Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Oncology vaccines represent a breakthrough in oncology, using the principles of immunotherapy to stimulate the body's immune system to fight cancer, providing a more targeted approach with fewer side effects than traditional treatments such as surgery, radiotherapy, and chemotherapy.
- TME
- cancer immunology
- oncology vaccines
- combination therapies
1. Introduction
According to a comprehensive epidemiological survey conducted in 2020, malignancy has been the leading cause of morbidity and mortality worldwide, with a shocking incidence of approximately 19,3 new cancers and nearly 10,1 deaths during the year [1]. Health and health systems face enormous challenges. However, new hope for cancer treatment is in the form of oncology vaccines. Oncology vaccines represent an oncology breakthrough that uses the principles of immunotherapy to stimulate the body's immune system to fight cancer, providing a more targeted approach with fewer side effects than conventional treatments such as surgery, radiotherapy, and chemotherapy [2].
The evolution of cancer vaccines has unfolded over decades, with each era marked by key advances (Figure 1). The journey began with William Coley in the late 19th century, who developed Colley toxin after noticing that infections often cause tumors in cancer patients to shrink [3]. This approach harnesses the immune system to fight cancer – a concept further expanded by Lewis Thomas and Frank McFarlane Burnett in the mid-20th century, when they introduced the ideas of cancer immune editing and tumor immunity [4]. The dawn of a clinically approved cancer vaccine came in 2010 with the FDA approval of sipuleucel-T, the first therapeutic cancer vaccine against prostate cancer [5]. Subsequent eras continue to this day and are characterized by the development of modern cancer vaccines and checkpoint inhibitors, such as the combination therapy approved in 2021 for the treatment of lymphoma [6]. Currently, cancer vaccine development emphasizes personalization, and multiple strategies are being studied, including neoantigen vaccines, DNA/RNA vaccines, and viral vectors, aimed at tailoring treatments to individual patients' tumors.
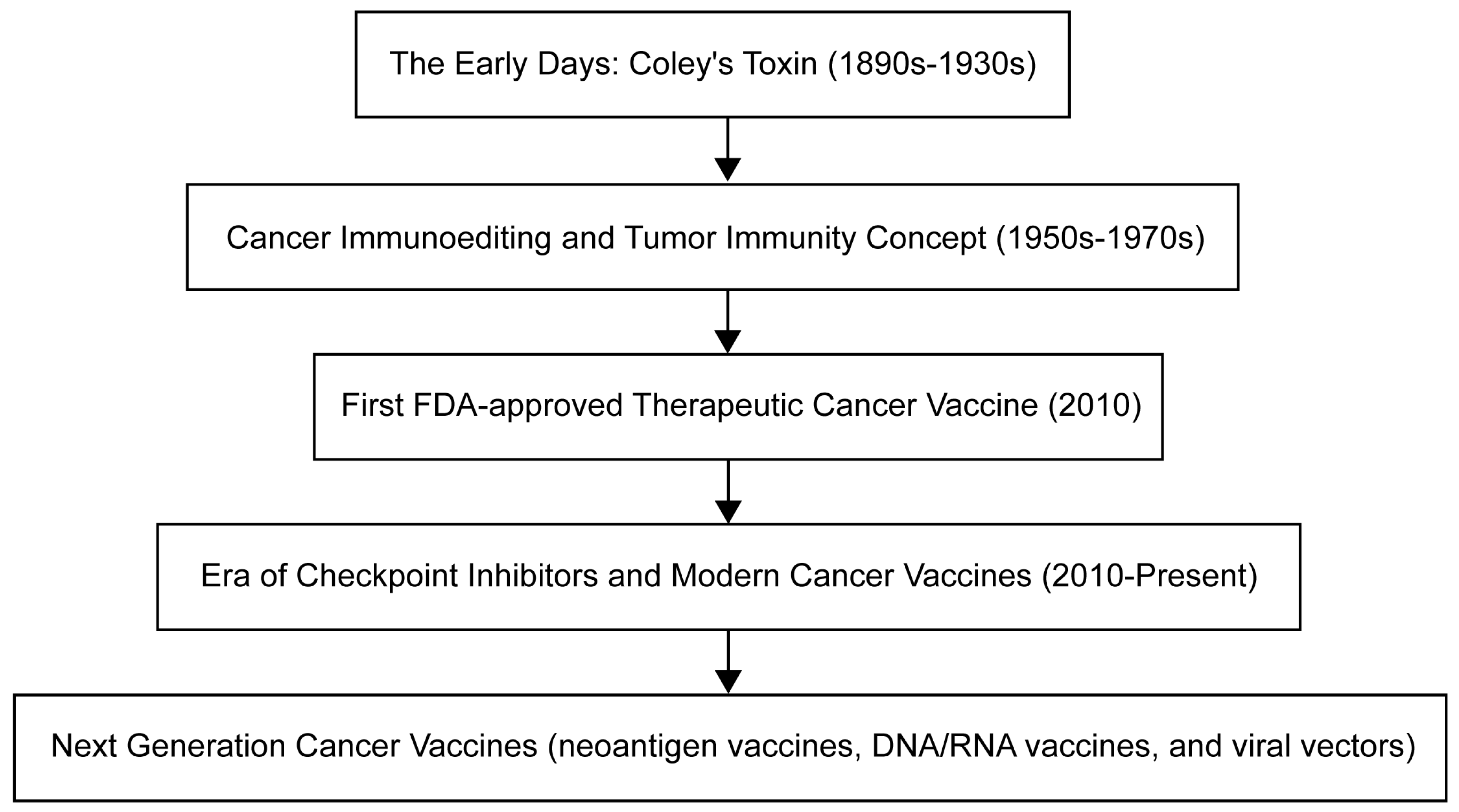
Figure 1. Timeline for oncology vaccine development.
In-depth study of the mechanistic complexity in which the tumor microenvironment (TME) in which tumors exist and evolve plays a key role in tumor progression. As a result, TME has become a scientific research center in the field of anti-cancer therapeutic drug evolution [7]. As proof of contemporary medical innovation, tumor vaccines have been carefully designed to harness the innate and adaptive nature of the host immune system to target abnormal cells. These groundbreaking formulations strategically regulate TME, thereby wisely minimizing collateral damage to healthy tissue [8].
The field of cancer immunotherapy is undergoing significant advances with the development of various oncology vaccines, each with its own unique advantages and challenges. Although peptide vaccines are effective in eliciting an immune response, adjuvants are often required [9]. DNA/RNA-based vaccines allow endogenous antigen generation but face barriers to delivery and expression [10][11]. Viral vector-based vaccines can stimulate a robust immune response, but are limited by safety concerns and pre-existing immunity [12]. While dendritic cell-based vaccines can induce robust responses, they require complex production processes and different patient responses [13]. Whole-cell-based vaccines address production and standardization issues while harnessing whole tumor cells to trigger an immune response [14]. A deep understanding of these vaccines and their limitations is critical for their effective deployment in cancer immunotherapy [15]. Current studies recognize the limited efficacy of oncology vaccines and are seeking to enhance therapeutic efficacy by combining them with other therapies such as immune checkpoint inhibitors, chemotherapy, radiotherapy, targeted therapy, and oncolytic viral therapy [16][17][18][19]. At the same time, the innovative field of personalized cancer vaccines is gaining traction, providing tailored immune responses to individual-specific tumors [8]. Despite some challenges, including the complexity of neoantigen identification and resource-intensive production, technological advances promise a faster and more affordable future for personalized cancer vaccines [20][21][22][23].
2. Tumor microenvironment
The tumor microenvironment (TME), which is critical to cancer onset and progression, is a headache for scientists. It is not only dynamic, but also very complex, shaping the heterogeneity of tumors. TME is a mixture of cellular and noncellular components (eg, cancer cells, stromal cells, immune cells, ECMs) and signaling molecules (eg, growth factors and cytokines) [24][25]. ECM is a tangled network of proteins, glycoproteins, and proteoglycans that provide structure and shape cell behavior through biochemical and biomechanical signals. When cancer alters the composition of the ECM, it can promote angiogenesis, immune evasion, and treatment resistance [26][27]. For example, as seen by overexpression of lysine oxidase (LOX), ECM hardness increases, promotes cell proliferation and survival through mechanisms such as FAK activation, and also enhances tumor aggressiveness [28]. Similarly, accumulation of hyaluronic acid is often associated with a poor prognosis, which can promote tumor growth and promote immune escape [29]. Alterations in matrix metalloproteinase (MMP) expression can remodel the ECM, favoring tumor invasion by releasing growth factors [30][31]. In addition, specific ECM proteins such as elastin, laminin, tenascin-C, and periosteum protein support tumor cell migration and survival when overexpressed [32]. Notably, changes in collagen orientation lead to a fibrous arrangement that provides a pathway for enhanced tumor cell migration, which often indicates a higher risk of metastasis [33]. ECM is also involved in the secretion of various growth factors such as transforming growth factor-β (TGF-β), interleukin-1β (IL-1β), IL-6, tumor necrosis factor-α (TNF-α), and vascular endothelial growth factor (VEGF), which are secreted by the individual cells of TME and can initiate tumor cell growth, survival, migration, angiogenesis, and epithelial-mesenchymal transformation (EMT). This is achieved by modulating its specific receptors and stimulus signaling pathways [34] (Figure 2). This intricate interaction between ECM and tumor cells not only drives cancer progression, but also presents treatment challenges due to factors such as drug permeability barriers and activation of cell survival pathways.
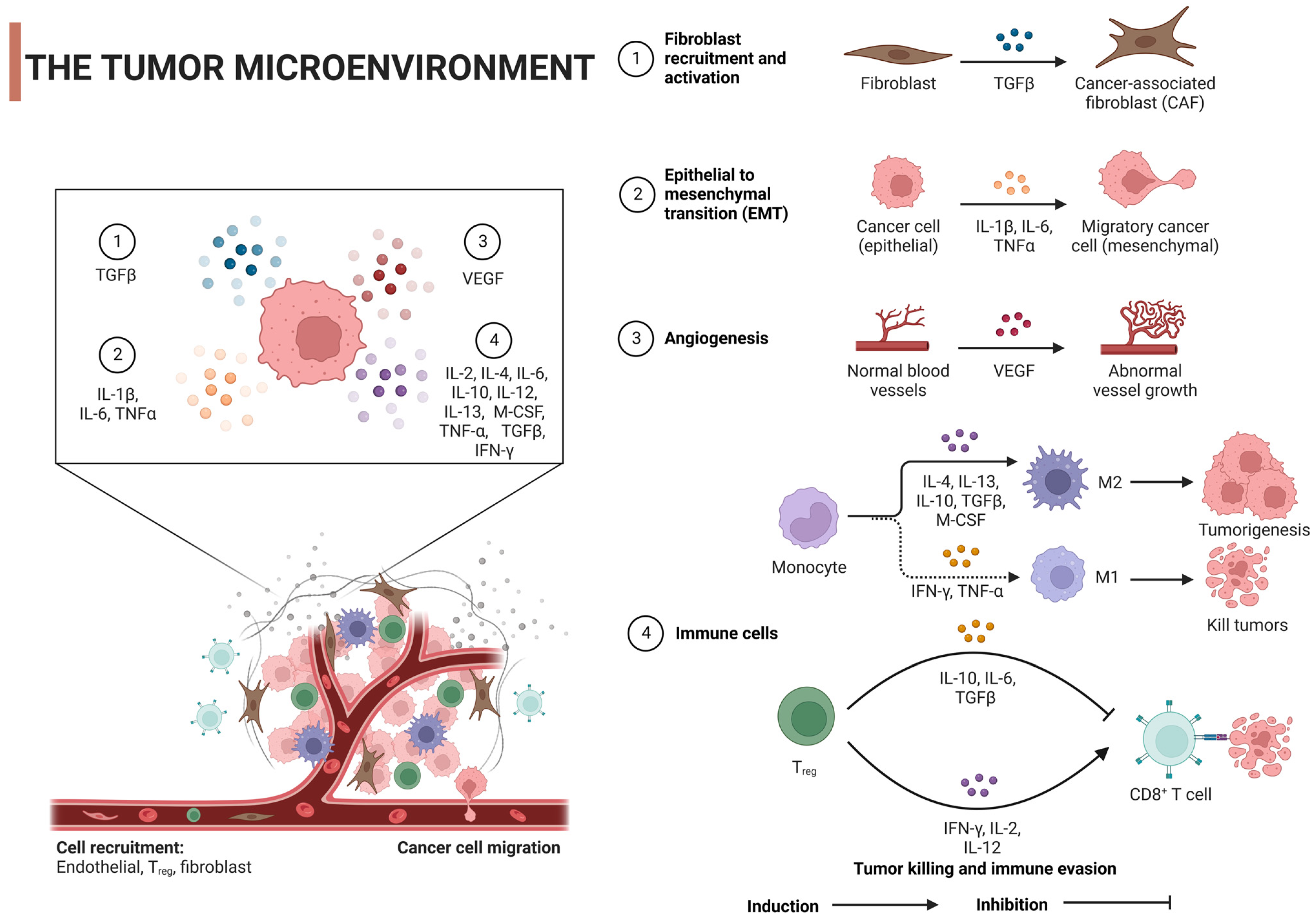
Figure 2. Regulatory mechanism of tumor microenvironment on tumor cells: TGF-β, transforming growth factor-β; IL, interleukin; VEGF, vascular endothelial growth factor; TNF-α, tumor necrosis factor α; IFN-γ, interferon γ; M-cerebrospinal fluid, macrophage colony-stimulating factor; Tregs, regulatory T cells. Created with BioRender.com.
Similarly, immune cell function is formed by a mixture of elements. Tumor cells secrete immunosuppressive cytokines and checkpoint ligands that alter the immune response [35,36], while diseases such as hypoxia and acidic pH caused by changes in tumor metabolism can inhibit immune activity [35][36]. At the same time, tumor glucose consumption, extracellular matrix changes, and metabolic competition due to immunosuppressive cell recruitment can inhibit a potent immune response [37]. Direct cell-cell interactions within TME, the impact of the gut microbiome on tumor immunity, therapeutic interventions, and the presence of chronic inflammation further modulate the balance between protumor and antitumor effects [38][39]. The interaction of these factors determines the complexity and dynamics of the immune response within TME.
With regard to immune cells, such as TAM and T cells, these cells can oscillate bidirectionally [40][41]. They can drive tumor growth or stop tumor growth. TME phenotype and cytokines control the expression of all these cellular and non-cellular components [42]. TAM is a key cell in TME with functional plasticity that promotes or inhibits tumor progression. This is largely influenced by the various cytokines present in TME, resulting in TAM adopting one of two main phenotypes: classically activated (M1) macrophages with antitumor activity and surrogate-activated (M2) macrophages, usually protumors [43]. Certain cytokines, such as interferon-γ (IFN-γ), produced primarily by T cells and NK cells, can drive the progression of TAM to the M1 phenotype. These M1 macrophages are characterized by their ability to present antigens, kill tumor cells, and produce pro-inflammatory cytokines such as IL-12 and TNF-α, thereby enhancing antitumor immunity [43]. TNF-α also plays a role in inducing the polarization of macrophages M1, thereby increasing their tumoricidal activity [44]. However, other cytokines, such as IL-4 and IL-13, both produced primarily by Th2 cells, can induce macrophage polarization to the M2 phenotype, which is often associated with tissue repair, immunomodulation, and tumor promotion [45]. Similarly, IL-10, an anti-inflammatory cytokine, drives TAM to the M2 phenotype, resulting in macrophages' normally immunosuppressive effects and the production of factors such as VEGF and EGF that promote tumor growth and angiogenesis [43]. Finally, TGF-β is known to induce M2 polarization and also has a variety of tumor-promoting effects, including promoting immune evasion, tissue remodeling, and angiogenesis [46].
