1. Crop Rotation Design
Historically, organic farms were mainly mixed farms with legume-rich, rotational and/or permanent grassland that was used for grazing and/or conserved forage production [
6,
25,
26]. The 2-to-5-year ley phase substantially reduces weed pressure that builds-up while annual food crops are grown in the rotation, with weed pressure being particularly high after field vegetable crops [
6,
25,
26,
27,
28,
29]. Stockless arable and horticultural crop production systems also have mixed grass/legume or pure legume leys in the rotations, but grassland management focuses primarily on optimizing N-balances and soil fertility for subsequent annual cash crops [
30,
31]. It is interesting to note that stockless, conventional arable farms with persistent and/or herbicide-resistant black-grass problems have started to re-introduced grass-leys and grazing livestock to control black-grass (Alopecurus myosuroides Huds.) in their rotations [
29]. Catch- and under-sown cover crops are also more widely used in organic crop rotations; although they are mainly included to prevent nutrient losses, they can also contribute to weed control [
32,
33].
The ley phase of the rotation was also shown to reduce the propagules of many soil and crop residue-borne diseases, as well as pests that affect annual crops (e.g., cereals and field vegetables) in the rotation [
6,
25,
26]. For example, there is evidence that the severity of foliar diseases (e.g.,
Septoria,
Fusarium) in wheat grown after grass clover leys is significantly lower when compared with wheat (
Triticum aestivum L.) crops grown after wheat, maize (
Zea mays L.) and/or potato (
Solanum tuberosum L.) [
24,
33]. An interaction between the position of wheat crops in the rotation and fertilization regimes (use of organic versus mineral NPK fertilizer) was also recently reported for Septoria severity in the UK [
33].
Septoria severity in mineral NPK-fertilized wheat crops was significantly (more than three times) higher when wheat was grown after wheat, compared with wheat grown after a grass-clover ley, whereas the difference in Septoria severity between pre-crops was not significant when crops were fertilized with manure, at the same total N-input level (
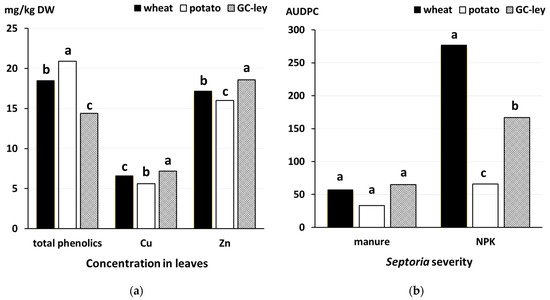
Figure 1) [
33]. However, for some diseases and pests the ley phase can also result in a build-up of inoculum. For example, long grass-clover leys can increase wireworm populations in soils and cause significant yield losses in potato crops that are planted immediately after a ley phase [
20,
34]. In this context, it is interesting to note, that in traditional, organic arable rotations in the UK wheat is grown immediately after leys, and then followed with a potato crop, primarily to reduce Septoria severity in wheat, and wireworm damage in potato (personal communication, Fred Halder, Murtle, Murtle Farm, Camphill Trust Aberdeen, UK).
Figure 1. Effect of (
a) preceding crops (wheat, potato, grass-clover [GC] leys) on total phenolic (
p = 0.001), Cu (
p = 0.002) and Zn (
p = 0.013) concentrations in wheat leaves and (
b) fertilization regime (manure versus mineral NPK fertilizer applied at the same total N-input rate of 170 kg N hectare) and preceding crops on
Septoria severity (
p < 0.001 for interaction) data from [
33]. Means for the same composition parameter or fertilizer type labelled with the same letter are not significantly different (Tukey’s honest significant difference test,
p < 0.05). AUDPC, area under the disease progress curve days x % severity.
The inclusion of legume crops in the rotation is prescribed by organic farming standards, and is an important component of the fertility management regime in organic farming systems. In between leys, organic farming regulations prescribe that farmers alternate crops from different plant families, and with contrasting life-cycles, canope structures, rooting patterns and periods of growth, to minimize the accumulation of crop species-specific weeds, disease and pest populations [
6,
25,
26,
27,
28]. Organic rotations are therefore thought to be, overall, more diverse than conventional crop rotations [
31], but there is concern that organic ley/arable rotations are not diverse enough to deliver the desired agronomic gains (e.g., reduced weed pressure; optimum nutrient-use efficiency) [
35,
36]. Therefore, the identification of agronomically useful and economically viable break crops for organic ley/arable rotations remains a major challenge [
36].
There have been few studies into the effect of rotation design on nutritional quality and safety parameters in crops. However, one clear example of rotational effects is the use of monoculture or less diverse arable crop rotations in conventional cereal production (e.g., planting wheat after wheat or maize pre-crops, which is common in conventional but not organic rotations) which has been shown to increase Fusarium disease severity, and Fusarium mycotoxin load in cereal grains [
37,
38,
39,
40].
Moreover, the crops grown before wheat in the rotation were shown to affect (i) leaf phenolic and mineral micronutrient concentrations, and (ii) foliar disease severity in the long-term, factorial Nafferton Farming Systems Comparison (NFSC) Trials [
33]. Specifically, this study reported that potato as the pre-crop resulted in significantly higher leaf phenolics, but lower Cu and Zn concentrations than wheat and grass clover leys in both manure and mineral NPK-fertilized crops (
Figure 1) [
33]. More recently, faba bean (
Vicia faba L.) cover crops were reported to result in the highest mineral, protein and prebiotic carbohydrate concentrations in subsequent kale (
Brassica oleracea L.) crops, when compared with Windham winter pea (
Pisum sativum L), hairy vetch (Vicia villosa Roth.), Lynx winter pea, crimson clover (
Trifolium incarnatum L.) and ryegrass (
Lolium perenne L.) [
41].
2. Fertilization Regimes
In intensive, conventional farming, the use of N, P, and KCl fertilizers has increased rapidly over the last 40 years. Whereas nutrient-use efficiency (the amount of mineral NPK needed to produce a kg of crops) has decreased, the dependence of conventional farming on NPK inputs to maintain current productivity levels has increased [
42]. There is now increasing concern about the rapidly rising cost, environmental impact and future availability of mineral NPK fertilizers, since their production requires high amounts of fossil fuels (mineral N), and because they are mined minerals and therefore non-renewable resources (mineral K and P) [
42,
43,
44,
45,
46]. For example, the manufacture and use of mineral N-fertilizer has been estimated to generate ~25% of all greenhouse gas (GHG) emissions from agriculture (=2.5% of all GHG-emissions) [
46], and according to some independent estimates, the currently known deposits of phosphorus will deplete within the next 60–150 years [
43,
44,
45].
