3,
4,
5,
6,
7,
8]. A tooth consists of three mineralized tissues derived from three different tooth germ tissues: enamel organ, dental papilla, and dental follicle. The dental follicle derives from the dental mesoderm, which originates from neural crest cells in early stages of mammalian development [
9]. This tissue is not only involved in the development of the periodontium, but is also responsible for tooth eruption [
10,
11,
12]. The dental follicle is collagenous tissue that is interspersed with small blood vessels. In the vicinity of these blood vessels, small cells express typical markers of dental stem cells, such as nestin [
13,
14,
15]. For periodontal development, it contains progenitor cells for the mineralizing cells, dental cementoblasts, and alveolar osteoblasts, as well as for periodontal soft-tissue fibroblasts that connect the mineralized alveolar bone and dental cementum [
16,
17,
18].
In contrast to the enamel organ, but similar to the dental papilla/pulp, the dental follicle can be obtained from wisdom tooth extractions, and so they were available for the isolation of somatic stem cells, which can be used for the regenerative dentistry [
19,
20]. In order to be able to isolate undifferentiated single cells, the tooth follicle separated from the extracted wisdom tooth not only has to be cut in small pieces, but also undergo further enzymatic treatment in order to dissolve the stable collagen matrix [
19]. Isolated single dental follicle cells (DFCs) built small colonies consisting of at least 50 cells after 14 days of cell culture in a standard medium for fibroblasts, which is typical for stem cells and osteogenic progenitor cells and has been demonstrated in numerous previous approaches [
21,
22]. Typical markers of dental follicle cells are the intermediate filament protein nestin and the protein marker of mesenchymal stem cells, STRO-1 [
14,
19,
23]. Differentiation approaches were performed to prove that these dental follicle cells are undifferentiated stem or precursor cells. For example, dental follicle cells could be differentiated into neural cells, which could be demonstrated by typical markers for neuronal cells [
24]. Most importantly, however, was the demonstration that DFCs can be differentiated into cells of the periodontium, such as cementoblasts or alveolar osteoblasts, and for regenerative medicine [
25,
26,
27,
28]. Therefore, an osteogenic differentiation approach under in vitro conditions was started. Surprisingly, this experiment showed that dental follicle cells not only differentiate into cells that produce mineralized tissue, but also into fibroblast-like cells that produce connective tissue [
13,
19]. It was possible to detect connective and bone tissue [
13]. However, this result also raises the question of whether it would not be possible to specifically differentiate the dental follicle cells into osteoblasts and avoid differentiation into fibroblasts.
To understand differentiation, the stem cell niche model, or rather molecular processes inside and outside the cell, must be examined in detail [
29,
30,
31,
32,
33,
34]. Although a number of molecules involved in osteogenic differentiation have been discovered, they need to be understood in context. The differentiation of dental follicle cells under in vitro conditions is induced by external growth factors. Examples include bone morphogenetic factor 2 (BMP2) and insulin growth factor 2 (IGF2) [
14,
35,
36]. In addition, differentiation depends on the composition of the extracellular matrix. Matrix proteins, such as collagen I and laminin, are of particular importance here. While collagen I induces early markers, such as alkaline phosphatase, in dental follicle cells, processes that favor mineralization will be aided by laminin [
37,
38].
Discovery of the BMP2/DLX3 Signaling Pathway
In order to discover further mechanisms, proteomics and transcriptomics were carried out [
39,
40,
41,
42]. In these analyses, differentially expressed genes were identified before and after the induction of osteogenic differentiation. Bioinformatics studies revealed that very few genes or proteins were previously associated with osteogenic differentiation, including BMP2, IGF2, and proteins/genes associated with the WNT signaling pathway [
43]. Other proteins that are associated with the BMP signaling pathway, such as SMAD proteins, were also found [
40]. We gave our full attention to this signaling pathway, along with the transcription factor DLX3, which was previously known to be regulated by BMP2 and involved in bone marrow-derived osteogenic progenitor cell differentiation and tooth development [
2,
44,
45,
46].