T cells, especially cytotoxic T cells (CTLs) and helper T cells, play a key role in anti-tumor immunity, and their activation, proliferation, and function are affected by various cytokines in TME. Several cytokines propel T cells to fight tumors. Interferon-γ (IFN-γ), produced primarily by Th1 cells and CTL, enhances the cytotoxic activity of CTL, promotes its tumor killing capacity, and increases the expression of MHC class I molecules on tumor cells, making them more susceptible to CTL-mediated killing [47]. IL-2 plays a vital role in the growth and differentiation of T cells, and it is commonly used in cancer immunotherapy, mainly by promoting the proliferation and activation of CTL and NK cells to enhance the immune system's ability to fight cancer [48]. IL-12 guides the differentiation of naïve T cells into Th1 cells, which generate IFN-γ and promotes CTL-mediated tumor destruction [49]. On the other hand, some cytokines induce tumorigenesis. IL-10 is an immunosuppressive cytokine that hinders the function of effector T cells and antigen-presenting cells, potentially promoting tumor growth by inhibiting antitumor immunity [50]. TGF-β can inhibit the function of T cells and other immune cells, helping tumor immune evasion. It also exhibits tumor-promoting effects in the later stages of cancer, promoting tumor cell invasion and metastasis, and initiating angiogenesis [51]. Finally, IL-6 can aid tumorigenesis by promoting chronic inflammation, cell survival, and angiogenesis, and it is involved in differentiating T cells into Th17 cells, which in some cases have been implicated in promoting inflammation and tumor growth [52].
TME offers a promising therapeutic pathway for cancer treatment. Immunosuppressive cytokines such as TGF-β when suppressed, may restore the antitumor immune response and reduce metastasis [53]. Although IL-10 generally suppresses TME immunity, its subtle effects suggest potential benefits in regulating its levels, and angiogenesis inhibition and immunosuppressive VEGF have led to FDA-approved therapies such as bevacizumab [54][55]. In terms of immune stimulation, IL-2, known to promote T cell growth, has seen therapeutic applications, and despite side effects, the effective activation of immune cells by IL-12 hints at its combinatorial therapeutic potential [56][57]. In addition, checkpoint inhibitors targeting the PD-1/PD-L1 axis, such as pembrolizumab, rejuvenate depleted T cells to counteract tumors [58].
Together, TME has had a significant impact on the success of cancer vaccines. While vaccines aim to activate immune cells that target tumor antigens, the immunosuppressive nature of TME hinders these activated T cells. Factors such as altered tumor antigen presentation, decreased expression of MHC molecules, and expression of immune checkpoints further impede vaccine-induced responses [59][60]. To improve the efficacy of cancer vaccines, researchers are exploring combination therapies with checkpoint inhibitors, ways to reduce the number of immunosuppressive cells in TME, strategies to break through the physical barrier of TME, binding of potent adjuvants, cytokine regulation, and the development of personalized vaccines for individual tumor antigen profiles [61][62][63].
3. Oncology vaccines
3.1. Cancer immunology
Adaptive immunity or acquired immunity is a highly specific and persistent defense against pathogens and abnormal cells, administered primarily by T and B lymphocytes [64]. This type of immunity is known for its immune memory, which provides long-lasting resistance to previously encountered things, such as pathogens or antigens [65]. Studying the ins and outs of adaptive immunity is critical to making vaccines and targeted immunotherapies. These breakthroughs offer great potential for the prevention and treatment of a range of diseases, from infections and autoimmune diseases to cancer [66][67][68].
In the early stages of cancer, the immune system is involved in a process called immune surveillance, where it hunts down and destroys abnormal cells, preventing tumor formation [69][70]. But cunning cancer cells use different techniques to evade the immune system and help tumors grow and progression [71]. By studying these immune evasion strategies, scientists have proposed new immunotherapies such as immune checkpoint inhibitors (ICIs) and adoptive cell transfer. These innovative therapies are designed to improve the body's ability to fight tumors and overcome cancer's ability to evade the immune system [72].
Immunotherapies, such as ICI and chimeric antigen receptor (CAR) T-cell therapy, have completely changed the treatment of cancer by using the immune system to fight cancer cells [73]. But even with these amazing advances, some patients respond poorly to immunotherapy and may even develop resistance to immunotherapy, which means that more needs to be learned about how the immune system and cancer interact [74]. Gaining this knowledge will help create new immunotherapy strategies and identify predictive biomarkers that lead to better patient outcomes [75][76].
3.2. Mechanism of Action
The fundamental function of cancer vaccines lies in their ability to mobilize innate and adaptive immune responses to identify, fight, and eradicate tumor cells [20]. The following discussion will provide a comprehensive analysis of its intricate mechanisms of action (Figure 3).
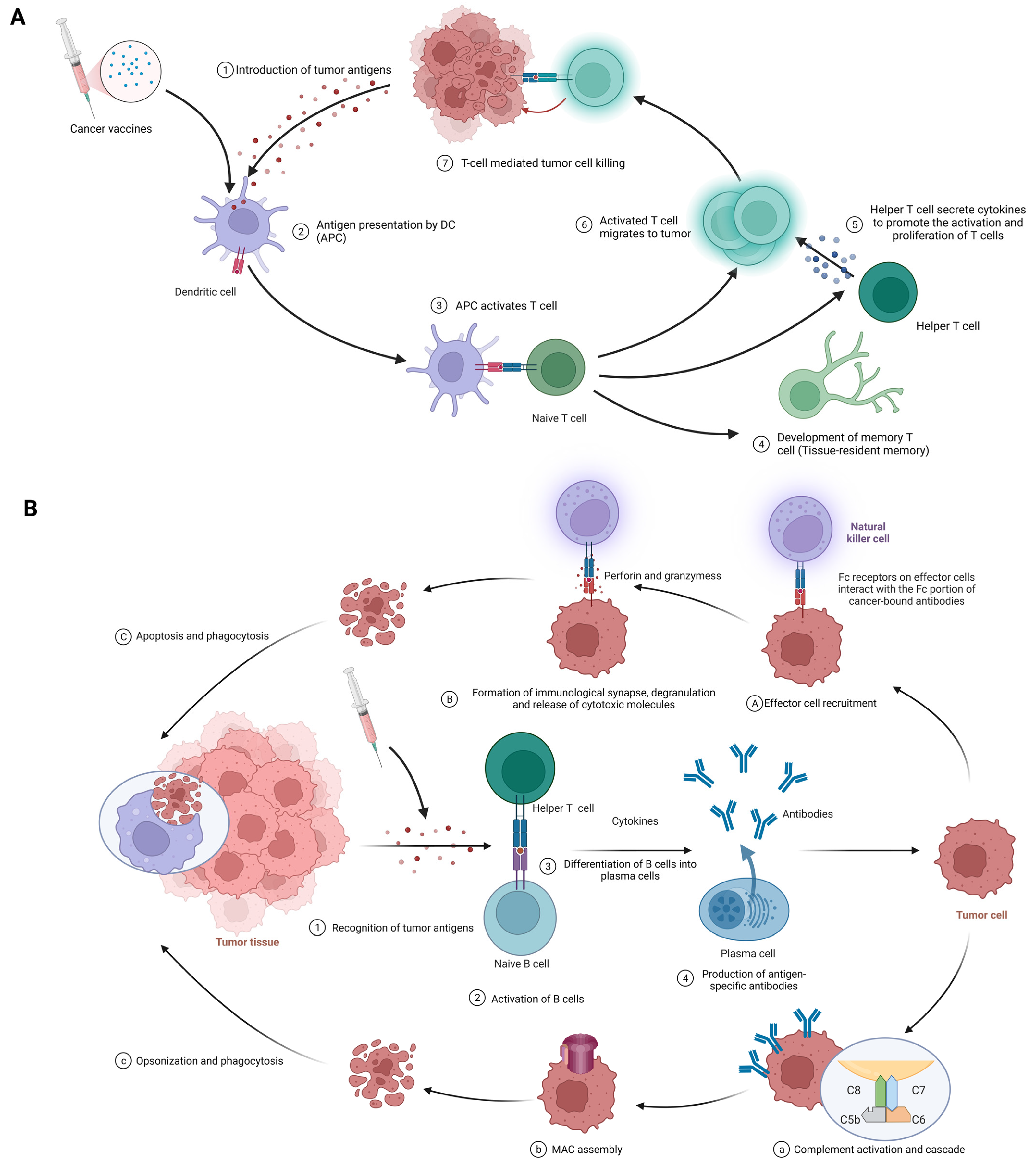
Figure 3. (A) Cellular immune response against cancer (step 1 to step 7); APCs, antigen-presenting cells; APC, antigen-presenting cell. (B) Humoral immunity response against cancer (step 1 to step 4). A–C; antibody-dependent cell-mediated cytotoxicity; a–c: complement-dependent cytotoxicity; MAC, membrane attack complex. Created with BioRender.com.
3.2.1. Cellular Immunity
The procedure of eliciting a cellular immune response against cancer, exemplified by the use of cancer vaccines, is complex and sequential. It commences with the delivery of tumor antigens and concludes with the activation of humoral immunity. The following is a detailed breakdown of each phase in this process (Figure 3A).
Introduction of tumor antigens: With cancer vaccines, referring to tumor antigens being introduced to antigen-presenting cells (APCs) like dendritic cells. Options include whole tumor cells, peptides, proteins, DNA, mRNA, or even dendritic cells loaded with tumor antigens or packing tumor-derived genetic material. Tumor vaccines play a role in this step. This crucial first step gets the ball rolling in the immune response against cancer [18][77].
Antigen processing and presentation: APCs capture, process, and present tumor-derived peptides on their surface. Dendritic cells efficiently cross-present exogenous antigens to both MHC class I and II molecules, activating both CD8+ cytotoxic T lymphocytes (CTLs) and CD4+ helper T cells [18][78].
Activation of T cells: Presentation of tumor antigens by APCs activates and expands antigen-specific T cells. T-cell activation requires antigen recognition via the T-cell receptor (TCR) and costimulatory signals provided by the interaction between costimulatory molecules on APCs and their receptors on T cells [79][80].
CTLs and helper T cells in action: CTLs directly kill tumor cells by recognizing and binding to MHC class I molecules presenting tumor antigens, while helper T cells produce cytokines that support CTLs’ activation, proliferation, and differentiation. CD4+ helper T cells also provide help to B cells, facilitating antibody production and enhancing the function of APCs and CTLs [2][40].
Stimulation of humoral immunity: Cancer vaccines induce B cells to produce antigen-specific antibodies. These antibodies target and eliminate tumor cells [15][81].
3.2.2. Humoral immunity
The procedure begins with the identification of tumor antigens and ultimately achieves effector function mediated by antibodies. Here are the details of each stage in this process. (Figure 3B).
Recognition of tumor antigens: Like cellular immunity, tumor cells have tumor-associated antigens (TAAs) and tumor-specific antigens (TSAs) that the immune system can recognize [82].
Activation of B cells: B cells detect and recognize tumor antigens through their B cell receptor (BCR). With the addition of some co-stimulatory signals from helper T cells, B cells are activated to proliferate and differentiate [70].
B cells differentiate into plasma cells: Upon activation, B cells transform into plasma cells, which are specifically designed to produce large amounts of specific antibodies against recognized tumor antigens [82].
Antigen-specific antibody production: Plasma cells produce antigen-specific antibodies that pass through the bloodstream and attach to tumor antigens on cancer cells [83].
Effector function of antibodies: Once bound, antibodies employ various strategies against tumor cells, such as:
-
Antibody-dependent cell-mediated cytotoxicity (ADCC) (Figure 3A(1-3)): In the first phase, effector cell recruitment: Fc receptor-carrying immune cells, such as natural killer (NK) cells, macrophages, and neutrophils, are attracted to the tumor site through interactions between their Fc receptors and Fc fragments of antibodies bound to malignant cells [84][85]. In the next stage, the formation of immune synapses, degranulation, and the release of cytotoxic molecules: Immune synapses are formed when the Fc receptor on effector cells interacts with the Fc portion of the cancer-binding antibody, enabling the targeted release of cytotoxic molecules [86]. Effector cell degranulation ensues, eventually eliminating cancer cells by perforin and granzyme [87]. Finally, phagocytosis: After apoptosis, macrophages engulf cancerous debris, facilitating clearance and preventing the spread of harmful cellular components [88].
-
Complement-dependent cytotoxicity (CDC) (Figure 3B(1-4)): In the first phase, complement activation and cascade reaction: the Fc moiety of the binding antibody binds to C1q (complement component 1,q subcomponent), stimulating the classical complement pathway and the formation of C3 convertase complexes [89][90]. In the next stage, membrane attack complex (MAC) assembly: complement protein cleavage and activation produces MAC, including C5b, C6, C7, C8, and C9 [91]. After that, cell lysis: MAC perforates the cancer cell membrane, resulting in uncontrolled ion and water movement, cell lysis, and death [91]. Finally, opsonics and phagocytosis: when complement activation promotes opsonship, cancer cells are targeted for destruction by phagocytes [90].
-
Neutralization of growth factors and inhibition of signaling pathways: Antibodies impede tumor cell proliferation, promote growth factors, and block signaling pathways that are critical for cancer cell survival and invasion [15].
4. Types and Characteristics of Cancer Vaccines
4.1. Peptide Vaccines
Peptide vaccines represent a potential cancer immunotherapy approach that uses short amino acid sequences derived from tumor-specific or tumor-associated antigens (TAAs) to elicit a targeted immune response against malignant cells [92]. TAAs, including differentiated antigens, overexpressed antigens, cancer/testis antigens, and mutant antigens, are viable targets for a variety of immunotherapy techniques, such as cancer vaccines, adoptive T cell therapies, and immune checkpoint inhibitors [93][94]. Peptide vaccines offer several advantages over whole protein or live attenuated vaccines, including ease of synthesis, specificity, and good safety because peptide vaccines are less likely to elicit an autoimmune response [95][96]. However, they have encountered limitations such as poor immunogenicity, rapid degradation in vivo, weak CD4+ T cell response, and challenges associated with immune evasion and tumor-induced immunosuppression [95]. To overcome these limitations, strategies include the addition of adjuvants, optimization of peptide sequences, enhancement of stability and immunogenicity with vectors, and combination of peptide vaccines with other immunotherapies [92].
Currently, relatively mature peptide vaccines include Nelipepimut-S (NeuVax), CIMAvax-EGF and MUC1-based peptide vaccines. Nelipepimut-S, also known as NeuVax, is a peptide vaccine targeting HER2/neu-expressing cancer cells primarily in patients with early-stage HER2 1+ and 2+ breast cancer who are not candidates for standard HER2 therapy [97]. It binds the E75 peptide of HER2/neu to GM-CSF as an adjuvant to enhance the immune response [98]. Although its scope may cover other HER2/neu cancers, such as ovarian and gastric cancers, clinical development has encountered obstacles [99]. Recent phase III trial results suggest that Nelipepimut-S continues to exhibit good efficacy and tolerability in breast cancer patients [100]. Similarly, CIMAvax-EGF, which targets non-small cell lung cancer (NSCLC) epidermal growth factor, merges recombinant EGF with protein carriers. It has shown promise for prolonging the lives of patients with advanced lung cancer and has completed phase III trials [101][102]. In addition, MUC1-based peptide vaccines focus on abnormally expressed glycoproteins in cancers such as breast and pancreatic cancer [103]. Although some agents have entered phase I and II trials with good safety and immunologic indicators, eliciting a robust clinical response remains complex, and combination therapies need to be explored [104][105]. Collectively, these vaccines represent cutting-edge cancer treatments, each with its own unique targets and stages of development.
4.2. DNA/RNA-Based Vaccines
DNA/RNA-based tumor vaccines work by delivering genetic material encoding tumor antigens to host cells. This approach utilizes a variety of vectors, such as viral vectors, lipid nanoparticles, or naked nucleic acids, to stimulate the immune system to recognize and destroy cancer cells, resulting in an adaptive immune response [106]. These vaccines offer several advantages over traditional methods, including safety, ease of manufacture, robust immune response induction, and adaptability to modifications. They also offer the possibility of individualization to meet each patient's unique tumor profile [107][108]. Although they are promising, challenges such as efficient delivery and uptake of DNA/RNA into cells, the risks of autoimmune responses, and the limitations of personalized costs and time pose significant barriers [109][110][111].