In contrast, traditionally organic and biodynamic farming systems aim to create closed, on-farm nutrient cycles with a minimum import of nutrients from outside the farm [
4,
6,
25,
26,
47]. In mixed organic farming systems, this is primarily based on (i) livestock depositing manure onto leys during grazing, (ii) the strategic application of farm-yard manure (produced while livestock are housed in barns or corrals) to annual crops (e.g., cereals, potato, field vegetables) in the rotation, and (iii) including legume ley or grain crops in the rotation. Since the losses of N from manure and/or soil are larger than those of P and K, the use of N-fixing legume crops is also designed to compensate for N-losses, and provide a balanced supply of N:P:K [
6,
25,
26,
36]. Although the mixed farm is still considered the ideal model for organic farming, economic pressures have resulted in organic farms becoming increasingly specialized [
47]. As a result, many specialized arable and horticultural crop producers do not have livestock, and import larger amounts of nutrients in the form of organic materials (e.g., manure, waste-based compost or digestate from biogas-units), permitted mineral P (e.g., finely ground raw phosphate) and K (e.g., KSO
4), and often also mineral micro-nutrient fertilizer products [
6,
25,
26,
30,
47]. Stockless organic systems are (i) more dependent on imports of organic (manure, compost) and/or permitted mineral P, K and micronutrient fertilizers and (ii) more reliant on mechanical weed control and/or flame weeding, and therefore they are thought to be less sustainable when compared with mixed organic farms [
6,
25,
27,
30,
47].
It is important to note that the inclusion of energy crops in organic rotations, and the use of digestate from on-farm biogas units as fertilizer, has provided an alternative nutrient recycling option, especially for stockless organic producers [
30,
48,
49,
50]. Moreover, the use of biogas units for manure processing on mixed organic farms is thought to minimize N-losses, since both nitrification and denitrification are inhibited during the anaerobic digestion process [
48,
49,
50].
The beneficial effects of organic fertilization regimes on nutritional quality and safety parameters have been associated with (i) regular organic matter inputs on soil biological activity, (ii) a more balanced mineralization-driven nutrient (and especially N) supply pattern from organic fertilizers, and (iii) the non-use of mineral N and superphosphate. In contrast, there is limited information on the effect of different types of organic fertilizer (e.g., farm-yard manure, manure slurry, manure-compost manure, green-waste compost, household-waste compost, alfalfa pellets) on the nutritional composition of crops.
2.1. Replacing Mineral N-Fertilizer with Organic Fertilizer Inputs
There is now a range of studies that report that using organic fertilizers such as manure, instead of mineral NPK-based fertilization regimes, results in higher leaf and/or grain phenolic concentrations, but reduced levels of lodging in cereals and foliar disease severity in both cereals and other crops [
13,
33,
51,
52,
53,
54]. For example, in the NFSC trials, phenolic concentrations were significantly lower, whereas powdery mildew and/or rust severity were significantly higher in mineral NPK, compared with manure-fertilized wheat crops [
13,
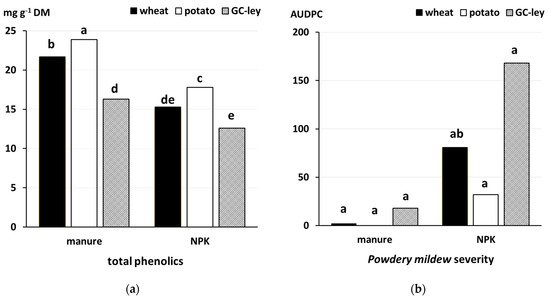
33]. In addition, in the years when the performance of wheat grown after contrasting preceding crops could be compared, wheat grown after grass-clover leys and mineral NPK as fertilizer had the highest mildew and lowest phenolics levels (
Figure 2). It is well known that phenolic compounds, which have antimicrobial activity, are a component of both constitutive and salicylic acid (SA)-inducible systemic resistance mechanisms in plants [
51]. Moreover, it has been demonstrated that increasing N-availability to plants, significantly reduces the concentrations of phenolic compounds and resistance against biotropic diseases in a dose-dependent manner [
52,
53,
54].
Figure 2. Effect of fertilization regime (F; manure versus mineral NPK fertilizer applied at the same total N-input rate of 170 kg N hectare) and preceding crop (PC) on (
a) total phenolic concentrations (
p = 0.089 for F×PC interaction), and (
b) powdery mildew severity (
p < 0.001 for FxPC interaction) in wheat leaves data from [
33]. Means labelled with the same letter are not significantly different (Tukey’s honest significant difference test,
p < 0.05). AUDPC, area under the disease progress curve days x % severity; DM, dry matter.
These results are of nutritional relevance, since phenolics are one of the main phytochemical groups with antioxidant activity in crops, and increased dietary intake of different types of phenolics has been linked to health benefits in humans, including a reduced risk of cardiovascular disease, type-2 diabetes and cancer [
55].
Results from the NFSC trials and other studies [
56,
57,
58] have suggested that replacing mineral NPK fertilizer inputs with manure at the same N-input level can also result in increased concentrations of other phytochemicals with antioxidant activity in crops, including vitamin C and folate in potato, cabbage (
Brassica oleracea L.) and lettuce (
Lactuca sativa L.), glucosinolates in cabbage, and total carotenoids in cabbage (
Table 1).