As expected, initial investigations showed that the expression of DLX3 is induced by BMP2 [
41]. Interestingly, upstream of DLX3, BMP2 not only activates SMAD proteins, but also protein kinase A (PKA), which, among other things, phosphorylates the β-catenin protein, which is a target protein of the WNT signaling pathway. After induction of osteogenic differentiation, β-catenin-like SMAD migrates into the cell nucleus. Here, β-catenin binds to its cofactor Lef, which in turn binds to the promoter of DLX3-like SMAD1/5 [
47]. These transcription factors induce the expression of DLX3, which not only participates in the expression of the osteogenic transcription factor RUNX2 [
46], but also induces the expression of BMP2, resulting in a positive feedback loop [
41]. On the other hand, contrary to what is assumed after β-catenin induction, canonical WNT signaling appears to have an inhibitory effect on DLX3 expression and osteogenic differentiation of DFCs, although how WNT affects BMP2/DLX2 signaling is unknown [
47]. In addition, BMP2 induces the activity of AKT (protein kinase B) [
48]. However, the AKT signaling pathway mediates BMP2-dependent induction of the alkaline phosphatase activity (osteogenic differentiation) via induction of the transcription factor EGR1 (Early growth response protein 1) [
48]. Interestingly, the parathyroid hormone-related protein (PTHrP) is secreted during osteogenic differentiation and has an inhibitory effect on early markers of osteogenic differentiation, such as ALP activity, but not on late markers, such as mineralization [
49]. While these signaling proteins are associated with the DLX3/BMP2 signaling pathway and are more likely to affect differentiation at an early stage, factors that may affect osteogenic differentiation at a later stage are presented below. One of these factors is protein kinase C.
2. Protein Kinase C (PKC) Signaling and Biomineralization
2.1. PKC in Osteogenic Progenitor Cells
One factor appears to be PKC, which can be divided into several groups, but only classical PKCs are involved in the osteogenic differentiation of DFCs [
50]. While the mechanism of the PKC signaling pathway, which is described below, is well known, little is known about its role in the osteogenic differentiation of DFCs. Interestingly, previous studies using both immortalized cell lines and somatic stem cells have demonstrated the involvement of PKC in osteogeogenic differentiation, with these studies showing that PKC can have both promoting and inhibitory effects. Miraoui et al. [
51] showed that fibroblast growth factor receptor 2 (FGFR2)-induced osteoblast differentiation in murine mesenchymal C3H10T1/2 depended on PKC activation. In contrast, Nakura et al. have shown that PKC inhibits osteogenic differentiation in mouse preosteoblastic cell line MC3T3-E1, but promotes cell proliferation [
52]. Similar conclusions were drawn with C2C12 cells by Lee and co-workers [
53], who showed that PKC inhibits osteogenic differentiation by regulating the transcription factor MSX2, which in turn inhibits the expression of the osteogenic transcription factor RUNX2. The more recent studies with somatic stem cells also showed similar results. Li et al. [
54], for example, showed that the inhibitory effect of miR-26a-5p on osteogenic differentiation of murine adipose-tissue-derived mesenchymal stem cells depends, among other things, on PKC inhibition. However, a study by Lotz et al. showed that PKC inhibition in human bone marrow-derived stem cells increased osteocalcin expression, but inhibited BMP2 expression [
55]. Results from another study showed that activation of PKC, specifically, PKCβ1, resulted in repression of muscle ring finger protein-1 (MURF1)-mediated ubiquitylation of the peroxisome proliferator-activated receptor γ2 (PPARγ2) transcription factor. Stabilized PPARγ2 proteins enhanced adipogenesis and consequently reduced osteoblastogenesis from MSCs, showing that PKC activation suppresses osteogenic differentiation [
56]. Similar inhibitory effects of PKC on osteogenic differentiation of PDL stem cells have recently been described. Wang et al. showed that Advanced glycation end product (AGE proteins) impaired the osteogenic potential via PKCβ2 [
57].