However, recent advances are encouraging, including several vaccines and technological advances in clinical trials that have improved the effectiveness of vaccine delivery [112]. The field expects to use advances in genome sequencing, bioinformatics, and nanotechnology to overcome current limitations, eager to combine these potent vaccines with other immunotherapy strategies to achieve total cancer eradication [113][114]. Among them, technologies such as lipid nanoparticles (LNPs) are pioneering in mRNA COVID-19 vaccines and are being applied to cancer vaccine development to facilitate the delivery of tumor-specific antigens [115][116]. Electroporation and viral vectors (eg, adenovirus) enhance DNA/RNA uptake, while nonviral nanovectors and microneedle patches are designed to enhance this delivery without inducing a strong anti-vector response [117][118]. To reduce autoimmune risk, researchers emphasize tumor-specific antigen selection, sequence optimization to reduce cross-reactivity, and transient expression techniques such as those inherent to mRNA vaccines [119][120]. In addition, tolerable adjuvants and nanoparticles tailored for targeted delivery are being utilized to fine-tune the immune response, maximizing antitumor efficacy while minimizing collateral damage to healthy tissue [119][121]. Taken together, these innovations underscore the evolving landscape of cancer vaccine design, balancing effective tumor targeting with patient safety. CV9104 is an mRNA-based cancer vaccine targeting prostate cancer [122]. It progressed to a phase II trial of metastatic castration-resistant prostate cancer (mCRPC) [122]. This vaccine represents an innovative application of mRNA in oncology.
4.3. Viral Vector-Based Vaccines
Viral vector-based oncology vaccines are a promising development in cancer immunotherapy [11]. These vaccines use the innate ability of the virus to infiltrate host cells and efficiently deliver tumor antigens, eliciting a strong targeted immune response [123][124][125]. This approach encourages host cells to produce tumor-specific or relevant antigens after infection, resulting in these antigens being exposed to T cells and subsequently initiating a strong defense against tumor cells [126]. One of the significant advantages of these vaccines is their ability to trigger effective cellular and humoral immune responses. They can also be designed to express multiple tumor antigens, expanding their range and enhancing the potency of the immune response [127][128]. Despite these benefits, certain challenges need to be addressed, such as the impact of pre-existing viral vector immunity and the logistical issues associated with mass production [129][130].
Among these challenges, pre-existing immunity to viral vectors poses significant challenges for their use in tumor vaccines, as the immune system may neutralize the vector before the vector has a therapeutic effect [131]. To address this, researchers are exploring a range of strategies: using rare or novel viral vectors with limited human exposure, pseudotyping to alter viral envelope proteins, adopting heterologous prime-augmentation strategies with different vectors, genetic modification of viral capsids to reduce recognizability, co-administration with immunomodulators to temporarily suppress certain immune responses, choosing non-intravenous delivery routes such as intratumoral administration to avoid high antibody concentrations, and adjusting the dose, or using high vector doses to overcome neutralization, Low doses are either repeated to evade immunodetection [132][133][134][135]. In addition, adjuvants are being explored to shift the focus of the immune response from the vehicle to the delivered tumor antigen [136][137]. These multifaceted approaches aim to optimize the efficacy of viral vector-based oncology vaccines in the face of pre-existing immunity.
OncoVEXGM-CSF or T-VEC (T-VEC) is an oncolytic HSV-1 vaccine modified for tumor selectivity and GM-CSF production, primarily against melanoma [138]. Following a successful phase III trial, the drug received FDA approval for unresectable recurrent melanoma [138]. CG0070 is another adenovirus-based vaccine for selective replication in cancer cells with defects in the Rb pathway and targeting bladder cancer [139]. LV305 is a lentivirus-based vaccine that delivers the NY-ESO-1 antigen gene to dendritic cells to target NY-ESO-1-expressing cancers such as melanoma and sarcoma [140]. JX-594 or Pexa-Vec is a vaccinia virus-based vaccine modified to express GM-CSF and selectively target cancer cells with high thymidine kinase activity, and several trials have been conducted, including a phase III trial for hepatocellular carcinoma [141][142][143]. These represent innovative intersections of viral therapy and immunotherapy in oncology.
4.4. Dendritic Cell-Based Vaccines
Dendritic cells (DCs) mediate immune responses by connecting the innate and adaptive immune systems and are critical in antigen presentation and subsequent T cell activation, so they have been found to be substantially relevant to cancer immunotherapy strategies [18][144]. This stems from a DC-based cancer vaccine that utilizes DCs loaded with tumor-associated antigens (TAAs) to promote a robust immune response against cancer cells [145]. Methods for loading these DCs with TAA are varied, from the use of tumor lysates and synthetic peptides to mRNAs encoding tumor antigens [146]. Clinical trials highlight the promise of these DC-based vaccines in a variety of cancers. Despite this exciting potential, challenges remain, including technical difficulties in DC vaccine production, changes in vaccine efficacy, immunosuppressive tumor microenvironments, and lack of reliable biomarkers for patient selection [147]. However, recent advances in personalized cancer immunotherapies, such as neoantigen-based vaccines, offer promising opportunities for DC-based vaccines that may be enhanced in combination with other therapeutics [8]. In a recent study, researchers introduced a metabolic glycan labeling technique using azigo to enhance the DC vaccine [148]. This method promotes not only DC activation and antigen presentation, but also efficient conjugation of cytokines [148]. In addition, it is expected to be widely used in various tumors, provide a platform for regulating the interaction between DCs and other immune cells, and enhance the anti-tumor efficacy of dendritic cell vaccines.
Prominent representatives include Provenge and DCVax-L. Provenge (sipuleucel-T) is an FDA-approved autologous cell immunotherapy for advanced prostate cancer [5]. It uses peripheral blood mononuclear cells (PBMCs) of patients exposed to the fusion protein PA2024, which binds antigens from prostate cancer cells to the immune activator GM-CSF, eliciting an immune response against antigen-expressing prostate cancer cells [149][150][151][152][153]. DCVax-L, on the other hand, is an autologous dendritic cell vaccine against glioblastoma multiforme (GBM) [154]. The vaccine prepares the patient's dendritic cells by loading tumor lysates from the patient's own tumor tissue, enabling the immune system to recognize and attack the corresponding cancer cells [154]. Both vaccines use dendritic cells to target cancer, but their clinical history and disease targets differ (Table 1).
4.5. Whole Cell Vaccines
Whole-cell-based vaccines provide a comprehensive approach to cancer immunotherapy by binding a large number of tumor-associated antigens to stimulate an efficient immune response. Mechanistically, these vaccines utilize irradiated tumor cells (autologous or allogeneic) to expose the immune system to the full antigen pool of the tumor [155], thereby inducing a specific and multivalent immune response against a range of tumor antigens [156]. This strategy provides a broad spectrum of known and unknown tumor antigens, avoids antigen loss or down-regulation (the typical escape mechanism employed by tumors [155]), and eliminates the need to identify specific antigens for each patient, which can be time-consuming and expensive [157]. However, there are limitations. The production of autologous whole cell vaccines can be labor-intensive and individualized, so tumor cells need to be isolated and cultured from each patient [157]. Given that the immunosuppressive tumor microenvironment limits vaccine efficacy, these vaccines often need to be co-administered with adjuvants or immunomodulators to improve their immunogenicity [8][157]. Potential autoantigen-induced autoimmunity in vaccine formulations is also of concern [156]. Therefore, while whole-cell-based vaccines offer a promising approach to cancer immunotherapy, these strategies need to be further optimized and refined to address these challenges and limitations.
Representatives include GVAX, Canvaxin, and Oncophage. GVAX is a whole-cell tumor vaccine, utilizing tumor cells genetically modified to secrete GM-CSF (an immune stimulant), and has been explored for cancers like pancreatic and prostate cancers, with mixed outcomes in later-phase trials [158][159]. Canvaxin, aimed at melanoma, combines irradiated autologous and allogeneic melanoma cells with the BCG adjuvant, but it failed to show significant survival benefits in a phase III trial for advanced melanoma [113][160]. Oncophage (Vitespen) is derived from patient-specific tumor heat shock proteins (HSPs) and primarily targets renal-cell carcinoma and melanoma [161][162]. It completed phase III trials with mixed results but secured approval in Russia for the treatment of kidney cancer [163]. While these vaccines showcase varied cancer immunotherapy strategies, each has faced challenges in late-stage clinical evaluations. (Table 1)
Table 1. Below is a tabular list of various tumor vaccines in the last decade.
| Types of Tumor Vaccines | Strengths | Weaknesses | Examples | Mechanisms of Action | Effects | Limitations | References |
|---|---|---|---|---|---|---|---|
| Peptide vaccines |
|
|
Nelipepimut-S (NeuVax) | HER2-derived peptide vaccine | Activation of T-cell response | Limited overall survival improvement | [164] |
| CIMAvax-EGF | EGF-based peptide vaccine | Inhibition of EGF signaling | No direct tumor targeting | [101] | |||
| MUC1-based peptide vaccine | Targeting MUC1 tumor-associated antigens | Enhanced immune response | Heterogeneous patient response | [165] | |||
| DNA/RNA-based vaccines |
|
|
CV9104 (CureVac) | Uses mRNA to encode six antigens overexpressed in prostate cancer | Induced antigen-specific immune responses in early clinical trials | Efficacy in late-stage trials yet to be established; possibility of inducing autoimmune responses | [166] |
| Viral-vector-based vaccines |
|
|
Adenovirus-based vaccines (OncoVEXGM-CSF, CG0070) | Adenoviruses are modified to express a tumor-specific antigen or an immunomodulatory molecule; these stimulate an immune response against the tumor | Effective in stimulating an immune response against the tumor | Immune response to the viral vector can limit repeat dosing | [167] |
| Lentivirus-based vaccines (LV305) | Lentiviruses are engineered to deliver tumor-specific antigens to dendritic cells to stimulate a T-cell response | Successful in initiating T-cell responses | Safety concerns over integration into the host genome | [23] | |||
| Vaccinia-virus-based vaccines (JX-594) |
Vaccinia viruses are genetically engineered to express a tumor antigen and/or immunostimulatory molecule; they can directly lyse cancer cells | Showed antitumor activity and were well tolerated in clinical trials | Immune response to the viral vector can limit its effectiveness | [168] | |||
| Dendritic-cell-based vaccines |
|
|
Provenge (Sipuleucel-T) | The patient’s own dendritic cells are exposed to a fusion protein (prostatic acid phosphatase linked to an immune cell stimulating factor) | Extended overall survival in metastatic castration-resistant prostate cancer | Limited clinical benefits, high cost, and complex manufacturing process | [5] |
| DCVax-L | Autologous dendritic cells are pulsed with tumor lysate | Prolonged progression-free survival in glioblastoma multiforme (GBM) patients | Not FDA-approved; requires personalized manufacturing | [169] | |||
| Whole-cell-based vaccines |
|
|
GVAX | Utilizes autologous/allogeneic tumor cells that have been genetically modified to secrete the immune-stimulating cytokine GM-CSF | Demonstrated a significant immune response against cancer, studied in various types of cancer, including pancreatic and prostate cancers | Production can be labor-intensive and personalized; often requires co-administration with adjuvants or other immunomodulatory agents to enhance their immunogenicity | [157][170] |
| Canvaxin | Allogeneic melanoma cells mixed with Bacillus Calmette–Guérin (BCG) to stimulate immune response | Intended for melanoma treatment, but development discontinued due to insufficient effectiveness | Limited efficacy; potential for BCG-related side effects | [171] | |||
| Oncophage (Vitespen) | Uses heat shock proteins (gp96) derived from the patient’s tumor as an autologous vaccine | Showed efficacy in extending disease-free survival in certain patients with kidney cancer and melanoma | Not universally effective; personalized manufacturing can be labor-intensive | [172][173] |
The preceding table encapsulates seminal instances of assorted classifications of cancer vaccines, encompassing peptide-based, DNA/RNA-based, viral-vector-based, dendritic-cell-based, and whole-cell-based vaccines. Each paradigm is delineated in exhaustive detail, supplemented by pertinent bibliographical citations for subsequent scholarly inquiry.
The preceding table encapsulates seminal instances of assorted classifications of cancer vaccines, encompassing peptide-based, DNA/RNA-based, viral-vector-based, dendritic-cell-based, and whole-cell-based vaccines. Each paradigm is delineated in exhaustive detail, supplemented by pertinent bibliographical citations for subsequent scholarly inquiry.
4.6. Another Cancer Vaccine Therapy: In Situ Cancer Vaccines
In situ cancer vaccines represent a therapeutic approach where the tumor inside a patient’s body is directly targeted to serve as its own vaccine [61]. Rather than extracting tumor cells for external processing and reintroduction, in situ vaccines stimulate the immune system by damaging the tumor in its native environment [61]. As the tumor cells die, they release antigens, which are then recognized by the immune system. Often, this is achieved by injecting immune-stimulating agents or oncolytic viruses into the tumor [174][175]. This not only aims to destroy the immediate tumor but also primes the immune system to recognize and combat tumor cells elsewhere in the body [174]. In situ cancer vaccines have shown promise in preliminary studies.
4.7. Influencing Factors of Tumor Vaccines
Boosting the power of cancer vaccines is a top priority for researchers, who are diving deep into adjuvants and combination therapies to ramp up immune responses, outsmart tumor immune evasion, and prevent cancers from coming back [176][177]. The efficacy of cancer vaccines hinges on several factors, including picking the right antigens, choosing the adjuvants wisely, and using the best delivery systems [2]. Antigen selection is of paramount significance. The antigens should be specific to the tumor, or associated with it, so that the immune response zeroes in on cancer cells without harming healthy tissues [178]. Plus, the chosen antigens need to be highly immunogenic and able to stimulate both CD8+ cytotoxic T cells and CD4+ helper T cells for a strong, long-lasting attack against tumors [15]. Adjuvants help by making cancer vaccines more immunogenic. They stimulate the innate immune system, encourage antigen uptake by APCs, and help activate and expand antigen-specific T cells [179][180]. There are different types of adjuvants, like alum, toll-like receptor (TLR) agonists, and cytokines, each with unique mechanisms of action and varying effectiveness [181][182]. The latest research shows that TLR agonists, such as TLR9 and TLR7/8 agonists, have shown promise by bolstering antigen-presenting cells and intensifying immune responses [183]. Researchers have developed a nanosystem that can inhibit a process called MerTK-mediated efferocytosis. This inhibition leads to the release of immunogenic contents into the tumor microenvironment, potentially boosting the body’s natural defenses against the tumor [184]. Similarly, STING agonists enhance dendritic cell activity, boosting T-cell responses against tumors [185][186]. Oncolytic viruses, while serving as direct antitumor agents, also act as adjuvants by releasing tumor antigens within an inflammatory milieu [130]. These recent breakthroughs encapsulate the dynamic progression in adjuvant research, aiming to optimize the immune system’s potency against tumors.
5. Combination Therapies
Combining cancer vaccines with other therapies has emerged as a promising strategy to enhance the overall therapeutic efficacy and overcome the limitations of single-agent treatments [187]. Cancer vaccines, which aim to stimulate a patient’s immune system to recognize and attack tumor cells, may benefit from being combined with other immunotherapies, such as immune checkpoint inhibitors, to boost immune responses and counteract immunosuppressive mechanisms within the tumor microenvironment (TME) [188].
Several types of combination therapies involving cancer vaccines and other treatment modalities have been explored in recent years, such as combining cancer vaccines with chemotherapy, targeted therapies, and radiation therapy [187]. These combination approaches hold significant promise for optimizing cancer treatment outcomes and providing more effective, personalized therapy options for patients [72].
5.1. Cancer Vaccine + Immune Checkpoint Inhibitors
These vaccines can be combined with other treatments to enhance their effectiveness. Immune checkpoint inhibitors like pembrolizumab (Keytruda), nivolumab (Opdivo), and ipilimumab (Yervoy) disable immune checkpoints, thereby unleashing a more potent attack on cancer cells [189][190][191][192]. This combination hopes to enhance recognition of cancer cells (via the vaccine) and amplify the immune response (via the checkpoint inhibitors) [5][176].
This combined approach has shown promise in preclinical models and early clinical trials by generating tumor-specific T cells and preventing their exhaustion [193]. The mechanism behind these effects is that cancer vaccines aim to boost T cells’ recognition of tumor antigens, but this immune response can be dampened by the tumor’s evasion mechanisms [194]. Enter immune checkpoint inhibitors, which block inhibitory checkpoints (PD-1 and CTLA-4) on T cells, essentially “releasing the brakes” and amplifying their antitumor activity [195]. By combining cancer vaccines, which enhance the number of tumor-recognizing T cells, with checkpoint inhibitors that ensure that these T cells are not suppressed, there is a synergistic boost in the antitumor immune response. Preliminary studies suggest that this combination augments tumor attack, potentially leading to improved patient outcomes [196][197][198][199].