Table 1. Effect of the crop protection (organic versus conventional) and fertilization regime (mineral NPK vs cattle manure) on nutritionally relevant phytochemicals in potato, cabbage and lettuce grown in the NFSC trial data from [
58]; values shown are main effect means; yield results are from 9, 6, 4 and 4 growing seasons for potato, cabbage, lettuce and onion (
Allium cepa L.), respectively; phytochemical and/or nitrate results are from 3 or 4 growing seasons for potato, cabbage, lettuce and onion.
| |
|
|
|
ANOVA Results (p-Values) |
| |
|
Crop Protection (CP) |
Fertilization (F) |
Man Effects |
Inter-Action |
| Crop |
Parameter Assessed |
CON |
ORG |
Mineral NPK |
Cattle Manure |
CP |
F |
CP×F |
| Potato |
Marketable tuber yield (t/ha) |
9.2 |
7.7 |
9.3 |
7.6 |
<0.001 |
<0.001 |
0.003 |
| |
total phenolics (µg/g FW) |
285 |
293 |
277 |
301 |
NS |
0.016 |
NS |
| |
vitamin C (µg/g FW) |
95 |
97 |
91 |
101 |
NS |
<0.001 |
NS |
| |
total carotenoids (µg/g FW) |
0.7 |
0.8 |
0.7 |
0.7 |
0.060 |
NS |
0.018 |
| |
folate (µg/g FW) |
0.17 |
0.18 |
0.17 |
0.18 |
NS |
<0.001 |
0.006 |
| |
glycoalkaloids (mg/kg FW) |
42 |
40 |
45 |
37 |
NS |
<0.001 |
NS |
| Cabbage |
Yield (t/ha FW) |
54 |
61 |
71 |
45 |
<0.001 |
<0.001 |
0.007 |
| |
total phenolics 1 (µg/g FW) |
11.0 |
9.4 |
8.8 |
11.6 |
0.093 |
0.001 |
NS |
| |
vitamin C (µg/g FW) |
224 |
223 |
211 |
236 |
NS |
0.006 |
NS |
| |
total carotenoids (µg/g FW) |
3.5 |
3.2 |
3.0 |
3.7 |
NS |
0.008 |
0.051 |
| |
folate (µg/g FW) |
0.4 |
0.3 |
0.3 |
0.4 |
<0.001 |
0.022 |
NS |
| |
total glucosinolates (g/g FW) |
1.37 |
1.23 |
1.13 |
1.48 |
0.037 |
<0.001 |
NS |
| |
total volatiles 2 (mg/g FW) |
33 |
31 |
28 |
35 |
NS |
<0.001 |
0.032 |
| Lettuce |
Yield (t/ha FW) |
38 |
36 |
39 |
34 |
NS |
<0.001 |
NS |
| |
total phenolics (µg/g FW) |
104 |
108 |
102 |
112 |
NS |
0.036 |
NS |
| |
vitamin C (µg/g FW) |
6.8 |
6.7 |
6.8 |
6.7 |
NS |
NS |
NS |
| |
total carotenoids (µg/g FW) |
4.4 |
5.0 |
4.6 |
4.8 |
0.085 |
NS |
NS |
| |
folate (µg/g FW) |
0.3 |
0.3 |
0.3 |
0.3 |
0.064 |
0.042 |
NS |
| Onion |
Marketable yield (t/ha FW) |
22 |
20 |
21 |
21 |
NS |
NS |
NS |
| |
total phenolics 3 (µg/g FW) |
725 |
721 |
740 |
706 |
NS |
NS |
NS |
| |
vitamin C (µg/g FW) |
100 |
93 |
86 |
107 |
NS |
NS |
NS |
| |
total carotenoids (µg/g FW) |
0.6 |
0.7 |
0.7 |
0.6 |
NS |
NS |
0.083 |
| |
folate (µg/g FW) |
0.4 |
0.4 |
0.4 |
0.4 |
NS |
NS |
NS |
In contrast, concentrations of toxic glycoalkaloids in potato tubers were significantly lower when manure, instead of mineral NPK, was used as fertilizer in the NFSC trials (
Table 1). However, the effects of mineral NPK versus organic fertilizer use on non-resistance related phytochemicals are less consistent than those observed for phenolics. For example, in the NFSC trials, the use of manure instead of mineral NPK resulted in slightly (~5–10%), but significantly, lower concentrations of tocopherol and tocotrienol in wheat grain [
58].
Differences in the N-supply pattern from organic and mineral N-fertilizer are also thought to, at least partially, explain the results of (i) the NFSC trials, and (ii) recent systematic reviews/meta-analyses and retail surveys that reported higher levels of antioxidant activity and concentrations of phytochemicals with antioxidant activity in organic crops, compared with conventional crops [
56,
57,
58,
59,
60]. However, previous studies and the NFSC trial results demonstrated that rotation design, variety choice and crop protection regimes may have also contributed to differences in phytochemical concentrations and antioxidant activity levels between organic and conventional crops [
7,
12,
13,
19,
50,
51,
53,
57,
58,
59,
60] (
Table 1;
Figure 3).
Mineral N-fertilizer use is thought to be the main reason for the significantly higher protein, nitrate and nitrite concentrations found in conventional crops, compared with organic crops in meta-analyses and the NFSC trials, but there is uncertainty about the potential nutritional and health impacts [
57].
2.2. Replacing Superphosphate with Organic Fertilizer Inputs
The use of water-soluble P-fertilizers (superphosphate, triple superphosphate) is known to suppress mycorrhizal development in crop plants [
61]. Mycorrhizal fungal associations have been shown to provide benefits to crop plants, including (i) improved uptake efficiency of P and micronutrients such as Zn, Cu and Se, (ii) increased resistance against soil-borne diseases, and (iii) greater tolerance against abiotic stress, including drought [
61,
62].
Organic crop management has been reported to result in higher densities and diversity of mycorrhizal fungal inocula in soil, and increased colonization and mycorrhizal development on plant roots [
61,
62,
63]. The non-use of water-soluble P-fertilizers in organic farming systems may, therefore have (i) contributed to the positive impacts of organic fertilization regimes on crop health (see e.g.,
Figure 1 and
Figure 2) and/or (ii) explained the higher concentrations of nutritionally desirable mineral micronutrients detected in organic crops in some comparative studies [
57,
59].
It is well known that the application of water-soluble mineral-P fertilizer results in an increase in the concentration of toxic metal cadmium (Cd) in crop plants [
64,
65,
66]. All phosphorus fertilizers contain Cd as a contaminant and levels may vary from trace amounts, to as much as 300 mg Cd kg
−1 of dry product [
64]. Fertilization can influence Cd accumulation in crops by direct Cd addition with P-fertilizers, and by indirect effects on rhizosphere chemistry, soil microbial activity, soil pH and Zn concentration, and by affecting plant growth [
64,
65,
66].