2.2. PKC Signaling in DFCs
First experiments with DFCs showed that the expression of classical PKCs, e.g., PKCα, is inhibited from day 7 of osteogenic differentiation, but these experiments also showed that manipulating PKC activity had little effect on ALP activity, which is an early marker of osteogenic differentiation peaking at day 7 [
50]. On the other hand, inhibition of classical PKCs supports the mineralization of DFCs, and this is the case even if PKCs were inhibited only a few days, but later than 1 week, after induction of differentiation, since inhibition of PKC occurring in the first week of osteogenic differentiation had no impact on the mineralization [
50]. These results suggest that PKC does not affect the early phase of differentiation. Therefore, the interaction of PKC with AKT, which had a positive influence on osteogenic differentiation in previous work [
48], was investigated [
50]. Interestingly, and in contradiction to this previous work, in this study, the induction of differentiation with both BMP2 and dexamethasone appears to inhibit AKT expression/activity [
50]. However, after inhibition of PKC, induction of osteogenic differentiation leads to activation of AKT [
50]. Thus, AKT seems to play a complex role in differentiation, which is why regulation of AKT activity during differentiation has been investigated. These experiments showed that AKT does not seem to have a direct impact on mineralization, since both the inhibitor MK2206 and the activator SC-79 inhibited mineralization in a dexamethasone-based differentiation medium [
50]. If one assumes that, as shown in this publication, PKC is essential for the activation of AKT, conversely, it should be investigated whether simultaneous inhibition of PKC and activation of AKT influences mineralization. When BMP2 was used for osteogenic differentiation, a stimulatory effect on the differentiation due to AKT inhibition was observed. These studies show that AKT is involved in the mechanism of differentiation. However, the importance of AKT on further signaling pathways, such as the BMP pathway, is ambiguous and depends on the cell line. Two recent studies on DFCs during osteogenic differentiation showed contradictory results regarding the activation of the BMP signaling pathway during differentiation. While Pieles et al. hypothesized an inhibitory effect of AKT on the BMP signaling pathway, Viale-Bouroncle and colleagues showed that AKT supports the BMP signaling pathway. These conflicting results suggest that AKT does not directly affect BMP signaling or that other, as yet unknown, signaling pathways influence AKT’s impact on BMP signaling. On the other hand, however, AKT activation seems to be essential for the inactivation of GSK-3β as a factor involved in the induction of β-catenin of the WNT pathway [
50]. The canonical WNT signaling pathway and, in particular, its transcription factor β-catenin, which is also induced by BMP2 in DFCs, play a role in the early phase of DFC differentiation [
47,
58]. While PKCs have a direct impact on non-canonical WNT signaling activation, the active form of β-catenin is induced after PKC inhibition [
50]. Since there have been results in recent years on the importance of the canonical WNT signaling pathway in DFCs, which had both a promoting and an inhibitory effect on osteogenic differentiation [
47,
58,
59,
60], this signaling pathway appears to have a modulating property for differentiation similar to the kinase AKT, which is characterized by the constitution of the individual DFC cell line. However, studies suggest that classical PKCs and canonical WNT signaling have a similar inhibitory effect on osteogenic differentiation of DFCs [
47,
50]. It will, therefore, be important for understanding differentiation to uncover by which biological processes classical PKCs are linked to the canonical WNT signaling pathway in DFCs during osteogenic differentiation.