5.2. Cancer Vaccine + Chemotherapy
Chemotherapy is a destructive force against cancer cells, hindering their growth and division, but may also inadvertently harm rapidly dividing normal cells such as those in bone marrow, the digestive tract, and skin [200]. The potential synergy between cancer vaccines and chemotherapy arises from some chemotherapeutic agents inducing immunogenic cell death, increasing the visibility of dying cancer cells to the immune system and potentially enhancing the efficacy of cancer vaccines [201]. Several chemotherapeutic agents have been identified to potentially enhance the efficacy of cancer vaccines due to their immunomodulatory effects. For instance, cyclophosphamide and temozolomide can deplete immune-suppressing regulatory T cells (Tregs), creating a more receptive tumor environment for vaccine action [202]. Docetaxel, used for cancers like breast and prostate cancers, can bolster antigen presentation, thereby enhancing immune recognition of tumor cells [203]. Gemcitabine targets and reduces myeloid-derived suppressor cells (MDSCs) [204]. When combined with cancer vaccines, these agents can modify the tumor environment, diminish immune suppression, or amplify the immune response against tumors, although the choice of combination depends on multiple factors, including cancer type and patient health [201].
However, there are substantial challenges to this approach. Determining the optimal timing and dosage of chemotherapy in relation to cancer vaccines remains a complex task [157]. The side effects of both chemotherapy and cancer vaccines, including chemotherapy’s often severe systemic side effects such as fatigue, infection, hair loss, and nausea, are a significant concern [205]. Furthermore, the treatment’s responsiveness is limited, as not all cancer types respond well to chemotherapy or cancer vaccines, with variability in individual patient responses adding to the complexity of treatment plans [206][207]. Additionally, the complexity of the tumor microenvironment, which can evolve various mechanisms to resist or evade treatment, may limit the effectiveness of these combined therapies [208].
5.3. Cancer Vaccine + Radiotherapy
Radiotherapy employs high-energy particles or waves, such as X-rays, gamma rays, electron beams, or protons, to annihilate or damage cancer cells. This radiation induces small breaks in the DNA inside cells, inhibiting their growth and division, and eventually leading to their death [209]. When combined with tumor vaccines, these treatments might produce a synergistic effect, with radiotherapy potentially leading to the release of cancer cell antigens and stimulating the immune system, thereby enhancing the effectiveness of cancer vaccines [210][211][212]. Research indicates that radiotherapy exerts both cytotoxic and immunomodulatory effects on the tumor microenvironment. Beyond directly damaging tumor cells, RT induces immunogenic cell death, leading to the release of damage-associated molecular patterns (DAMPs) [213]. These DAMPs serve as “danger signals”, enhancing dendritic cell function and fostering antitumor immune responses. Concurrently, radiotherapy damages the tumor vasculature, increasing its permeability due to direct effects on endothelial cells and the upregulated release of VEGF from irradiated tumor cells [213][214]. This can lead to both transient improvements in oxygen and nutrient delivery and enhanced immune cell infiltration into the tumor.
However, this combination approach has its limitations. Not all patients or cancer types respond well to either radiotherapy or cancer vaccines, making the efficacy of this approach unclear in a broad population [215]. The optimal timing and dosage of radiotherapy relative to cancer vaccines are not well understood, posing a risk of radiotherapy killing immune cells stimulated by the vaccine, and thereby reducing the effectiveness of the treatment [216]. Both treatments can cause side effects, such as skin changes, fatigue, and other symptoms for radiotherapy, and usually mild but possibly flu-like symptoms for cancer vaccines [61], and some tumors may develop resistance to radiotherapy, which could limit the effectiveness of this combined approach [217].
5.4. Cancer Vaccine + Targeted Therapy
Compared to cancer vaccines, targeted therapies obstruct specific proteins or processes that aid in cancer growth and progression, offering a more cancer-cell-selective approach compared to traditional chemotherapy and resulting in fewer side effects. Notable targeted therapies include small-molecule inhibitors, like Gleevec (imatinib), and monoclonal antibodies, like Herceptin (trastuzumab) [218][219][220]. When utilized in combination, targeted therapies aim to inhibit cancer cells’ proliferation and survival, rendering the cancer cells more susceptible to the immune response provoked by the cancer vaccine. Studies have revealed the potential of this combination, with targeted therapies able to modulate the tumor microenvironment, thereby possibly enhancing the effectiveness of the vaccine-stimulated immune response [221] and helping to prevent or delay resistance to targeted therapies [2]. However, challenges and limitations remain, including the development of resistance to targeted therapies over time [222], potential side effects ranging from mild skin rashes or diarrhea to severe liver toxicity or heart problems [223], limited responsiveness in certain cancer types or patients [206], and the complex and not fully understood interaction effects between cancer vaccines and targeted therapies, which could potentially interfere with the vaccine-stimulated immune response [16].
5.5. Cancer Vaccine + Oncolytic Virotherapy
Oncolytic virotherapy constitutes a novel paradigm in the therapeutic approach towards malignant neoplasms, exhibiting a mechanism of action that distinguishes it from traditional tumor vaccines. It capitalizes on the unique capabilities of selected or genetically engineered viruses, which are orchestrated to specifically target and eradicate neoplastic cells [19][224]. Upon administration, these oncolytic viruses infiltrate the patient’s system, subjugating cancerous cells and commandeering their biological machinery for viral replication, consequently leading to cell lysis [225][226]. This lysogenic cycle not only facilitates direct oncolysis but also liberates tumor-specific antigens, providing a catalyst for the patient’s immune system to mount an anticancer response—an underpinning that is shared with the concept of tumor vaccines [130]. This dual-action mechanism that harmonizes direct cellular destruction with immune activation embodies a promising pathway in the realm of cancer therapy. The dual-action mechanism encompasses direct tumor cell lysis, releasing tumor-associated antigens, and the unveiling of damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs). These elements activate both the innate and adaptive arms of the immune system, enhancing antitumor responses. As newly assembled viral entities continue their onslaught against other malignant cells, a self-propagating cycle is established.
Recent advancements in this burgeoning field have entailed the exploration of diverse oncolytic virus models, such as the Parapoxvirus ovis model, known to induce an immunogenic form of cell death termed pyroptosis [227]. Scientific investigations have also scrutinized the immunostimulatory effects of these bioengineered viruses and combinational therapeutic strategies that kindle pyroptosis, consequently fostering potent antitumor activity [228][229][230]. These engineered viruses have some potential in the delivery of antitumor drugs [231]. Pioneering therapeutic strategies, such as the KISIMA/VSV-GP heterologous prime–boost methodology and the development of adenovirus-based tumor vaccines, have further emphasized the potential of oncolytic virotherapy as a formidable armament in the arsenal of cancer immunotherapy [232].
Cancer vaccines and oncolytic virotherapy offer a potential synergistic approach for cancer treatment. Cancer vaccines introduce cancer-specific antigens into the body, training the immune system to recognize and attack cells displaying these antigens [5]. Concurrently, oncolytic virotherapy uses engineered viruses that selectively infect and eliminate cancer cells, subsequently releasing tumor antigens and new viral particles that can infect nearby cancer cells, thereby stimulating an immune response [19]. In combination, the cancer vaccine’s potential enhancement of the immune response, coupled with the direct cellular damage from the oncolytic virus, could increase therapeutic effectiveness. Initial studies suggest that this combination can result in a more robust and long-lasting immune response against tumors, even potentially overcoming some immune evasion tactics employed by cancer cells [233]. Nevertheless, this approach is not without limitations. The immune system could respond to the oncolytic virus, reducing its cancer-killing effectiveness [234]. Additionally, delivering both oncolytic viruses and cancer vaccines to the tumor site, particularly in solid tumors, is challenging [235]. The diverse nature of cancers and variability in patient responses can limit the overall responsiveness of this combined therapy [236]. Finally, safety is a significant concern, as both oncolytic virotherapy and cancer vaccines can cause side effects, with the former potentially leading to severe or life-threatening reactions in rare instances [237]. Recent advancements will change this domain. Viruses are now engineered for heightened tumor specificity, some are armed with therapeutic genes to turn tumors into producers of anticancer agents, and combinations with treatments like immune checkpoint inhibitors are showing synergistic effects [238][239]. Moreover, refined genetic engineering techniques have improved the safety profiles of these oncolytic viruses, making them more amenable for therapeutic applications [240][241]. This means that they have reduced virulence in non-target tissues and minimized side effects.
6. Personalized Cancer Vaccines
Personalized cancer vaccine therapy is an innovative approach that tailors cancer treatment to a patient’s unique tumor profile. The process can be outlined as follows (Figure 4).
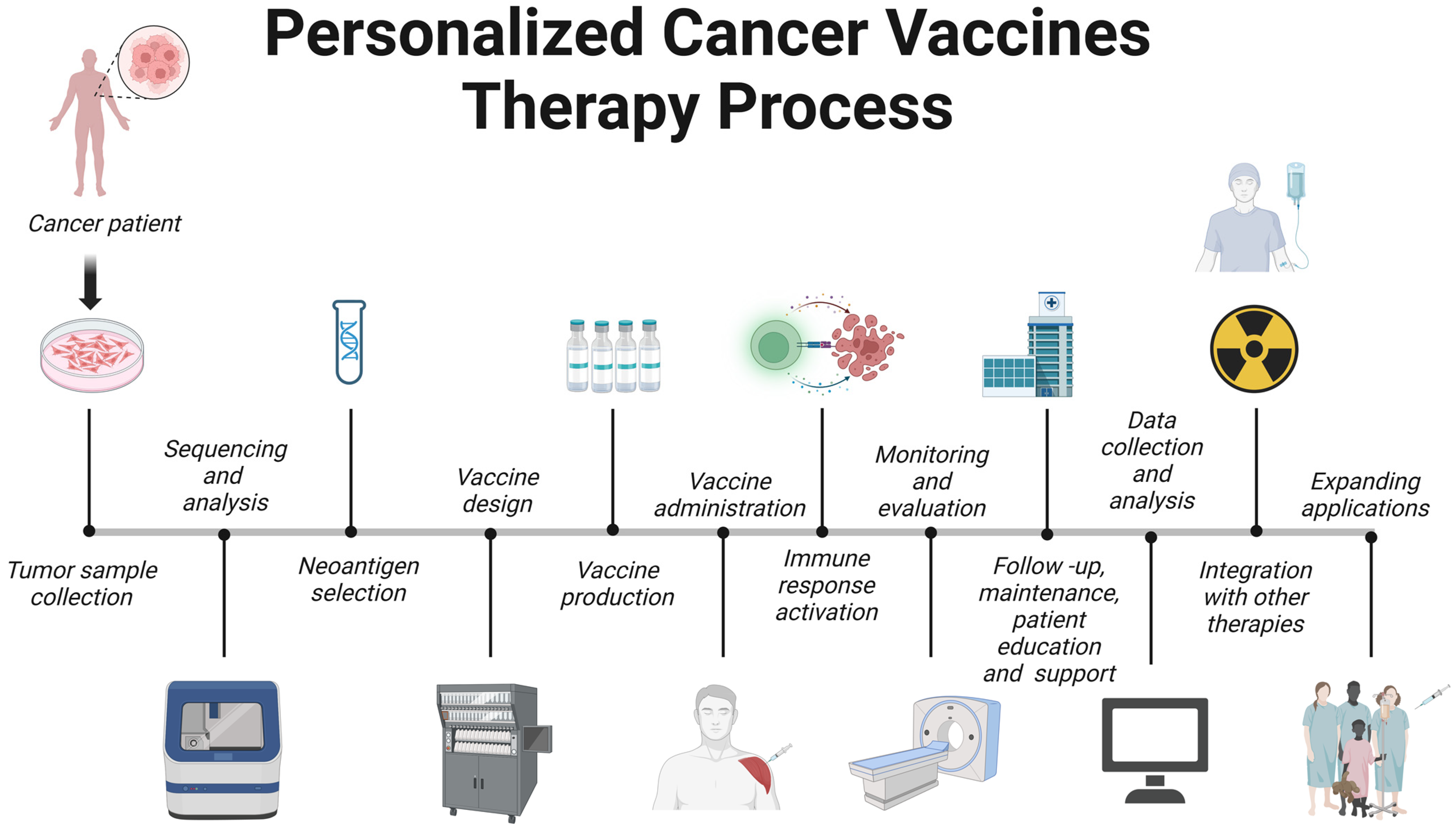
Figure 4. Therapeutic pipeline for personalized vaccines. Created with BioRender.com.
6.1. Tumor Sample Collection
A critical step in this process is the collection of tumor samples, which provide the essential genetic material needed to tailor the vaccine to the individual’s specific cancer [242]. The latest research advances have highlighted the importance of obtaining high-quality, well-preserved samples through minimally invasive techniques such as fine-needle aspiration or core-needle biopsy [243]. Additionally, there is a growing emphasis on collecting tumor samples at multiple timepoints throughout the course of treatment to account for the inherent heterogeneity of tumors and the potential for evolving cancer mutations [244]. The integration of these cutting-edge technologies and best practices in tumor sample collection is essential for maximizing the success of personalized cancer vaccine therapy [20].
6.2. Sequencing and Analysis
Sequencing and analysis empower researchers and clinicians to identify tumor-specific mutations and neoantigens that may function as potential therapeutic targets [20]. The development of next-generation sequencing (NGS) technologies, such as whole-exome and whole-genome sequencing, has greatly sped up the process and increased the precision of pinpointing tumor-specific mutations [175][245]. Concurrently, innovative computational approaches like machine learning algorithms and in silico prediction tools have emerged to forecast neoantigens with high immunogenicity, thereby expediting the selection of optimal vaccine candidates [242][246][247]. Moreover, incorporating multi-omics data, which include transcriptomics, proteomics, and epigenomics, offers a more comprehensive understanding of the tumor microenvironment and its influence on the effectiveness of personalized cancer vaccine therapy [248]. These advancements in sequencing and analysis have markedly improved our capacity to develop customized cancer vaccines, and they further emphasize the significance of multidisciplinary collaboration within the cancer immunotherapy field.
6.3. Neoantigen Selection
Neoantigen selection is vital for eliciting a robust and effective immune response [20][21]. Factors considered in the selection process include the binding affinity of the neoantigen to major histocompatibility complex (MHC) molecules, the immunogenicity of the epitope, and the likelihood of generating T-cell receptor (TCR) recognition [249][250]. Recent studies have also highlighted the importance of incorporating multi-omics data to ensure that the chosen neoantigens are effectively processed and presented on the cell surface [248]. By combining these innovative approaches, researchers have substantially improved the process of neoantigen selection, bolstering the potential for successful personalized cancer vaccine therapy.
6.4. Vaccine Design
Vaccine design directly influences the efficacy of the immune response against tumor cells [8]. Recent research advances have led to the development of various vaccine platforms, including peptide-based, nucleic-acid-based (DNA or RNA), viral-vector-based, and dendritic-cell-based vaccines, each with their own set of advantages and challenges [8][21][22]. The selection of appropriate adjuvants and delivery systems is essential for enhancing the immunogenicity of the vaccine and promoting the activation and expansion of tumor-specific T cells [251]. Innovative techniques such as liposomal and nanoparticle-based delivery systems have shown promise in improving vaccine stability, cellular uptake, and antigen presentation [252]. The swift pace of advancements in vaccine design methodologies, in conjunction with a burgeoning understanding of the tumor microenvironment and immune system, has notably augmented the potential of personalized cancer vaccine therapy, laying the groundwork for more effective and targeted cancer treatments.