Systematic reviews and meta-analyses reported that organic cereals have significantly lower Cd concentrations, but there is insufficient published information for other crops [
57]. Results from the NFSC trials demonstrated that the use of mineral NPK fertilizer results in significantly higher Cd concentrations, when compared with manure at the same total N-input level, in a range of crops, including wheat, potato, cabbage, onion and lettuce (
Figure 3). However, it is important to point out that, in the meta-analyses of published data, significant differences in Cd concentrations between organic and conventional cereals were only detected in experimental studies, whereas analyses of farm and retail survey data showed no significant differences for Cd [
57]. In most experimental studies (including the NFSC trials), no permitted mineral-P fertilizer was applied in the organic management plots [
57,
58,
65], whereas finely ground rock-phosphate (which also contains Cd as a contaminant) is used on many commercial organic farms, and especially by stockless arable and specialist horticultural producers [
6,
25,
26]. This may explain the contrasting results obtained in experimental studies and farm/retail surveys, and indicates that there is potential to reduce Cd exposure by reducing mineral P-inputs in both organic and conventional farming.
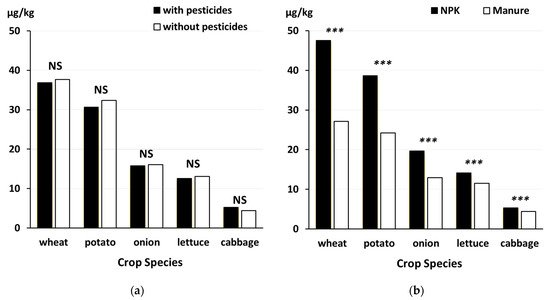
Figure 3. Effect of contrasting (
a) crop protection (with and without the use of synthetic chemical pesticides) and (
b) fertilization regimes (mineral NPK versus manure applied at the same total N-input rate of 170 kg N hectare) used in organic and conventional farming on cadmium concentrations in wheat grain, potato tubers, onions, cabbage heads and lettuce leaves Data from [
58,
65]. Bars represent the main effect means of data from four (potato, onion, lettuce, cabbage) or five (wheat) growing seasons. ***, significant difference (
p < 0.001); NS, not significant.
In this context, it should be noted that the wheat, potato and vegetable cultivars used in the NFSC trials were varieties from conventional farming-focused breeding programs, which were selected for performance in conventional mineral NPK-fertilized production systems.
2.3. Rhizobium and Mycorrhizal Fungal Inocula
Organic farming standards permit the use of symbiotic microbial inocula that improve N-fixation in soil (e.g., Rhizobium or free-living N-fixing bacteria) and nutrient uptake by plants (e.g., VA-mycorrhizal fungi) [
4]. However, there is controversy about whether such inocula are necessary and deliver substantial benefits on mixed organic farms with diverse rotations that have been under organic management for many years [
6,
25,
26].
Rhizobium inocula are used in both conventional and organic farming, especially for grain legumes [
67,
68,
69,
70,
71] and were shown to increase N-fixation and soil-N and N/crude protein in grain legume and cereal crops grown after inoculated grass-clover leys [
69]. Rhizobium species are legume species-specific and when new legume crops (e.g., soya) are introduced into rotations, treatments of seed with Rhizobium inocula is recommended [
71]. The use of Rhizobium inocula was also reported to increase mycorrhizal development from ‘natural’ mycorrhizal fungal inocula present in soil [
70]. The combined use of (i) Rhizobium inocula for clover during the establishment of a two-year clover ley and (ii) application of domestic waste compost with a high C:N ratio was proposed as a strategy to increase N-fixation and avoid N-losses [
69]. N-losses can be substantial when grass clover leys are incorporated into soil and the combined use of Rhizobium inocula and high C:N ratio compost was demonstrated to substantially increase both yield and protein concentration in organic wheat crops established after leys [
69]. There is, to our knowledge, limited published information on the effects of Rhizobium inocula on other nutritionally relevant quality parameters.
Mycorrhizal inocula are not widely used in both conventional and organic food crop production, although a lot of research has been done on their development [
71,
72,
73,
74,
75,
76]. This is thought to be mainly because arbuscular mycorrhizal fungal (AMF) inoculum products are relatively expensive, and produce inconsistent and sometimes negative effects on crop yields [
71,
72,
73,
74,
75,
76]. Moreover, it remains unclear whether, and to what extent, AMF inocula increase P and mineral micronutrient uptake and concentrations in crop plants [
76].
The effect of AMF inocula on vitamin and antioxidant levels in vegetables was compared in manure and mineral NPK-fertilized crops in the Rodale Institute’s long-term farming system trial [
72]. AMF-inoculation significantly reduced vitamin C and antioxidant levels in carrots (
Dacus sativus Hoff.), but had no consistent effect in bell peppers (
Capsicum annuum L.) or tomato (
Solanum lycopersicum L.) crops [
72]. Interestingly, the reduction in vitamin C from AMF-inocula was significantly greater in the mineral NPK-fertilized conventional system (87%) than in the manure-fertilized organic system (28%) [
72].
3. Crop Protection
3.1. Soil/Substrate Suppressiveness against Pests and Diseases
Regular inputs of organic matter have been shown to increase the organic matter content, biological activity, microbial and invertebrate biodiversity, structural stability and erosion resistance of agricultural soils [
77,
78,
79,
80].
Organic matter inputs, in particular composts produced in controlled aerobic composting processes, were also shown to increase (i) the suppressiveness of soils against soil-borne bacterial and fungal diseases and nematodes [
81,
82,
83,
84,
85,
86,
87,
88,
89,
90], and (ii) the populations of beneficial invertebrates (e.g., ground beetles) in soils [
80,
87,
88]. Close associations between the increase in soil biological activity and disease suppressiveness associated with organic matter/compost inputs have also frequently been observed [
83,
84,
86]. Suppressive composts can be made from a range of organic feedstocks and are widely used in commercial organic horticultural production systems [
86,
87,
88].
An extensive review of published studies by Bonanomi and co-workers [
87] examined 252 scientific papers and reported positive correlations between disease suppressiveness of soils after compost/organic matter inputs and (i) soil biological activity, and (ii) concentrations of specific groups of antagonistic soil microorganisms (e.g., Trichoderma species, fluorescent pseudomonads) in soil. The finding that soil chemical or steam disinfection was shown to significantly reduce or eradicate disease suppressiveness also suggests that components of soil biota are a major driver for “disease suppressiveness” [
87].