A signaling pathway that is of importance for cell differentiation is the NFκ-B signaling pathway. This signaling pathway both inhibits and promotes the differentiation of osteogenic progenitor cells and is inhibited in dental follicle cells after induction of differentiation [
61,
62,
63,
64]. However, it appears to have an inhibitory effect on osteogenic differentiation of DFCs, as decreased mineralization was shown after NFκ-B induction with NFκ-B activator PMA. Interestingly, PMA activates NFκ-B, but independently of classical PKC [
65]. Novel PKCs, which do not influence osteogenic differentiation of DFCs, can directly activate the NFκ-B signaling pathway [
65]. However, a previous study showed that classical PKCs caused the activation of RELB of noncanonical NF-κB signaling, but not RELA of canonical NF-κB signaling, in cancer cells [
66]. So, this could be an explanation why it was not possible to induce osteogenic differentiation markers by inhibiting the canonical NFκ-B signaling pathway. Interestingly, regulation of AKT has only little influence on the expression of NFκ-B signaling pathway proteins after induction of osteogenic differentiation with dexamethasone. On the other hand, NF-κB can be induced by AKT, and both signaling pathways play important roles in cell viability [
67]. We believe that the NFκ-B signaling pathway does not play a significant role in osteogenic differentiation. Previous studies on the proteome and transcriptome did not find any significant evidence that the NFκ-B signaling pathway is involved in differentiation [
68]. It is possible that PMA induces PKC, but impairs cell viability or cell proliferation, as well [
69], and it is therefore possible that PMA suppressed the mineralization of DFCs by impairing cell viability without activation of NFκ-B signaling, which could support cell viability [
70]. This conclusion is supported by the following observation from a previous study. The inducer of the non-canonical WNT signaling pathway WNT5A, which supports cell viability during osteogenic differentiation [
71], inhibited the activation and expression of classical PKCs in DFCs. The results presented here can be summarized as follows [
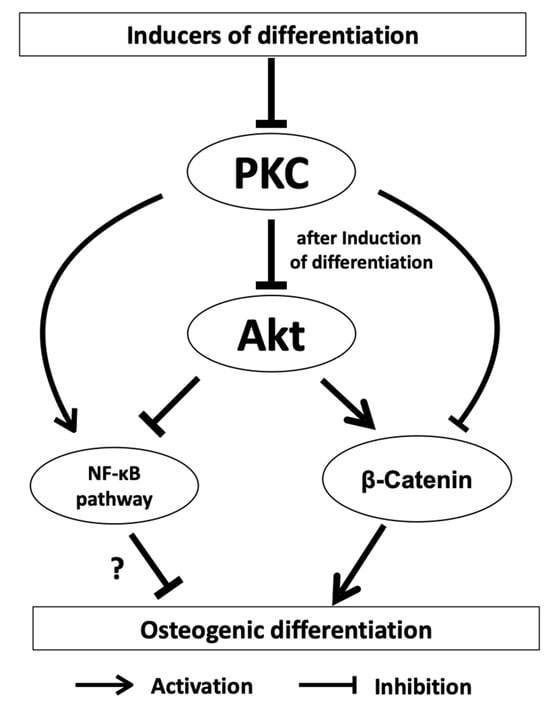
50]. PKCs inhibit osteogenesis specifically by regulating the kinase AKT, thereby affecting the downstream activity of β-catenin and the NF-κB signaling pathway (
Figure 1).
Figure 1. PKCs inhibit osteogenesis by regulating the kinase AKT, which influences downstream β-catenin and the NF-κB signaling pathway. The effect of the NF-κB signaling pathway on osteogenic differentiation is probably small (?).
The question, therefore, arises to what extent do PKCs influence crucial biological processes or are influenced by them, and what significance do these processes have for the osteogenic differentiation? If we look at the mechanism for the activation of classical PKC, it is striking that this cascade starts with the activation of a G protein-coupled receptor (GPCR), which activates phospholipase C (PLC) downstream. Activation of PLC cleaves the lipid phosphatidylinositol-4,5-bisphosphate (PIP2) by hydrolysis into diacylglycerol (DAG) and inositol-1,4,5-trisphosphate (IP3), which releases calcium as another cofactor for the activation of PKC [
72,
73]. DAG also plays an important role in the activation of PKC as a cofactor [
73], and its presence in the lipidome is closely related to the synthesis of lipids [
74]. While phosphorylation also plays an important role in PKC activation, the interaction of classical PKC and DAG demonstrates the connection between signaling pathways and crucial biological processes, such as lipid metabolism, which will be discussed below in its closer relation to osteogenic differentiation of DFCs.
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines11102787