6.5. Vaccine Production
The high quality of production in personalized cancer vaccine therapy is indispensable for ensuring the delivery of efficacious and safe treatment options to patients [22]. Noteworthy innovations in the production process encompass the incorporation of automation and process optimization to curtail the manufacturing duration and boost scalability [253]. Furthermore, the utilization of continuous manufacturing processes and the establishment of modular facilities have amplified flexibility and adaptability in vaccine production, thereby streamlining the supply chain for personalized therapies [254]. To ensure product quality, regulatory agencies have enforced good manufacturing practices (GMPs) and stringent quality control measures. The integration of advanced bioinformatics tools has also contributed to the acceleration of vaccine development and production, enabling more rapid clinical translation and patient access [108]. As the field of personalized cancer vaccine therapy continues to expand, further innovations in vaccine production technologies and processes will be vital to meeting the growing demand and ensuring the timely delivery of these tailored treatments.
6.6. Vaccine Administration
Vaccine administration directly impacts the induction of a robust immune response against tumor cells. The latest research advances have led to the exploration of various routes of administration, including subcutaneous, intradermal, intramuscular, and intranodal, with each route presenting unique benefits and challenges for different vaccine platforms [255][256]. The choice of administration route can influence the vaccine’s biodistribution, antigen presentation, and subsequent immune response [23]. Researchers are also investigating the optimal dosing and scheduling of these personalized vaccines to maximize their efficacy while minimizing potential adverse effects [21][257]. Recent studies have delved into the synergistic effects of merging personalized cancer vaccines with other immunotherapies to amplify therapeutic outcomes and surmount immune resistance [258].
As the realm of personalized cancer vaccine therapy continues to expand, a nuanced understanding of the optimal administration strategies, including route, dosage, and scheduling, will be pivotal for maximizing the therapeutic potential of these individualized treatments.
6.7. Immune Response Activation
The activation of the immune response is a central objective in personalized cancer vaccine therapy, aiming to instigate a robust and specific immune response against tumor cells expressing neoantigens. Recent research strides have provided a more comprehensive understanding of the mechanisms underpinning the activation of both innate and adaptive immune responses [20]. Personalized cancer vaccines endeavor to prime the immune system by presenting tumor-specific neoantigens to antigen-presenting cells (APCs) such as dendritic cells, which subsequently activate cytotoxic T lymphocytes (CTLs) to target and eliminate tumor cells [22][176][259]. A key facet of immune response activation lies in the optimization of vaccine design, thereby amplifying the immunogenicity of the vaccine and fostering the expansion of tumor-specific T cells [260].
Recent investigations have probed into combining personalized cancer vaccines with other immunotherapies to augment the antitumor immune response and counter the immune evasion strategies utilized by cancer cells [258]. As the realm of personalized cancer vaccine therapy progresses, a deeper comprehension of immune response activation and its modulation will be indispensable for maximizing the therapeutic potential of these tailored treatments.
6.8. Monitoring and Evaluation
Monitoring and evaluation are integral to ensuring the safety, efficacy, and optimization of these individualized treatments. Recent research breakthroughs have resulted in the development of comprehensive methodologies to assess both immune response and clinical outcomes in patients receiving personalized cancer vaccines [21][22]. Essential parameters for evaluating the immunological response include monitoring the expansion of vaccine-specific T cells, the production of cytokines, and the infiltration of immune cells into the tumor microenvironment [256]. These evaluations furnish invaluable insights into the vaccine’s capacity to activate and modulate the immune system.
Clinical evaluation entails tracking objective responses, such as reduction in tumor size, progression-free survival, and overall survival, while also considering the patients’ quality of life [23][261]. As personalized cancer vaccines are often administered in combination with other immunotherapies, it is crucial to identify synergistic effects and ascertain the optimal treatment regimen [258][262].
Moreover, monitoring and evaluating safety profiles is imperative for identifying and managing potential adverse effects associated with personalized cancer vaccines, such as autoimmune reactions or systemic inflammation [107]. The ongoing refinement of monitoring and evaluation strategies will contribute to maximizing the therapeutic potential and safety of personalized cancer vaccine therapies, thereby enhancing patient outcomes.
6.9. Follow-Up, Maintenance, and Patient Education and Support
Continuous follow-up empowers healthcare professionals to monitor patients’ responses to therapy, assess potential adverse effects, and make necessary adjustments to treatment plans [21][263]. Maintenance therapy, including administering booster vaccinations or adjusting combination therapies, is pivotal for maintaining the antitumor immune response and preventing cancer recurrence [22][72].
Patient education forms a key aspect of personalized cancer vaccine therapy, as it enables patients to make informed decisions about their treatment and manage potential side effects [255]. Patients should be well informed about potential adverse effects of the treatment and the importance of promptly reporting these to their healthcare providers [107][259]. Moreover, the emotional and psychological wellbeing of patients can significantly influence their response to treatment, underscoring the need for comprehensive support systems. This may include mental health counseling, peer support groups, and resources for family members, creating an environment that nurtures the patient’s resilience and determination [264]. As personalized cancer vaccine therapy continues to progress, it is crucial to maintain an integrated approach that addresses all facets of patient care, including follow-up, maintenance, education, and support. This holistic approach can maximize the therapeutic potential of personalized cancer vaccines and lead to improved patient outcomes [265].
6.10. Data Collection and Analysis
Data collection and analysis stand as the cornerstones of personalized cancer vaccine development and evaluation. Recent strides in high-throughput sequencing technologies, computational methodologies, and bioinformatics have ushered in a new era, enabling researchers to more precisely identify and prioritize neoantigens, craft personalized vaccines, and keep track of immune responses [20][22][256]. Furthermore, the application of machine learning algorithms and artificial intelligence has emerged as a powerful tool to navigate the complexity of these data landscapes. This technology bolsters our capacity to predict the immunogenicity of neoantigens and assess the effectiveness of personalized cancer vaccines [250]. Standardized data collection and analysis protocols are paramount to ensure reproducibility, facilitate comparisons across studies, and promote the development of robust and reliable personalized cancer vaccine therapies [23].
6.11. Integration with Other Therapies
The integration of personalized cancer vaccines with other therapeutic modalities offers an intriguing avenue in the realm of cancer treatment. Recent scientific findings highlight the potential synergistic effects of marrying personalized cancer vaccines with other forms of immunotherapies. Notably, ICIs, which target immunosuppressive pathways like PD-1 (programmed cell death protein 1), PD-L1 (programmed cell death ligand 1), and CTLA-4 (cytotoxic T-lymphocyte-associated protein 4), have shown promising results in combination with personalized cancer vaccines [107]. The rationale behind this combination is that the vaccine can stimulate a specific antitumor immune response, while ICIs can further enhance the function and persistence of tumor-specific T cells [189].
Additionally, personalized cancer vaccines can be combined with conventional therapies, such as chemotherapy and radiotherapy, to induce immunogenic cell death and release tumor-associated antigens, creating a more conducive environment for the activation of vaccine-induced immune responses [137]. Furthermore, combining personalized cancer vaccines with targeted therapies, such as kinase inhibitors or monoclonal antibodies, has shown promise in preclinical models by modulating the tumor microenvironment and improving immune cell infiltration [263][266].
6.12. Expanding Applications
The exploration of personalized cancer vaccine therapy applications continues to surge, unlocking new possibilities for cancer treatment across a diverse range of tumor types and stages. Groundbreaking studies have revealed the potential and effectiveness of tailored cancer vaccines against melanoma, glioblastoma, and non-small-cell lung cancer [22][261]. Aside from solid tumors, there is growing evidence of personalized cancer vaccines proving promising against hematological malignancies such as acute myeloid leukemia (AML) and multiple myeloma (MM) [267][268]. Furthermore, researchers are delving into the possibility of implementing personalized cancer vaccines during earlier disease stages, or as supplementary therapy after surgery or radiotherapy, with the ultimate goal of preventing relapse or disease progression [124].
As the development of personalized cancer vaccine therapy progresses, gaining a comprehensive understanding of the factors impacting vaccine effectiveness—including the tumor microenvironment, individual immune responses, and the dynamic interplay among various treatment approaches—becomes essential in extending its application scope. Unrelenting research and innovation are indispensable for harnessing the full therapeutic capacity of personalized cancer vaccines and enhancing outcomes for a wide spectrum of cancer patients. The world of clinical trials is abuzz with personalized cancer vaccines and neoantigen-targeting therapies [269][270]. Challenges exist, such as optimizing vaccine design and manufacturing, plus pinpointing the patients most likely to benefit from personalized immunotherapies [193][271]. Add in resistance mechanisms, sky-high costs, and complex production, and suddenly these groundbreaking treatments seem less accessible and affordable. So, the cancer immunotherapy research needs to zero in on resistance, refine neoantigen identification tech, and make personalized therapies more cost-effective and scalable [272]. This is where multi-omics approaches and AI come into play, helping to precisely stratify patients and craft combination therapies that boost efficacy while minimizing side effects [73]. Tackling these challenges and embracing emerging technologies will be the key to revolutionizing cancer treatment and offering new hope to countless patients.
This entry is adapted from the peer-reviewed paper 10.3390/ph16101384
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249.
- Mellman, I.; Coukos, G.; Dranoff, G. Cancer immunotherapy comes of age. Nature 2011, 480, 480–489.
- Wiemann, B.; Starnes, C.O. Coley’s toxins, tumor necrosis factor and cancer research: A historical perspective. Pharmacol. Ther. 1994, 64, 529–564.
- Burnet, M. Cancer-A Biological Approach: III Viruses Associated with Neoplastic Conditions. Br. Med. J. 1957, 1, 841.
- Kantoff, P.W.; Higano, C.S.; Shore, N.; Berger, E.; Small, E.J. Sipuleucel T immunotherapy for CRPC. N. Engl. J. 2010, 363, 411–422.
- Neelapu, S.S.; Locke, F.L.; Bartlett, N.L.; Lekakis, L.J.; Miklos, D.B.; Jacobson, C.A.; Braunschweig, I.; Oluwole, O.O.; Siddiqi, T.; Lin, Y.; et al. Axicabtagene Ciloleucel CAR T-Cell Therapy in Refractory Large B-Cell Lymphoma. N. Engl. J. Med. 2017, 377, 2531–2544.
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437.
- Sahin, U.; Türeci, Ö. Personalized vaccines for cancer immunotherapy. Science 2018, 359, 1355–1360.
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229.
- Saade, F.; Petrovsky, N. Technologies for enhanced efficacy of DNA vaccines. Expert Rev. Vaccines 2012, 11, 189–209.
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines-a new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279.
- Cecilia Larocca, B.S.; Schlom, J. Viral Vector Y Based Therapeutic Cancer Vaccines. Cancer J. 2011, 17, 359–371.
- Bubenik, J. Dendritic cell-based cancer vaccines. Folia Biol. 1999, 45, 71–74.
- De Gruijl, T.D.; Van Den Eertwegh, A.J.M.; Pinedo, H.M.; Scheper, R.J. Whole-cell cancer vaccination: From autologous to allogeneic tumor- and dendritic cell-based vaccines. Cancer Immunol. Immunother. 2008, 57, 1569–1577.
- Melero, I.; Gaudernack, G.; Gerritsen, W.; Huber, C.; Parmiani, G.; Scholl, S.; Thatcher, N.; Wagstaff, J.; Zielinski, C.; Faulkner, I.; et al. Therapeutic vaccines for cancer: An overview of clinical trials. Nat. Rev. Clin. Oncol. 2014, 11, 509–524.
- Galluzzi, L.; Buqué, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunological Effects of Conventional Chemotherapy and Targeted Anticancer Agents. Cancer Cell 2015, 28, 690–714.
- Sharabi, A.B.; Lim, M.; DeWeese, T.L.; Drake, C.G. Radiation and checkpoint blockade immunotherapy: Radiosensitisation and potential mechanisms of synergy. Lancet Oncol. 2015, 16, e498–e509.
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277.
- Russell, S.J.; Peng, K.W.; Bell, J.C. Oncolytic virotherapy. Nat. Biotechnol. 2012, 30, 658–670.
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science 2015, 348, 69–74.
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 2017, 547, 217–221.
- Keskin, D.B.; Anandappa, A.J.; Sun, J.; Tirosh, I.; Mathewson, N.D.; Li, S.; Oliveira, G.; Giobbie-Hurder, A.; Felt, K.; Gjini, E.; et al. Neoantigen vaccine generates intratumoral T cell responses in phase Ib glioblastoma trial. Nature 2019, 565, 234–239.
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016, 534, 396–401.
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674.
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596.
- Naci, D.; El Azreq, M.A.; Chetoui, N.; Lauden, L.; Sigaux, F.; Charron, D.; Al-Daccak, R.; Aoudjit, F. α2β1 integrin promotes chemoresistance against doxorubicin in cancer cells through extracellular signal-regulated kinase (ERK). J. Biol. Chem. 2012, 287, 17065–17076.
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252.
- Laczko, R.; Szauter, K.M.; Jansen, M.K.; Hollosi, P.; Muranyi, M.; Molnar, J.; Fong, K.S.K.; Hinek, A.; Csiszar, K. Active lysyl oxidase (LOX) correlates with focal adhesion kinase (FAK)/paxillin activation and migration in invasive astrocytes. Neuropathol. Appl. Neurobiol. 2007, 33, 631–643.
- Caon, I.; Bartolini, B.; Parnigoni, A.; Caravà, E.; Moretto, P.; Viola, M.; Karousou, E.; Vigetti, D.; Passi, A. Revisiting the hallmarks of cancer: The role of hyaluronan. Semin. Cancer Biol. 2019, 62, 9–16.
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183.
- Niland, S.; Riscanevo, A.X.; Eble, J.A. Matrix metalloproteinases shape the tumor microenvironment in cancer progression. Int. J. Mol. Sci. 2022, 23, 146.
- Lepucki, A.; Orlińska, K.; Mielczarek-Palacz, A.; Kabut, J.; Olczyk, P.; Komosińska-Vassev, K. The Role of Extracellular Matrix Proteins in Breast Cancer. J. Clin. Med. 2022, 11, 1250.
- Azimzade, Y.; Saberi, A.A.; Sahimi, M. Regulation of migration of chemotactic tumor cells by the spatial distribution of collagen fiber orientation. Phys. Rev. E 2019, 99, 062414.
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307.
- Multhoff, G.; Vaupel, P. Hypoxia compromises anti-cancer immune responses. Adv. Exp. Med. Biol. 2020, 1232, 131–143.
- Boedtkjer, E.; Pedersen, S.F. The Acidic Tumor Microenvironment as a Driver of Cancer. Annu. Rev. Physiol. 2020, 82, 103–126.
- Roman-Trufero, M.; Auner, H.W.; Edwards, C.M. Multiple myeloma metabolism—A treasure trove of therapeutic targets? Front. Immunol. 2022, 13, 897862.
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell. 2018, 33, 570–580.
- Li, W.; Deng, Y.; Chu, Q.; Zhang, P. Gut microbiome and cancer immunotherapy. Cancer Lett. 2019, 447, 41–47.
- Gajewski, T.F.; Schreiber, H.; Fu, Y.X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022.
- Fridman, W.H.; Pagès, F.; Saut̀s-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306.
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598.
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555.
- Koelwyn, G.J.; Quail, D.F.; Zhang, X.; White, R.M.; Jones, L.W. Exercise-dependent regulation of the tumour microenvironment. Nat. Rev. Cancer 2017, 17, 620–632.
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35.
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFβ in the tumour microenvironment. Nat. Rev. Cancer 2013, 13, 788–799.
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc Biol. 2004, 75, 163–189.
- Rosenberg, S.A. IL-2: The First Effective Immunotherapy for Human Cancer. J. Immunol. 2014, 192, 5451–5458.
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146.
- Kim, H.J.; Cantor, H. CD4 T-cell subsets and tumor immunity: The helpful and the not-so-helpful. Cancer Immunol. Res. 2014, 2, 91–98.
- David, C.J.; Massagué, J. Contextual determinants of TGFβ action in development, immunity and cancer. Nat. Rev. Mol. Cell Biol. 2018, 19, 419–435.
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248.
- De Streel, G.; Lucas, S. Targeting immunosuppression by TGF-β1 for cancer immunotherapy. Biochem. Pharmacol. 2021, 192, 114697.
- Cattin, S.; Fellay, B.; Pradervand, S.; Trojan, A.; Ruhstaller, T.; Rüegg, C.; Fürstenberger, G. Bevacizumab specifically decreases elevated levels of circulating KIT+CD11b+ cells and IL-10 in metastatic breast cancer patients. Oncotarget 2016, 7, 11137–11150.