However, some organic waste materials used as feed material for compost were also shown to reduce disease severity via (i) inducing crop resistance mechanisms (e.g., the chitin soil amendments made from shellfish processing waste) [
83,
84] or (ii) releasing phytochemicals (e.g., tannins, phenolic compounds and isothiocyanates) that directly inhibit pathogen development in the soil (e.g., Brassica crop residues/waste or bark from certain tree species) [
83,
84,
88,
89,
90]. It is interesting to note that the use of Brassica cover crops/green manures, which have a “biofumigation” effect when incorporated into the soil, are now recommended as an alternative method to chemical soil disinfection for the control of parasitic nematodes in both conventional and organic farming [
90].
There is limited information on the effect of organic soil amendments on the nutritional composition of crop plants. However, the use of soil amendments that induce crop resistance (e.g., chitosan, chitin) has been linked to higher phenolic concentrations and antioxidant activity in vegetables, herbs and fruit and/or increased antioxidative and antimutagenic activity in organic green vegetables [
91,
92,
93].
3.2. Environmental Enrichment Practices
Environmental enrichment/diversification practices include the use of (i) companion plants and intercrops to distract invertebrate pests and thereby reduce pest damage, and (ii) managed field margins and/or within-field beetle banks to maintain high population densities of natural enemies of crop pests and/or pollinators [
80,
94,
95,
96,
97,
98,
99,
100] (
https://youtu.be/XvPP6W0m1GA, accessed 30 November 2021). The use of diversification strategies is thought to be more prevalent in organic farming systems. Moreover, natural enemy and pollinator density—but not necessarily diversity—was reported to be greater on organic farms [
96,
97,
101,
102] However, in most studies it remained unclear to what extent the effects on pest and pollinator populations were due to the non-use of insecticides or environmental enrichment practices used on organic farms.
The studies of natural enemy populations by Eyre and co-workers [
80,
95,
96,
97] suggested that a wide range of parameters, including crop type, and contrasting fertilization and crop protection methods used in organic and conventional farming, but also field margin vegetation and landscape features, affect natural enemy populations. For example, in the NFSC trial plots, ground beetle abundance was greater in many crops when manure instead of mineral NPK was used as fertilizer. However, abundance was reduced by insecticide applications [
80]. In contrast, in wheat crops, the abundance of spiders was greater in mineral NPK, compared with manure-fertilized plots [
80].
There is, to our knowledge, no information on the effects of environmental enrichment on the nutritional composition of crop plants, apart from contributing to crops being grown with lower or no pesticide inputs. However, the benefits of organic farming practices on pollinator populations are thought to improve the sustainability of crop production and future food security [
96,
97,
101,
102].
3.3. Elicitors of Crop Resistance
Elicitors permitted for use in organic farming are produced from plant extracts or organic waste materials, and are primarily used for the control of seed-borne [
103,
104], soil-borne [
83,
84,
91] and foliar diseases [
51,
91,
92,
93,
105,
106,
107]. The main compounds currently available and used by commercial organic farmers are:
-
Chitin (a long-chain polymer of N-acetylglucosamine, which is a primary component of fungal cell walls and the exoskeleton of insects and crustaceans) is mainly produced from the shells of shrimps and other crustaceans;
-
Chitosan (a linear polysaccharide composed of randomly distributed β-(1→4)-linked D-glucosamine and N-acetyl-D-glucosamine) is produced by treating the chitin shells of shrimp and other crustaceans with an alkaline substance, such as sodium hydroxide;
-
Giant knotweed (Reynoutria sachalinensis Schmidt ex Maxim.) extracts (tradename: Milsana®).
Soil and foliar chitin/chitosan, and foliar Giant knotweed extract treatments were shown to induce salicylic acid-dependent defense responses in plants, which includes the increased expression of phenolics and other phytochemicals with antioxidant activity in crop plants [
51,
91,
92,
93,
107]. For example, the large difference in antioxidant activity between organic and conventional spinach (
Spinacia oleracea L.) and onion observed in Japan [
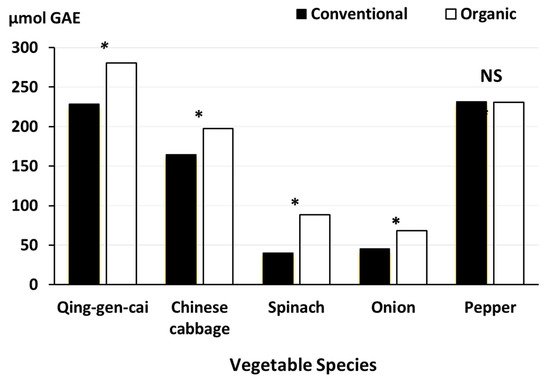
91] was linked to (i) 500 kg chitosan/ha being incorporated into the soil before planting, and (ii) crops being sprayed regularly with a 0.01% chitosan solution during the growing season (
Figure 4).
Figure 4. Effect of crop management (organic vs conventional) on antioxidant activity/capacity in vegetables. GAE, gallic acid equivalents data from [
91]. The organic (but not the conventional) crop management included the incorporation of 500 kg chitosan/ha before planting, and crops being sprayed regularly with a 0.01% chitosan solution during the growing season. *, significant difference (
p < 0.05); NS, not significant.
However, it is important to note that differences in fertilization regimes (e.g., the use of manure instead of mineral NPK fertilizer for organic vegetables) are also likely to have contributed to the higher antioxidant activity in organic crops observed in this study.
3.4. Biological Control Products
Biological pest and disease control products are permitted under EU organic farming regulations [
4] and pest control products based on
Bacillus thuringiensis (Bt), or natural enemies of invertebrate pests, are widely used [
6,
25,
26,
108].
Biological control products based on predators and parasitoids are now available for a wide range of invertebrate pests/herbivores, and are extensively used in both organic and conventional production systems and especially protected horticultural crops [
108]. It would go beyond the scope of this review to describe the different products and their mode of action, but detailed information on the products available to farmers can be found on the websites of the main producers/suppliers of biological control products (e.g., Koppert B.V., Berkel, The Netherlands;
www.koppert.com/products-solutions, accessed on 29 November 2021; Biobest Group NV, Westerlo, Belgium,
www.biobestgroup.com/en/biobest/products, accessed on 29 November 2021).
There has also been extensive research and development efforts to develop biological control products for crop diseases based on bacterial (e.g.,
Bacillus and
Pseudomonas spp.) and fungal antagonists (e.g.,
Trichoderma spp.) and a range of products are now commercially available and used in organic farming [
109,
110]. However, compared with Bt and natural enemy-based biological pest control products, biological disease control products are thought to be less widely used in organic farming [
83,
86].