- Bolandi, S.M.; Abdolmaleki, Z.; Assarehzadegan, M.A. Bevacizumab regulates inflammatory cytokines and inhibits VEGFR2 signaling pathway in an ovalbumin-induced rat model of airway hypersensitivity. Inflammopharmacology 2021, 29, 683–694.
- Arenas-Ramirez, N.; Woytschak, J.; Boyman, O. Interleukin-2: Biology, Design and Application. Trends. Immunol. 2015, 36, 763–777.
- Prochazkova, J.; Pokorna, K.; Holan, V. IL-12 inhibits the TGF-β-dependent T cell developmental programs and skews the TGF-β-induced differentiation into a Th1-like direction. Immunobiology 2012, 217, 74–82.
- Kwok, G.; Yau, T.C.C.; Chiu, J.W.; Tse, E.; Kwong, Y.L. Pembrolizumab (Keytruda). Hum. Vaccines Immunother. 2016, 12, 2777–2789.
- Lei, X.; Lei, Y.; Li, J.-K.; Du, W.-X.; Li, R.-G.; Yang, J.; Li, J.; Li, F.; Tan, H.-B. Immune cells within the tumor microenvironment: Biological functions and roles in cancer immunotherapy. Cancer Lett. 2020, 470, 126–133.
- Jou, J.; Harrington, K.J.; Zocca, M.B.; Ehrnrooth, E.; Cohen, E.E.W. The changing landscape of therapeutic cancer vaccines-novel platforms and neoantigen identification. Clin. Cancer Res. 2021, 27, 689–703.
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic cancer vaccines. Nat. Rev. Cancer 2021, 21, 360–378.
- Thomas, S.; Prendergast, G.C. Cancer vaccines: A brief overview. Methods Mol. Biol. 2016, 1403, 755–761.
- Jiang, W.; He, Y.; He, W.; Wu, G.; Zhou, X.; Sheng, Q.; Zhong, W.; Lu, Y.; Ding, Y.; Lu, Q.; et al. Exhausted CD8+ T Cells in the Tumor Immune Microenvironment: New Pathways to Therapy. Front. Immunol. 2021, 11, 622509.
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4+ T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148.
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From vaccines to memory and back. Immunity 2010, 33, 451–463.
- Irvine, D.J.; Swartz, M.A.; Szeto, G.L. Engineering synthetic vaccines using cues from natural immunity. Nat. Mater. 2013, 12, 978–990.
- Pulendran, B.; Ahmed, R. Immunological mechanisms of vaccination. Nat. Immunol. 2011, 12, 509–517.
- Weiner, L.M.; Surana, R.; Wang, S. Monoclonal antibodies: Versatile platforms for cancer immunotherapy. Nat. Rev. Immunol. 2010, 10, 317–327.
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer immunoediting: From immunosurveillance to tumor escape. Nat. Immunol. 2002, 3, 991–998.
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases-elimination, equilibrium and escape. Curr. Opin. Immunol. 2014, 27, 16–25.
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.M.C.S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198.
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355.
- Chen, D.S.; Mellman, I. Elements of cancer immunity and the cancer-immune set point. Nature 2017, 541, 321–330.
- Galon, J.; Bruni, D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat. Rev. Drug Discov. 2019, 18, 197–218.
- Galon, J.; Mlecnik, B.; Bindea, G.; Angell, H.K.; Berger, A.; Lagorce, C.; Lugli, A.; Zlobec, I.; Hartmann, A.; Bifulco, C.; et al. Towards the introduction of the “Immunoscore” in the classification of malignant tumours. J. Pathol. 2014, 232, 199–209.
- Topalian, S.L.; Taube, J.M.; Pardoll, D.M. Neoadjuvant checkpoint blockade for cancer immunotherapy. Science 2020, 367, 6477.
- Finn, O.J. Cancer vaccines: Between the idea and the reality. Nat. Rev. Immunol. 2003, 3, 630–641.
- Cella, M.; Engering, A.; Pinet, V.; Pieters, J.; Lanzavecchia, A. Inflammatory stimuli induce accumulation of MHC class II complexes on dendritic cells. Nature 1997, 388, 782–787.
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242.
- Rosenberg, S.A.; Yang, J.C.; Restifo, N.P. Cancer immunotherapy: Moving beyond current vaccines. Nat. Med. 2004, 10, 909–915.
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Rev. Cancer 2012, 12, 278–287.
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570.
- Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125 (Suppl. 2), S41–S52.
- Golay, J.; Introna, M. Mechanism of action of therapeutic monoclonal antibodies: Promises and pitfalls of in vitro and in vivo assays. Arch. Biochem. Biophys. 2012, 526, 146–153.
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47.
- Orange, J.S. Formation and function of the lytic NK-cell immunological synapse. Nat. Rev. Immunol. 2008, 8, 713–725.
- Bryceson, Y.T.; March, M.E.; Ljunggren, H.G.; Long, E.O. Activation, coactivation, and costimulation of resting human natural killer cells. Immunol. Rev. 2006, 214, 73–91.
- Greenberg, S.; Grinstein, S. Phagocytosis and innate immunity Greenberg and Grinstein 137. Sci. Rep. 2002, 14, 136–145.
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50.
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797.
- Morgan, B.P. The membrane attack complex as an inflammatory trigger. Immunobiology 2016, 221, 747–751.
- Schumacher, T.; Bunse, L.; Pusch, S.; Sahm, F.; Wiestler, B.; Quandt, J.; Menn, O.; Osswald, M.; Oezen, I.; Ott, M.; et al. A vaccine targeting mutant IDH1 induces antitumour immunity. Nature 2014, 512, 324–327.
- Coulie, P.G.; Van Den Eynde, B.J.; Van Der Bruggen, P.; Boon, T. Tumour antigens recognized by T lymphocytes: At the core of cancer immunotherapy. Nat. Rev. Cancer 2014, 14, 135–146.
- Melief, C.J.M.; Van Hall, T.; Arens, R.; Ossendorp, F.; Van Der Burg, S.H. Therapeutic cancer vaccines. J. Clin. Investig. 2015, 125, 3401–3412.
- Melief, C.J.M.; Van der Burg, S.H.; Toes, R.E.M.; Ossendorp, F.; Offringa, R. Effective therapeutic anticancer vaccines based on precision guiding of cytolytic T lymphocytes. Immunol. Rev. 2002, 188, 177–182.
- Jiang, S.; Gong, M.; Xu, X.N. Editorial: Research, Development and Clinical Trials for Peptide-Based Vaccines. Front. Immunol. 2022, 13, 894989.
- Schneble, E.J.; Berry, J.S.; Trappey, F.A.; Clifton, G.T.; Ponniah, S.; Mittendorf, E.; Peoples, G.E. The HER2 peptide nelipepimut-S (E75) vaccine (NeuVaxTM) in breast cancer patients at risk for recurrence: Correlation of immunologic data with clinical response. Immunotherapy 2014, 6, 519–531.
- Chick, R.C.; Clifton, G.T.; Hale, D.F.; Vreeland, T.J.; Hickerson, A.T.; Bohan, P.M.K.; McCarthy, P.M.; Litton, J.K.; Alatrash, G.; Murthy, R.K.; et al. Subgroup analysis of nelipepimut-S plus GM-CSF combined with trastuzumab versus trastuzumab alone to prevent recurrences in patients with high-risk, HER2 low-expressing breast cancer. Clin. Immunol. 2021, 225, 108679.
- Clifton, G.T.; Peoples, G.E.; Mittendorf, E.A. The development and use of the E75 (HER2 369-377) peptide vaccine. Future Oncol. 2016, 12, 1321–1329.
- Mittendorf, E.A.; Lu, B.; Melisko, M.; Hiller, J.P.; Bondarenko, I.; Brunt, A.M.; Sergii, G.; Petrakova, K.; Peoples, G.E. Efficacy and safety analysis of nelipepimut-S vaccine to prevent breast cancer recurrence: A randomized, multicenter, phase III clinical trial. Clin. Cancer Res. 2019, 25, 4248–4254.
- Rodriguez, P.C.; Popa, X.; Martínez, O.; Mendoza, S.; Santiesteban, E.; Crespo, T.; Amador, R.M.; Fleytas, R.; Acosta, S.C.; Otero, Y.; et al. A Phase III Clinical Trial of the Epidermal Growth Factor Vaccine CIMAvax-EGF as Switch Maintenance Therapy in Advanced Non-Small Cell Lung Cancer Patients. Clin. Cancer Res. 2016, 22, 3782–3790.
- Tagliamento, M.; Rijavec, E.; Barletta, G.; Biello, F.; Rossi, G.; Grossi, F.; Genova, C. CIMAvax-EGF, a therapeutic non-small cell lung cancer vaccine. Expert Opin. Biol. Ther. 2018, 18, 829–835.
- Hanisch, F.G. Design of a MUC1-based cancer vaccine. Biochem. Soc. Trans. 2005, 33, 705–708.
- Gao, T.; Cen, Q.; Lei, H. A review on development of MUC1-based cancer vaccine. Biomed. Pharmacother. 2020, 132, 110888.
- Taylor-Papadimitriou, J.; Burchell, J.M.; Graham, R.; Beatson, R. Latest developments in MUC1 immunotherapy. Biochem. Soc. Trans. 2018, 46, 659–668.
- Zahm, C.D.; Colluru, V.T.; McNeel, D.G. DNA vaccines for prostate cancer. Pharmacol. Ther. 2017, 174, 27–42.
- Sahin, U.; Derhovanessian, E.; Miller, M.; Kloke, B.-P.; Simon, P.; Löwer, M.; Bukur, V.; Tadmor, A.D.; Luxemburger, U.; Schrörs, B.; et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 2017, 547, 222–226.
- Kreiter, S.; Vormehr, M.; van de Roemer, N.; Diken, M.; Löwer, M.; Diekmann, J.; Boegel, S.; Schrörs, B.; Vascotto, F.; Castle, J.C.; et al. Mutant MHC class II epitopes drive therapeutic immune responses to cancer. Nature 2015, 520, 692–696.
- Mei, Y.; Wang, X. RNA modification in mRNA cancer vaccines. Clin. Exp. Med. 2023, 14, 1–15.
- Kuai, R.; Ochyl, L.J.; Bahjat, K.S.; Schwendeman, A.; Moon, J.J. Designer vaccine nanodiscs for personalized cancer immunotherapy. Nat. Mater. 2017, 16, 489–498.
- Su, C.; Peng, L.; Sham, J.; Wang, X.; Zhang, Q.; Chua, D.; Liu, C.; Cui, Z.; Xue, H.; Wu, H.; et al. Immune Gene-Viral Therapy with Triplex Efficacy Mediated by Oncolytic Adenovirus Carrying an Interferon-γ Gene Yields Efficient Antitumor Activity in Immunodeficient and Immunocompetent Mice. Mol. Ther. 2006, 13, 918–927.
- Liu, M.A. Transgene FDNA vaccines: A review. J. Intern. Med. 2003, 253, 402–410.
- Blass, E.; Ott, P.A. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat. Rev. Clin. Oncol. 2021, 18, 215–229.
- Wang, Y.; Zhang, L.; Xu, Z.; Miao, L.; Huang, L. mRNA Vaccine with Antigen-Specific Checkpoint Blockade Induces an Enhanced Immune Response against Established Melanoma. Mol. Ther. 2018, 26, 420–434.
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586.
- Miao, L.; Zhang, Y.; Huang, L. mRNA vaccine for cancer immunotherapy. Mol. Cancer 2021, 20, 41.
- Cu, Y.; Broderick, K.E.; Banerjee, K.; Hickman, J.; Otten, G.; Barnett, S.; Kichaev, G.; Sardesai, N.Y.; Ulmer, J.B.; Geall, A. Enhanced Delivery and Potency of Self-Amplifying mRNA Vaccines by Electroporation in Situ. Vaccines 2013, 1, 367–383.
- Ulmer, J.B.; Geall, A.J. Recent innovations in mRNA vaccines. Curr. Opin. Immunol. 2016, 41, 18–22.
- Ho, W.; Gao, M.; Li, F.; Li, Z.; Zhang, X.Q.; Xu, X. Next-Generation Vaccines: Nanoparticle-Mediated DNA and mRNA Delivery. Adv. Healthc. Mater. 2021, 10, 2001812.
- Huang, X.; Zhang, G.; Tang, T.Y.; Gao, X.; Liang, T.B. Personalized pancreatic cancer therapy: From the perspective of mRNA vaccine. Mil. Med. Res. 2022, 9, 53.
- Brisse, M.; Vrba, S.M.; Kirk, N.; Liang, Y.; Ly, H. Emerging Concepts and Technologies in Vaccine Development. Front. Immunol. 2020, 11, 583077.
- Rausch, S.; Schwentner, C.; Stenzl, A.; Bedke, J. mRNA vaccine CV9103 and CV9104 for the treatment of prostate cancer. Hum. Vaccines Immunother. 2014, 10, 3146–3152.
- Rosenberg, S.A.; Restifo, N.P. Adoptive cell transfer as personalized immunotherapy for human cancer. Science 2015, 348, 62–68.
- Finn, O.J. The dawn of vaccines for cancer prevention. Nat. Rev. Immunol. 2018, 18, 183–194.
- Lichty, B.D.; Breitbach, C.J.; Stojdl, D.F.; Bell, J.C. Going viral with cancer immunotherapy. Nat. Rev. Cancer 2014, 14, 559–567.
- O’Bryan, S.M.; Mathis, J.M. Oncolytic Virotherapy for Breast Cancer Treatment. Curr. Gene Ther. 2018, 18, 192–205.
- Drake, C.G. Prostate cancer as a model for tumour immunotherapy. Nat. Rev. Immunol. 2010, 10, 580–593.
- Gubin, M.M.; Artyomov, M.N.; Mardis, E.R.; Schreiber, R.D. Tumor neoantigens: Building a framework for personalized cancer immunotherapy. J. Clin. Investig. 2015, 125, 3413–3421.
- Russell, S.J.; Peng, K.W. Viruses as anticancer drugs. Trends Pharmacol. Sci. 2007, 28, 326–333.
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic viruses: A new class of immunotherapy drugs. Nat. Rev. Drug Discov. 2015, 14, 642–662.
- Rollier, C.S.; Reyes-Sandoval, A.; Cottingham, M.G.; Ewer, K.; Hill, A.V.S. Viral vectors as vaccine platforms: Deployment in sight. Curr. Opin. Immunol. 2011, 23, 377–382.
- Ura, T.; Okuda, K.; Shimada, M. Developments in viral vector-based vaccines. Vaccines 2014, 2, 624–641.
- Pol, J.; Bloy, N.; Obrist, F.; Eggermont, A.; Galon, J.; Cremer, I.; Erbs, P.; Limacher, J.M.; Preville, X.; Zitvogel, L.; et al. Trial watch: Oncolytic viruses for cancer therapy. Oncoimmunology 2014, 3, 37–41.
- Prestwich, R.J.D.; Harrington, K.J.; Pandha, H.S.; Vile, R.G.; Melcher, A.A.; Errington, F. Oncolytic viruses: A novel form of immunotherapy. Expert Rev. Anticancer Ther. 2008, 8, 1581–1588.
- Anderson, R.J.; Schneider, J. Plasmid DNA and viral vector-based vaccines for the treatment of cancer. Vaccine 2007, 25 (Suppl. 2), 24–34.
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer 2012, 12, 237–251.
- Galluzzi, L.; Buqué, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic cell death in cancer and infectious disease. Nat. Rev. Immunol. 2017, 17, 97–111.
- Watanabe, D.; Goshima, F. Oncolytic virotherapy by HSV. Adv. Exp. Med. Biol. 2018, 1045, 63–84.
- Ramesh, N.; Ge, Y.; Ennist, D.L.; Zhu, M.; Mina, M.; Ganesh, S.; Reddy, P.S.; Yu, D.C. CG0070, a conditionally replicating granulocyte macrophage colony-stimulating factor—Armed oncolytic adenovirus for the treatment of bladder cancer. Clin. Cancer Res. 2006, 12, 305–313.
- Albershardt, T.C.; Campbell, D.J.; Parsons, A.J.; Slough, M.M.; Ter Meulen, J.; Berglund, P. LV305, a dendritic cell-targeting integration-deficient ZVex TM-based lentiviral vector encoding NY-ESO-1, induces potent anti-tumor immune response. Mol. Ther. Oncolytics 2016, 3, 305–313.
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia JX-594 in liver cancer. Nat. Med. 2013, 19, 329–336.
- Breitbach, C.; Bell, J.C.; Hwang, T.-H.; Kirn, D.; Burke, J. The emerging therapeutic potential of the oncolytic immunotherapeutic Pexa-Vec (JX-594). Oncolytic Virother. 2015, 4, 25–31.
- Parato, K.A.; Breitbach, C.J.; Le Boeuf, F.; Wang, J.; Storbeck, C.; Ilkow, C.; Diallo, J.-S.; Falls, T.; Burns, J.; Garcia, V.; et al. The oncolytic poxvirus JX-594 selectively replicates in and destroys cancer cells driven by genetic pathways commonly activated in cancers. Mol. Ther. 2012, 20, 749–758.
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252.
- Rosenberg, S.A.; Yang, J.C.; Schwartzentruber, D.J.; Hwu, P.; Marincola, F.M.; Topalian, S.L.; Restifo, N.P.; Sznol, M.; Schwarz, S.L.; Spiess, P.J.; et al. Impact of Cytokine Administration on the Generation of Antitumor Reactivity in Patients with Metastatic Melanoma Receiving a Peptide Vaccine. J. Immunol. 1999, 163, 1690–1695.
- Tuyaerts, S.; Aerts, J.L.; Corthals, J.; Neyns, B.; Heirman, C.; Breckpot, K.; Thielemans, K.; Bonehill, A. Current approaches in dendritic cell generation and future implications for cancer immunotherapy. Cancer Immunol. Immunother. 2007, 56, 1513–1537.
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24.
- Han, J.; Bhatta, R.; Liu, Y.; Bo, Y.; Elosegui-Artola, A.; Wang, H. Metabolic glycan labeling immobilizes dendritic cell membrane and enhances antitumor efficacy of dendritic cell vaccine. Nat. Commun. 2023, 14, 5049.
- Carreno, B.M.; Magrini, V.; Becker-Hapak, M.; Kaabinejadian, S.; Hundal, J.; Petti, A.A.; Ly, A.; Lie, W.R.; Hildebrand, W.H.; Mardis, E.R.; et al. A dendritic cell vaccine increases the breadth and diversity of melanoma neoantigen-specific T cells. Science 2015, 348, 803–808.
- Morse, M.A.; Clay, T.M.; Hobeika, A.C.; Osada, T.; Khan, S.; Chui, S.; Niedzwiecki, D.; Panicali, D.; Schlom, J.; Lyerly, H.K. Phase I study of immunization with dendritic cells modified with fowlpox encoding carcinoembryonic antigen and costimulatory molecules. Clin. Cancer Res. 2005, 11, 3017–3024.
- Anguille, S.; Smits, E.L.; Lion, E.; Van Tendeloo, V.F.; Berneman, Z.N. Clinical use of dendritic cells for cancer therapy. Lancet Oncol. 2014, 15, 257–267.
- Antonarakis, E.S.; Small, E.J.; Petrylak, D.P.; Quinn, D.I.; Kibel, A.S.; Chang, N.N.; Dearstyne, E.; Harmon, M.; Campogan, D.; Haynes, H.; et al. Antigen-specific CD8 lytic phenotype induced by sipuleucel-T in hormone-sensitive or castration-resistant prostate cancer and association with overall survival. Clin. Cancer Res. 2018, 24, 4662–4671.
- Carballido, E.; Fishman, M. Sipuleucel-T: Prototype for development of anti-tumor vaccines. Curr. Oncol. Rep. 2011, 13, 112–119.
- Polyzoidis, S.; Ashkan, K. DCVax®-L—Developed by Northwest Biotherapeutics. Hum. Vaccines Immunother. 2014, 10, 3139–3145.
- Guo, C.; Manjili, M.H.; Subjeck, J.R.; Sarkar, D.; Fisher, P.B.; Wang, X.Y. Therapeutic cancer vaccines. In Past, Present, and Future, 1st ed.; Advances in Cancer Research; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 119, pp. 421–475.
- Shu, S.; Zheng, R.; Lee, W.T.; Cohen, P.A. Immunogenicity of dendritic-tumor fusion hybrids and their utility in cancer immunotherapy. Crit. Rev. Immunol. 2007, 27, 463–483.
- Emens, L.A.; Jaffee, E.M. Leveraging the activity of tumor vaccines with cytotoxic chemotherapy. Cancer Res. 2005, 65, 8059–8064.
- Principe, D.R.; Korc, M.; Kamath, S.D.; Munshi, H.G.; Rana, A. Trials and tribulations of pancreatic cancer immunotherapy. Cancer Lett. 2021, 504, 1–14.
- Ward, J.E.; McNeel, D.G. GVAX: An allogeneic, whole-cell, GM-CSF-secreting cellular immunotherapy for the treatment of prostate cancer. Expert Opin. Biol. Ther. 2007, 7, 1893–1902.
- Petricciani, J.; Koren, E.; Morton, D. Analysis of the in vivo proliferative capacity of a whole cell cancer vaccine. Biologicals 2016, 44, 60–63.
- Di Pietro, A.; Tosti, G.; Ferrucci, P.F.; Testori, A. Oncophage: Step to the future for vaccine therapy in melanoma. Expert Opin. Biol. Ther. 2008, 8, 1973–1984.
- Jacobsohn, K.M.; Wood, C.G. Adjuvant Therapy for Renal Cell Carcinoma. Semin. Oncol. 2006, 33, 576–582.
- Itoh, K.; Yamada, A.; Mine, T.; Noguchi, M. Recent advances in cancer vaccines: An overview. Jpn. J. Clin. Oncol. 2009, 39, 73–80.
- Mittendorf, E.A.; Ardavanis, A.; Litton, J.K.; Shumway, N.M.; Hale, D.F.; Murray, J.L.; Perez, S.A.; Ponniah, S.; Baxevanis, C.N.; Papamichail, M.; et al. Primary analysis of a prospective, randomized, single-blinded phase II trial evaluating the HER2 peptide GP2 vaccine in breast cancer patients to prevent recurrence. Oncotarget 2016, 7, 66192–66201.
- Lepisto, A.J.; Moser, A.J.; Zeh, H.; Lee, K.; Bartlett, D.; McKolanis, J.R.; Geller, B.A.; Schmotzer, A.; Potter, D.P.; Whiteside, T.; et al. A phase I/II study of a MUC1 peptide pulsed autologous dendritic cell vaccine as adjuvant therapy in patients with resected pancreatic and biliary tumors. Cancer Ther. 2008, 6, 955–964.
- Kübler, H.; Scheel, B.; Gnad-Vogt, U.; Miller, K.; Schultze-Seemann, W.; Dorp, F.V.; Parmiani, G.; Hampel, C.; Wedel, S.; Trojan, L.; et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: A first-in-man phase I/IIa study. J. Immunother. Cancer 2015, 3, 26.
- Kaufman, H.L.; Kim, D.W.; Deraffele, G.; Mitcham, J.; Coffin, R.S.; Kim-Schulze, S. Local and distant immunity induced by intralesional vaccination with an oncolytic herpes virus encoding GM-CSF in patients with stage IIIc and IV melanoma. Ann. Surg. Oncol. 2010, 17, 718–730.
- Hwang, T.-H.; Moon, A.; Burke, J.; Ribas, A.; Stephenson, J.; Breitbach, C.J.; Daneshmand, M.; De Silva, N.; Parato, K.; Diallo, J.-S.; et al. A mechanistic proof-of-concept clinical trial with JX-594, a targeted multi-mechanistic oncolytic poxvirus, in patients with metastatic melanoma. Mol. Ther. 2011, 19, 1913–1922.
- Liau, L.M.; Ashkan, K.; Tran, D.D.; Campian, J.L.; Trusheim, J.E.; Cobbs, C.S.; Heth, J.A.; Salacz, M.; Taylor, S.; D’andre, S.D.; et al. First results on survival from a large Phase 3 clinical trial of an autologous dendritic cell vaccine in newly diagnosed glioblastoma. J. Transl. Med. 2018, 16, 142.
- Lutz, E.; Yeo, C.J.; Lillemoe, K.D.; Biedrzycki, B.; Kobrin, B.; Herman, J.; Sugar, E.; Piantadosi, S.; Cameron, J.L.; Solt, S.; et al. A lethally irradiated allogeneic granulocyte-macrophage colony stimulating factor-secreting tumor vaccine for pancreatic adenocarcinoma: A phase II trial of safety, efficacy, and immune activation. Ann. Surg. 2011, 253, 328–335.
- Morton, D.L.; Hsueh, E.C.; Essner, R.; Foshag, L.J.; O’Day, S.J.; Bilchik, A.; Gupta, R.K.; Hoon, D.S.; Ravindranath, M.; Nizze, J.A.; et al. Prolonged survival of patients receiving active immunotherapy with Canvaxin therapeutic polyvalent vaccine after complete resection of melanoma metastatic to regional lymph nodes. Ann. Surg. 2002, 236, 438–449.
- Wood, C.; Srivastava, P.; Bukowski, R.; Lacombe, L.; Gorelov, A.I.; Gorelov, S.; Mulders, P.; Zielinski, H.; Hoos, A.; Teofilovici, F.; et al. An adjuvant autologous therapeutic vaccine (HSPPC-96; vitespen) versus observation alone for patients at high risk of recurrence after nephrectomy for renal cell carcinoma: A multicentre, open-label, randomised phase III trial. Lancet 2008, 372, 145–154.
- Testori, A.; Richards, J.; Whitman, E.; Mann, G.B.; Lutzky, J.; Camacho, L.; Parmiani, G.; Tosti, G.; Kirkwood, J.M.; Hoos, A.; et al. Phase III comparison of vitespen, an autologous tumor-derived heat shock protein gp96 peptide complex vaccine, with physician’s choice of treatment for stage IV melanoma: The C-100-21 study group. J. Clin. Oncol. 2008, 26, 955–962.
- Lawler, S.E.; Speranza, M.C.; Cho, C.F.; Chiocca, E.A. Oncolytic viruses in cancer treatment a review. JAMA Oncol. 2017, 3, 841–849.
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer vaccines: The next immunotherapy frontier. Nat. Cancer 2022, 3, 911–926.
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264.
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune checkpoint blockade: A common denominator approach to cancer therapy. Cancer Cell 2015, 27, 450–461.
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The prioritization of cancer antigens: A National Cancer Institute pilot project for the acceleration of translational research. Clin. Cancer Res. 2009, 15, 5323–5337.
- O’Hagan, D.T.; Fox, C.B. New generation adjuvants—From empiricism to rational design. Vaccine 2015, 33, B14–B20.
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650.
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503.
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608.
- Kocic, G.; Hadzi-Djokic, J.; Colic, M.; Veljkovic, A.; Tomovic, K.; Roumeliotis, S.; Smelcerovic, A.; Liakopoulos, V. The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue. Int. J. Mol. Sci. 2023, 24, 509.
- Zhuang, W.-R.; Wang, Y.; Nie, W.; Lei, Y.; Liang, C.; He, J.; Zuo, L.; Huang, L.-L.; Xie, H.-Y. Bacterial outer membrane vesicle based versatile nanosystem boosts the efferocytosis blockade triggered tumor-specific immunity. Nat. Commun. 2023, 14, 1675.
- Garland, K.M.; Sheehy, T.L.; Wilson, J.T. Chemical and Biomolecular Strategies for STING Pathway Activation in Cancer Immunotherapy. Chem. Rev. 2022, 122, 5977–6039.
- Li, A.; Yi, M.; Qin, S.; Song, Y.; Chu, Q.; Wu, K. Activating cGAS-STING pathway for the optimal effect of cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 35.
- Hegde, P.S.; Chen, D.S. Top 10 Challenges in Cancer Immunotherapy. Immunity 2020, 52, 17–35.
- Patel, S.A.; Minn, A.J. Combination Cancer Therapy with Immune Checkpoint Blockade: Mechanisms and Strategies. Immunity 2018, 48, 417–433.
- Ribas, A.; Dummer, R.; Puzanov, I.; VanderWalde, A.; Andtbacka, R.H.I.; Michielin, O.; Olszanski, A.J.; Malvehy, J.; Cebon, J.; Fernandez, E.; et al. Oncolytic Virotherapy Promotes Intratumoral T Cell Infiltration and Improves Anti-PD-1 Immunotherapy. Cell 2017, 170, 1109–1119.e10.
- Gandhi, L.; Rodríguez-Abreu, D.; Gadgeel, S.; Esteban, E.; Felip, E.; De Angelis, F.; Domine, M.; Clingan, P.; Hochmair, M.J.; Powell, S.F.; et al. Pembrolizumab plus Chemotherapy in Metastatic Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 2078–2092.
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.-J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 381, 1535–1546.
- Motzer, R.J.; Penkov, K.; Haanen, J.; Rini, B.; Albiges, L.; Campbell, M.T.; Venugopal, B.; Kollmannsberger, C.; Negrier, S.; Uemura, M.; et al. Avelumab plus Axitinib versus Sunitinib for Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2019, 380, 1103–1115.
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723.
- Morse, M.A.; Gwin, W.R.; Mitchell, D.A. Vaccine Therapies for Cancer: Then and Now. Target. Oncol. 2021, 16, 121–152.
- Igarashi, Y.; Sasada, T. Cancer Vaccines: Toward the Next Breakthrough in Cancer Immunotherapy. J. Immunol. Res. 2020, 2020, 5825401.
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-Related Adverse Events Associated with Immune Checkpoint Blockade. N. Engl. J. Med. 2018, 378, 158–168.
- Dusetzina, S.B.; Winn, A.N.; Abel, G.A.; Huskamp, H.A.; Keating, N.L. Cost sharing and adherence to tyrosine kinase inhibitors for patients with chronic myeloid leukemia. J. Clin. Oncol. 2014, 32, 306–311.
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1–Positive Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833.
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N. Engl. J. Med. 2015, 373, 23–34.
- Zraik, I.M.; Heß-Busch, Y. Management of chemotherapy side effects and their long-term sequelae. Urol. Ausg. A 2021, 60, 862–871.
- Obeid, M.; Tesniere, A.; Ghiringhelli, F.; Fimia, G.M.; Apetoh, L.; Perfettini, J.-L.; Castedo, M.; Mignot, G.; Panaretakis, T.; Casares, N.; et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat. Med. 2007, 13, 54–61.
- Wainwright, D.A.; Dey, M.; Chang, A.; Lesniak, M.S. Targeting tregs in malignant brain cancer: Overcoming IDO. Front. Immunol. 2013, 4, 116.
- Ma, Z.; Zhang, W.; Dong, B.; Xin, Z.; Ji, Y.; Su, R.; Shen, K.; Pan, J.; Wang, Q.; Xue, W. Docetaxel remodels prostate cancer immune microenvironment and enhances checkpoint inhibitorbased immunotherapy. Theranostics 2022, 12, 4965–4979.
- Sasso, M.S.; Lollo, G.; Pitorre, M.; Solito, S.; Pinton, L.; Valpione, S.; Bastiat, G.; Mandruzzato, S.; Bronte, V.; Marigo, I.; et al. Low dose gemcitabine-loaded lipid nanocapsules target monocytic myeloid-derived suppressor cells and potentiate cancer immunotherapy. Biomaterials 2016, 96, 47–62.
- Hauner, K.; Maisch, P.; Retz, M. Nebenwirkungen der Chemotherapie. Urol. Ausg. A 2017, 56, 472–479.
- Farkona, S.; Diamandis, E.P.; Blasutig, I.M. Cancer immunotherapy: The beginning of the end of cancer? BMC Med. 2016, 14, 73.
- Emens, L.A.; Cruz, C.; Eder, J.P.; Braiteh, F.; Chung, C.; Tolaney, S.M.; Kuter, I.; Nanda, R.; Cassier, P.A.; Delord, J.P.; et al. Long-term Clinical Outcomes and Biomarker Analyses of Atezolizumab Therapy for Patients with Metastatic Triple-Negative Breast Cancer: A Phase 1 Study. JAMA Oncol. 2019, 5, 74–82.