Biological control products are not known to produce detectable chemical residues in foods, and there are currently no concerns about potential negative health impacts of the consumption of crops treated with biological crop-protection products [
111].
3.5. Barrier, Mating Disruption, and Mass-Trapping Methods for Pest Control
Barrier-based pest control strategies (e.g., the use of crop covers, netting and insect proof green and screenhouses) are permitted and widely used for pest control in organic horticultural crop production [
4,
112,
113]. However, compared with unprotected open field conditions, the use of insect-proof screens was reported to (i) increase relative humidity by 2–21%; (ii) reduce solar radiation by 50–87%; (iii) reduce air velocity by between 15 and 39%, and (iv) affect air temperature and evapotranspiration [
114]. These changes in environmental conditions are also known to have significant effects on crop growth, yield, and quality parameters [
114]. For example, they may explain the lower folate and glucosinolate concentrations in cabbage crops grown with organic crop protection regimes (which used insect-proof crop covers for pest control only), when compared with cabbage crops produced with conventional, pesticide application-based crop protection regimes (
Table 1). It is interesting to note that in the NFSC trials, cabbage was the only vegetable crop in which (i) netting was used in the organic crop protection protocol; (ii) significant main effects of crop protection on nutritionally desirable phytochemicals were detected, and (iii) significantly lower pest damage and higher crop yields were recorded in crops grown with organic regimes, compared with conventional crop protection regimes (
Table 1).
Organic farming standards also permit the use of pheromone sprays for mating disruption, the release of sterile males, and mass-trapping based on pheromones and nutrient attractants for pest control [
4]. These methods are extensively used by organic farmers, especially in perennial horticultural crops including apples (
Malus domestica Borkh.), citrus (
Citrus L.) and olives (
Olea europaea L.) [
115,
116,
117,
118,
119,
120], and are thought to have a substantially lower impact on natural enemy populations than (i) synthetic chemical insecticides used in conventional farming, and (ii) plant and microbial fermentation-based insecticides (pyrethrum, spinosad) that are permitted and used for pest control in both organic and conventional production [
120].
There is limited information on the effects of barrier, mating disruption, and mass-trapping methods on the nutritional quality of crops, although it should be noted that they can have a significant impact on minimizing pesticide use and residues in food [
115,
116,
117,
118,
119,
120].
3.6. Thermal Crop Protection Methods and Pesticide Products Used in Organic Farming
The use of thermal soil disinfection and weed control methods are permitted and used in organic farming, although the use of thermal soil disinfection is now restricted by many organic certification bodies [
4,
121]. This is mainly because thermal crop protection substantially increases fossil fuel use and the carbon footprint of food production, and was shown to have negative impacts on soil structure and biological activity [
121].
The use of pesticides made from mined minerals (e.g., S-fungicides and insecticides; Cu-fungicides and bactericides) is permitted in both organic and conventional farming systems [
4,
6,
25,
26,
122], although the use of Cu-products is restricted under EU-farming regulations to 6 kg Cu/ha/annum, and requires farmers to justify and obtain a derogation from their certification body [
4].
In the NFSC trials, Cu-fungicides (at a rate equivalent to 6 kg Cu/ha/annum) were only used in the organic crop protection regime for late blight control in potato crops [
20]. Conventional crop protection regimes for potatoes used synthetic chemical fungicides for late blight control. The finding that potato tubers from Cu-fungicide treated plots (organic crop protection) had significantly higher Cu concentrations than potatoes from plots treated with synthetic chemical fungicides (
Figure 6) suggests that the use of Cu-fungicide can increase Cu-concentration in crops. This view is supported by the finding that Cu-concentrations were not significantly different (lettuce, onion) or higher with conventional crop protection (cabbage) in the other vegetable crops grown in the NFSC trials (
Figure 5).
Figure 5. Effect of contrasting (
a) crop protection (CON, conventional crop protection based on synthetic chemical herbicides, fungicides and insecticides; ORG, organic crop protection based on mechanical weed control and hand weeding and the use of Cu-fungicides in potato only) and (
b) fertilization regime (mineral NPK versus manure applied at the same total N-input rate of 170 kg N hectare) used in organic and conventional farming on
copper concentrations in potato tubers, onions, cabbage heads and lettuce leaves, Data from [
58]. Bars represent main effect means of data from four growing seasons. ***, significant difference (
p < 0.001); *, significant difference (
p < 0.05); T, trend (0.01 <
p < 0.05); NS, not significant.
It is important to note that S and Cu are essential mineral nutrients for both plants and animals/humans, and that there are currently no concerns about potential negative health impacts of the consumption of crops treated with mineral-based crop-protection products [
4,
111,
122]. However, there is concern about (i) the potential health implications for farmers handling Cu-fungicides, because concentrated Cu solutions are highly toxic and can be taken up through the skin, and (ii) negative environmental impacts (e.g., reduced soil biological activity or phytotoxicity) of extensive Cu-fungicide use in orchard crops (e.g., apple, grapevine) [
111,
122,
123].
Organic farming standards permit the use of a small number of insecticide products based on plant extracts (e.g., pyrethrin and azadirachtin, which are made from chrysanthemum [
Chrysanthemum indicum L.] flowers and the neen tree [
Azadirachta A. Juss.] fruit, respectively) or microbial fermentation extracts (e.g., spinosad, which is made from the insecticidal chemicals spinosyn A and D produced by the actinomycete
Saccharopolyspora spinosa) for crop protection [
4].
Similar to synthetic chemical insecticides, pyrethrin, azadirachtin and spinosad may also have negative effects on non-target invertebrates including (i) natural enemy populations present in agricultural ecosystems and (ii) parasitoids and predators released into crops as biological control treatments [
124,
125].
Although they are considered to be less toxic than some groups of synthetic chemical insecticides (e.g., organophosphates, carbamates and neonicotinoids), there is some concern about the potential negative health impacts of exposure to pyrethrins and spinosad residues [
111,
126,
127].
3.7. Synthetic Chemical Pesticides That Are Prohibited in Organic Farming
Negative health effects of occupational (e.g., farmers and crop protection contractors handling pesticides) and environmental exposure to synthetic chemical pesticides are well documented, and there is also increasing evidence for negative health impacts of chronic dietary pesticide exposure [
126,
127,
128,
129,
130,
131,
132,
133,
134,
135,
136].
In the EU more than 300 different synthetic chemical pesticides (herbicides, insecticides, acaricides, fungicides, plant-growth regulators and soil-disinfecting chemicals) are licensed and widely used for crop protection in conventional food production [
128,
129]. In contrast, EU organic farming regulations prohibit the use of all synthetic, chemical pesticides for crop protection and as veterinary treatments [
4,
111].
Three comprehensive literature reviews and two meta-analyses of organic versus conventional crop composition data reported that levels of pesticide residues are substantially lower in organic than conventional foods, and concluded that organic food consumption may be one strategy to reduce dietary exposure to pesticides and associated health risks [
57,
137,
139]. For example, the most recent meta-analysis by Baranski and co-workers [
57] reported that organic crops are four times less likely to contain detectable pesticide residues than conventional crops, and that the level of pesticide contamination in conventional crops differs significantly between cereals, vegetables and fruit products (
Figure 6a). Specifically, conventional fruits were found to have substantially higher pesticide loads than conventional vegetables and cereal-based foods, whereas contamination levels were similar in organic fruits, vegetables and cereal-based foods, but substantially lower than in their conventional comparators (
Figure 6a). Similar results were obtained in recent retail surveys of fruits, vegetables and cereal-based foods [
141], which reported that the number of different pesticide residues detected was substantially higher in conventional products, compared with organic fruit, vegetables and cereal-based food products (
Figure 6b). A large retail survey of wheat flour products carried out over 3 years in the UK and Germany reported significant effects of wheat species (
Triticum aestivum L. vs.
T. spelta L. flour) and refining (white versus whole-grain flour) and the country in which flour samples were collected (the UK versus Germany), on pesticide residue concentrations in conventional, but not organic wheat flour, which generally contained very low pesticide residues [
140] (
Figure 7). Specifically, the reduction in pesticide exposure associated with organic food consumption was significantly higher with wholegrain, compared with refined flour, common wheat compared with spelt wheat flour, and in the UK compared with Germany (
Figure 7).
Figure 6. Effect of organic and conventional production methods on (
a) the percent of samples testing positive for at least one pesticide residue. Data from [
57], and (
b) the number of different pesticide residues detected in fruits, vegetable and cereal-based foods. Data from [
141]. #, cereals and cereal-based food products. ***, significant difference (
p < 0.001); **, significant difference (
p < 0.01); *, significant difference (
p < 0.05); T, trend (0.01 <
p < 0.05); NS, not significant.
Figure 7. Effect of organic and conventional production methods on the total pesticide residue concentrations in (
a) whole-grain and white/refined wheat flour, (
b) spelt and common wheat flour and (
c) wheat flour samples produced in the UK and Germany). Data from [
140]. Means/bars for the same production system (=same color) labelled with the same lower-case letter and means/bars for the same flour types, wheat species or country labeled with the same capital letters are not significant different (
p < 0.05).
These results are consistent with regulatory pesticide monitory data from Europe, which also suggest that pesticide residue levels in whole-grain cereal products, fruit and vegetables are higher than in food such as refined cereal products, and most meat and dairy products [
128].
It is important to note that the concentrations of pesticides in pesticide-treated crops may also be affected by both environmental conditions and agronomic practices other than pesticide use [
58,
142,
143,
144]. For example, wheat produced with conventional, pesticide-based crop protection regimes had significantly higher chlormequat levels in manure-fertilized plots, compared with mineral NPK-fertilized plots in the NFSC trial [
58] (
Figure 8). Chlormequat is a plant-growth regulator that is licensed in the EU only for use in cereals (especially wheat) to prevent lodging. The higher chlormequat residues in manure-fertilized crops are thought to be due to the 1–2-week earlier wheat grain maturity and senescence observed in manure, compared with NPK-fertilized wheat crops, which shortens the period available for the metabolism of chlormequat by the plant [
58]. Compound feeds made from manure-fertilized cereal, potato, carrot and onion crops grown in the NFSC trials and used in animal dietary intervention studies therefore contained significantly higher concentrations of chlormequat, than feeds produced with mineral NPK fertilizer [
142].
Figure 8. Effect of fertilization regimes (manure versus mineral NPK at the same total N-input level) on chlormequat residues in wheat grain produced in NFSC trial plots managed with conventional, pesticide-based crop protection. Data from [
58]. ***, significant difference (
p < 0.001). The maximum residue level (MRL) set by the EU for chlormequat residues in wheat grain is 7 mg/kg.
In contrast, concentrations of the insecticide aldicarb and the herbicide diquat were lower in compound feeds made from manure-fertilized crops, compared with mineral NPK-fertilized cereal, potato, carrot and onion crops, when feeds made from pesticide-treated crops were compared [
58,
143]. Aldicarb is an insecticide applied as granules to soils for soil-borne pests, and nematode control and was only used in potato crops, whereas Diquat is a semi-selective herbicide that was applied in potato and vegetable crops in the NFSC trials [
58,
143]. Soil-applied herbicides and insecticides are known to be absorbed by soil organic matter, and aldicarb metabolism in topsoil is thought to be mainly by microbial activity [
144,
145], which is known to be higher in soils receiving regular organic fertilizer inputs [
78,
146,
147,
148]. The lower levels of diquat and aldicarb found in crops from manure-fertilized soils are therefore thought to have been due to higher levels of microbial metabolism and/or absorption to soil organic matter, but this was not confirmed by soil analyses in the NFSC trials [
58].
The use of pesticides and especially herbicides can affect the metabolism and physiology of crop plants in many different ways [
149]. For example, herbicides were shown to inhibit photosynthesis and energy metabolism and/or inhibit root growth and nutrient uptake from soil [
149,
150,
151,
152,
153,
154]. This may lead to an increased susceptibility to crop diseases and lower concentrations of nutritionally desirable phytochemicals involved in photosynthesis (e.g., carotenoids), and mineral micronutrients (e.g., Ca, Mg, Fe, Zn) in plants [
149,
150,
151,
152,
153,
154]. For example, the negative effects of glyphosate-based herbicides (GBHs, the most widely used crop protection products globally) on crop health and nutritional composition were recently reviewed [
155] and include:
-
“Impairment of the innate physiological defenses by interruption of the shikimic acid pathway”;
-
“Interference with rhizosphere microbial ecology (in particular, GBHs have the potential to enhance the population and/or virulence of some phytopathogenic microbial species in the crop rhizosphere)”;
-
“As yet incompletely elucidated reduction in the uptake and utilization of nutrient metals by crops”.