- Hinshaw, D.C.; Shevde, L.A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019, 79, 4557–4567.
- Abshire, D.; Lang, M.K. The Evolution of Radiation Therapy in Treating Cancer. Semin. Oncol. Nurs. 2018, 34, 151–157.
- Dewan, M.Z.; Galloway, A.E.; Kawashima, N.; Dewyngaert, J.K.; Babb, J.S.; Formenti, S.C.; Demaria, S. Fractionated but not single-dose radiotherapy induces an immune-mediated abscopal effect when combined with anti-CTLA-4 antibody. Clin. Cancer Res. 2009, 15, 5379–5388.
- Golden, E.B.; Demaria, S.; Schiff, P.B.; Chachoua, A.; Formenti, S.C. An abscopal response to radiation and ipilimumab in a patient with metastatic non-small cell lung cancer. Cancer Immunol. Res. 2013, 1, 365–372.
- Weichselbaum, R.R.; Liang, H.; Deng, L.; Fu, Y.X. Radiotherapy and immunotherapy: A beneficial liaison? Nat. Rev. Clin. Oncol. 2017, 14, 365–379.
- Gupta, A.; Probst, H.C.; Vuong, V.; Landshammer, A.; Muth, S.; Yagita, H.; Schwendener, R.; Pruschy, M.; Knuth, A.; van den Broek, M. Radiotherapy Promotes Tumor-Specific Effector CD8+ T Cells via Dendritic Cell Activation. J. Immunol. 2012, 189, 558–566.
- Barker, H.E.; Paget, J.T.E.; Khan, A.A.; Harrington, K.J. The tumour microenvironment after radiotherapy: Mechanisms of resistance and recurrence. Nat. Rev. Cancer 2015, 15, 409–425.
- Twyman-Saint Victor, C.; Rech, A.J.; Maity, A.; Rengan, R.; Pauken, K.E.; Stelekati, E.; Benci, J.L.; Xu, B.; Dada, H.; Odorizzi, P.M.; et al. Radiation and dual checkpoint blockade activate non-redundant immune mechanisms in cancer. Nature 2015, 520, 373–377.
- Lugade, A.A.; Moran, J.P.; Gerber, S.A.; Rose, R.C.; Frelinger, J.G.; Lord, E.M. Local Radiation Therapy of B16 Melanoma Tumors Increases the Generation of Tumor Antigen-Specific Effector Cells That Traffic to the Tumor. J. Immunol. 2005, 174, 7516–7523.
- Antonia, S.J.; Villegas, A.; Daniel, D.; Vicente, D.; Murakami, S.; Hui, R.; Yokoi, T.; Chiappori, A.; Lee, K.H.; De Wit, M.; et al. Durvalumab after Chemoradiotherapy in Stage III Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 1919–1929.
- Galluzzi, L.; Humeau, J.; Buqué, A.; Zitvogel, L.; Kroemer, G. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat. Rev. Clin. Oncol. 2020, 17, 725–741.
- Shuel, S.L. Targeted cancer therapies. Can. Fam. Physician 2022, 68, 515–518.
- Huang, R.-Y.; Eppolito, C.; Lele, S.; Shrikant, P.; Matsuzaki, J.; Odunsi, K. LAG3 and PD1 co-inhibitory molecules collaborate to limit CD8+ T cell signaling and dampen antitumor immunity in a murine ovarian cancer model. Oncotarget 2015, 6, 27359–27377.
- Beatty, G.L.; Gladney, W.L. Immune escape mechanisms as a guide for cancer immunotherapy. Clin. Cancer Res. 2015, 21, 687–692.
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726.
- Zitvogel, L.; Galluzzi, L.; Smyth, M.J.; Kroemer, G. Mechanism of Action of Conventional and Targeted Anticancer Therapies: Reinstating Immunosurveillance. Immunity 2013, 39, 74–88.
- Yun, C.O.; Hong, J.W.; Yoon, A.R. Current clinical landscape of oncolytic viruses as novel cancer immunotherapeutic and recent preclinical advancements. Front. Immunol. 2022, 13, 953410.
- Parato, K.A.; Senger, D.; Forsyth, P.A.J.; Bell, J.C. Recent progress in the battle between oncolytic viruses and tumours. Nat. Rev. Cancer 2005, 5, 965–976.
- Wang, W.; Liu, S.; Dai, P.; Yang, N.; Wang, Y.; Giese, R.A.; Merghoub, T.; Wolchok, J.; Deng, L. Elucidating mechanisms of antitumor immunity mediated by live oncolytic vaccinia and heat-inactivated vaccinia. J. Immunother. Cancer 2021, 9, e002569.
- Lin, J.; Sun, S.; Zhao, K.; Gao, F.; Wang, R.; Li, Q.; Zhou, Y.; Zhang, J.; Li, Y.; Wang, X.; et al. Oncolytic Parapoxvirus induces Gasdermin E-mediated pyroptosis and activates antitumor immunity. Nat. Commun. 2023, 14, 224.
- Enekegho, L.; Stuart, D. Smart Oncolytic Adenovirotherapy to Induce Killing of Cancer Cells and Elicit Antitumor Immunity. Eureka 2022, 7, 1–18.
- Zuo, S.; Wei, M.; Xu, T.; Kong, L.; He, B.; Wang, S.; Wang, S.; Wu, J.; Dong, J.; Wei, J. An engineered oncolytic vaccinia virus encoding a single-chain variable fragment against TIGIT induces effective antitumor immunity and synergizes with PD-1 or LAG-3 blockade. J. Immunother. Cancer 2021, 9, e002843.
- Das, K.; Belnoue, E.; Rossi, M.; Hofer, T.; Danklmaier, S.; Nolden, T.; Schreiber, L.-M.; Angerer, K.; Kimpel, J.; Hoegler, S.; et al. A modular self-adjuvanting cancer vaccine combined with an oncolytic vaccine induces potent antitumor immunity. Nat. Commun. 2021, 12, 5195.
- Zhang, B.; Shu, Y.; Hu, S.; Qi, Z.; Chen, Y.; Ma, J.; Wang, Y.; Cheng, P. In Situ Tumor Vaccine Expressing Anti-CD47 Antibody Enhances Antitumor Immunity. Front. Oncol. 2022, 12, 897561.
- Hofer, T.; Rossi, M.; Carboni, S.; Di Berardino Besson, W.; von Laer, D.; Wollmann, G.; Derouazi, M.; Santiago-Raber, M.L. Heterologous Prime-Boost Vaccination with a Peptide-Based Vaccine and Viral Vector Reshapes Dendritic Cell, CD4+ and CD8+ T Cell Phenotypes to Improve the Antitumor Therapeutic Effect. Cancers 2021, 13, 6107.
- Bartlett, D.L.; Liu, Z.; Sathaiah, M.; Ravindranathan, R.; Guo, Z.; He, Y.; Guo, Z.S. Oncolytic viruses as therapeutic cancer vaccines. Mol. Cancer 2013, 12, 103.
- Russell, S.J.; Peng, K.W. Oncolytic Virotherapy: A Contest between Apples and Oranges. Mol. Ther. 2017, 25, 1107–1116.
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379.
- Pol, J.; Kroemer, G.; Galluzzi, L. First oncolytic virus approved for melanoma immunotherapy. Oncoimmunology 2016, 5, e1115641.
- Harrington, K.; Freeman, D.J.; Kelly, B.; Harper, J.; Soria, J.C. Optimizing oncolytic virotherapy in cancer treatment. Nat. Rev. Drug Discov. 2019, 18, 689–706.
- Goradel, N.H.; Baker, A.T.; Arashkia, A.; Ebrahimi, N.; Ghorghanlu, S.; Negahdari, B. Oncolytic virotherapy: Challenges and solutions. Curr. Probl. Cancer 2021, 45, 100639.
- Mondal, M.; Guo, J.; He, P.; Zhou, D. Recent advances of oncolytic virus in cancer therapy. Hum. Vaccines Immunother. 2020, 16, 2389–2402.
- Niemann, J.; Kühnel, F. Oncolytic viruses: Adenoviruses. Virus Genes 2017, 53, 700–706.
- Moaven, O.; Mangieri, C.W.; Stauffer, J.A.; Anastasiadis, P.Z.; Borad, M.J. Strategies to Develop Potent Oncolytic Viruses and Enhance Their Therapeutic Efficacy. JCO Precis. Oncol. 2021, 1, 733–743.
- Gubin, M.M.; Zhang, X.; Schuster, H.; Caron, E.; Ward, J.P.; Noguchi, T.; Ivanova, Y.; Hundal, J.; Arthur, C.D.; Krebber, W.J.; et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 2014, 515, 577–581.
- Wong, S.Q.; Fellowes, A.; Doig, K.; Ellul, J.; Bosma, T.J.; Irwin, D.; Vedururu, R.; Tan, A.Y.-C.; Weiss, J.; Chan, K.S.; et al. Assessing the clinical value of targeted massively parallel sequencing in a longitudinal, prospective population-based study of cancer patients. Br. J. Cancer 2015, 112, 1411–1420.
- McGranahan, N.; Swanton, C. Clonal Heterogeneity and Tumor Evolution: Past, Present, and the Future. Cell 2017, 168, 613–628.
- Miao, D.; Van Allen, E.M. Genomic determinants of cancer immunotherapy. Curr. Opin. Immunol. 2016, 41, 32–38.
- Łuksza, M.; Riaz, N.; Makarov, V.; Balachandran, V.P.; Hellmann, M.D.; Solovyov, A.; Rizvi, N.A.; Merghoub, T.; Levine, A.J.; Chan, T.A.; et al. A neoantigen fitness model predicts tumour response to checkpoint blockade immunotherapy. Nature 2017, 551, 517–520.
- O’Donnell, T.J.; Rubinsteyn, A.; Bonsack, M.; Riemer, A.B.; Laserson, U.; Hammerbacher, J. MHCflurry: Open-Source Class I MHC Binding Affinity Prediction. Cell Syst. 2018, 7, 129–132.e4.
- Kalaora, S.; Wolf, Y.; Feferman, T.; Barnea, E.; Greenstein, E.; Reshef, D.; Tirosh, I.; Reuben, A.; Patkar, S.; Levy, R.; et al. Combined analysis of antigen presentation and T-cell recognition reveals restricted immune responses in melanoma. Cancer Discov. 2018, 8, 1366–1375.
- Bjerregaard, A.M.; Nielsen, M.; Hadrup, S.R.; Szallasi, Z.; Eklund, A.C. MuPeXI: Prediction of neo-epitopes from tumor sequencing data. Cancer Immunol. Immunother. 2017, 66, 1123–1130.
- Lu, Y.C.; Robbins, P.F. Cancer immunotherapy targeting neoantigens. Semin. Immunol. 2016, 28, 22–27.
- Dubensky, T.W.; Reed, S.G. Adjuvants for cancer vaccines. Semin. Immunol. 2010, 22, 155–161.
- Zhou, J.Z.; Jou, J.; Cohen, E. Vaccine strategies for human papillomavirus-associated head and neck cancers. Cancers 2022, 14, 33.
- Heathman, T.R.; Nienow, A.W.; McCall, M.J.; Coopman, K.; Kara, B.; Hewitt, C.J. The translation of cell-based therapies: Clinical landscape and manufacturing challenges. Regen. Med. 2015, 10, 49–64.
- Pollock, J.; Coffman, J.; Ho, S.V.; Farid, S.S. Integrated continuous bioprocessing: Economic, operational, and environmental feasibility for clinical and commercial antibody manufacture. Biotechnol Prog. 2017, 33, 854–866.
- Zhang, X.; Sharma, P.K.; Peter Goedegebuure, S.; Gillanders, W.E. Personalized cancer vaccines: Targeting the cancer mutanome. Vaccine 2017, 35, 1094–1100.
- Hilf, N.; Kuttruff-Coqui, S.; Frenzel, K.; Bukur, V.; Stevanović, S.; Gouttefangeas, C.; Platten, M.; Tabatabai, G.; Dutoit, V.; Van Der Burg, S.H.; et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature 2019, 565, 240–245.
- Kloor, M.; Von Knebel Doeberitz, M. The immune biology of microsatellite-unstable cancer. Trends Cancer 2016, 2, 121–133.
- Yarchoan, M.; Johnson, B.A.; Lutz, E.R.; Laheru, D.A.; Jaffee, E.M. Targeting neoantigens to augment antitumour immunity. Nat. Rev. Cancer 2017, 17, 209–222.
- Palucka, K.; Banchereau, J. Dendritic-Cell-Based Therapeutic Cancer Vaccines. Immunity 2013, 39, 38–48.
- Saung, M.T.; Ke, X.; Howard, G.P.; Zheng, L.; Mao, H.Q. Particulate carrier systems as adjuvants for cancer vaccines. Biomater. Sci. 2019, 7, 4873–4887.
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 2015, 348, 124–128.
- Postow, M.A.; Callahan, M.K.; Wolchok, J.D. Immune checkpoint blockade in cancer therapy. J. Clin. Oncol. 2015, 33, 1974–1982.
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61.
- Snyder, A.; Makarov, V.; Merghoub, T.; Yuan, J.; Zaretsky, J.M.; Desrichard, A.; Walsh, L.A.; Postow, M.A.; Wong, P.; Ho, T.S.; et al. Genetic Basis for Clinical Response to CTLA-4 Blockade in Melanoma. N. Engl. J. Med. 2014, 371, 2189–2199.
- Mitchell, S.A. Cancer-Related Fatigue: State of the Science. PMR 2010, 2, 364–383.
- Vanpouille-Box, C.; Alard, A.; Aryankalayil, M.J.; Sarfraz, Y.; Diamond, J.M.; Schneider, R.J.; Inghirami, G.; Coleman, C.N.; Formenti, S.C.; Demaria, S. DNA exonuclease Trex1 regulates radiotherapy-induced tumour immunogenicity. Nat. Commun. 2017, 8, 15618.
- Van Tendeloo, V.F.; Van de Velde, A.; Van Driessche, A.; Cools, N.; Anguille, S.; Ladell, K.; Gostick, E.; Vermeulen, K.; Pieters, K.; Nijs, G.; et al. Induction of complete and molecular remissions in acute myeloid leukemia by Wilms’ tumor 1 antigen-targeted dendritic cell vaccination. Proc. Natl. Acad. Sci. USA 2010, 107, 13824–13829.
- Rosenblatt, J.; Glotzbecker, B.; Mills, H.; Vasir, B.; Tzachanis, D.; Levine, J.D.; Joyce, R.M.; Wellenstein, K.; Keefe, W.; Schickler, M.; et al. PD-1 blockade by CT-011, anti-PD-1 antibody, enhances ex vivo t-cell responses to autologous dendritic cell/myeloma fusion vaccine. J. Immunother. 2011, 34, 409–418.
- Maude, S.L.; Laetsch, T.W.; Buechner, J.; Rives, S.; Boyer, M.; Bittencourt, H.; Bader, P.; Verneris, M.R.; Stefanski, H.E.; Myers, G.D.; et al. Tisagenlecleucel in Children and Young Adults with B-Cell Lymphoblastic Leukemia. N. Engl. J. Med. 2018, 378, 439–448.
- Schuster, S.J.; Bishop, M.R.; Tam, C.S.; Waller, E.K.; Borchmann, P.; McGuirk, J.P.; Jäger, U.; Jaglowski, S.; Andreadis, C.; Westin, J.R.; et al. Tisagenlecleucel in Adult Relapsed or Refractory Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2019, 380, 45–56.
- Hartmann, J.; Schüßler-Lenz, M.; Bondanza, A.; Buchholz, C.J. Clinical development of CAR T cells—Challenges and opportunities in translating innovative treatment concepts. EMBO Mol. Med. 2017, 9, 1183–1197.
- Sanmamed, M.F.; Fernández-Landázuri, S.; Rodríguez, C.; Zárate, R.; Lozano, M.D.; Zubiri, L.; Perez-Gracia, J.L.; Martín-Algarra, S.; González, A. Quantitative cell-free circulating BRAFV600E mutation analysis by use of droplet digital PCR in the follow-up of patients with melanoma being treated with BRAF inhibitors. Clin. Chem. 2015, 61, 297–304.
This entry is offline, you can click here to edit this entry!