4. Crop Breeding and Selection
The first organic and “low-input” farming-focused breeding/selection programs started in the late 1990s, and for many crop species there are still no organic-farming-specific breeding programs [
8,
9,
10,
11,
156,
157]. As a result, the majority of crop varieties used in organic farming were developed for conventional farming, and are therefore thought to lack important traits required for optimum crop yield, health and/or quality in organic systems [
7,
8,
9,
10,
11,
156,
157,
158].
However, organic farmers tend to use (i) different and often older, traditional cultivars and/or (ii) varieties with higher disease resistance and/or competitiveness against weeds [
7,
8,
9,
10,
11]. Many organic farmers also prefer to use open-pollinating varieties rather than hybrids, because they perceive them as having better sensory quality, or because they want to be able to save seed for the next season [
10,
158].
The approaches, standards and philosophy of organic plant breeding have recently been reviewed [
156,
157,
158] and are therefore not described in detail here. However, it is important to note several important characteristics of organic breeding programs: namely that (i) breeding and selection are farmer-participatory, (ii) selection is carried out under low-input and/or organic farming conditions, and (iii) maintenance breeding/seed production is carried out in contrasting pedoclimatic environments to optimize local adaptation [
156,
157,
158].
Most breeding and variety selection programs for organic farming have focused on crop species where modern conventional varieties were found to perform poorly in organic systems, and/or where the combination of traits desired by organic farmers are very different from those delivered by conventional breeding programs (e.g., for wheat, brassicas such as broccoli [Brassica oleracea L. cultivar group italica] and more recently soya [
Glycine max (L.) Merr.]) [
10,
13,
152,
153]. For example, conventional wheat breeding programs have focused on reducing the straw length to minimize lodging risk and increase grain-yield potential in high mineral NPK and pesticide input systems [
13,
153]. In contrast, organic wheat breeding programs have focused on the delivery of varieties with longer stems/straw, high levels of disease resistance, competitiveness against weeds, and the ability to produce a high protein content under low input and/or organic farming conditions [
13,
153].
There is increasing evidence for significant differences in the nutritional composition between modern varieties/hybrids developed for the conventional farming sector and (i) varieties developed in breeding programs for the low-input or organic farming sector, and/or (ii) older and/or traditional varieties selected or preferred by organic farmers. Results from studies with wheat, table grapes (Vitis vinifera L.) and butternut squash (Curcurbita L.) are described, as examples, in separate sections below.
4.1. Wheat
Studies with common wheat (
T. aestivum) in the US and spelt wheat (
T. spelta) in Crete reported that modern, short-straw varieties have lower mineral micronutrient (e.g., Cu, Fe, Se and/or Zn) concentrations in the grain than traditional, long-straw varieties [
13,
40]. In a UK study with common wheat (
T. aestivum), a long-straw variety from a Swiss organic breeding program was also shown to have higher levels of rust resistance, and higher phenolic concentrations than a modern, short-straw variety developed for conventional production [
13]. The study identified strong positive associations between foliar phenolic concentrations, and both rust and Septoria disease severity [
13].
In contrast, in field experiments with spelt wheat (
T. spelta) in Crete, a modern short-straw spelt variety had significantly higher grain phenolic levels than two, traditional long-straw “Urspelt” varieties and an intermediate straw-length variety developed for the organic farming sector [
40].
Overall, the available evidence suggests that longer-straw wheat varieties, favored by organic farmers, have higher levels of mineral micro-nutrients, whereas there is no clear trend for a correlation between straw length and phenolic concentrations.
It is interesting to note that (i) the lower levels of lodging observed with manure, compared with mineral NPK fertilizer inputs, makes it less risky to use longer-straw varieties in organic farming systems, and (ii) the greater competitiveness against weeds and higher grain protein levels in longer straw varieties are a benefit in organic farming systems, which prohibit the use of water-soluble mineral N-fertilizers to increase grain protein levels, and herbicides to reduce weed competition [
13,
40].
4.2. Grapes
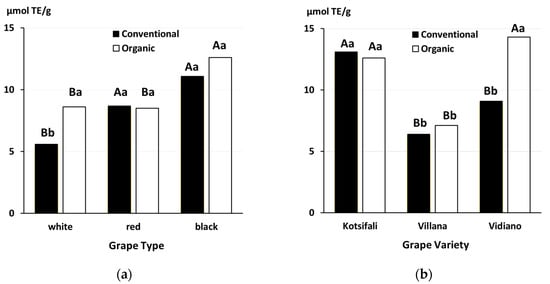
A group of recent studies comparing the nutritional composition of table grapes (which included a comprehensive literature review/meta-analysis of retail surveys in the UK and farm surveys in Crete) also identified significant interactions between variety choice and the production system. These studies compared the effect of production systems (organic versus conventional), growing environments (South Africa versus the Mediterranean) and different grape types/varieties on antioxidant activity, and the phenolic concentrations in table grapes [
12,
19,
60]. They reported grape type (white, red and black) and/or variety as the strongest driver for antioxidant and phenolic levels in grapes and that, overall, organic production was found to result in higher phenolic levels in grapes, compared with conventional production. However, when data from all grape varieties assessed in the UK retail survey were compared, only organic white grapes, but not red and black grapes, were shown to have significantly higher antioxidant capacity compared with their conventional comparators [
12] (
Figure 9a). When the results obtained for individual grape varieties were compared, both the retail and farm survey-based data showed (i) no significant effect of production systems for the majority of varieties, and (ii) higher levels of antioxidants and/or phenolics in organic grapes for some varieties (see
Figure 9b for results from the farm survey). It is important to point out that the reasons for the contrasting effects of production systems on different grape types/varieties are currently unknown [
12,
19,
60].
Figure 9. Effect of organic and conventional production methods on antioxidant activity/capacity (TEAC) in (
a) white, red and black table grape analyses, as part of a UK retail survey, data from [
12] (2-factor ANOVA detected a significant interaction (
p = 0.025) between grape type and production systems) and (
b) grapes of three indigenous, traditional grape varieties analyzed as part of a farm survey in Crete, Greece, data from [
19] (2-factor ANOVA detected a significant interaction (
p = 0.016) between grape variety and production systems). Means with the same capital letter for the same production system and means with the same lower-case letter for the same grape type or variety are not significantly different according to Tukey’s contrasts (
p < 0.005).
This entry is adapted from the peer-reviewed paper 10.3390/agronomy11122494