Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemistry & Molecular Biology
S100A10 (p11, annexin II light chain, calpactin light chain) is a multifunctional protein with a wide range of physiological activity. S100A10 is unique among the S100 family members of proteins since it does not bind to Ca2+, despite its sequence and structural similarity.
- S100A10
- Ca2+-binding
- fibrinolysis
- cancer
- depression
1. Binding Partners of S100A10
In addition to annexin A2, several additional proteins have been shown to interact with S100A10. The physiological significance remains unknown for most of these protein–protein interactions. Additionally, it is unclear if S100A10 is the only protein in the S100 family that will interact with these proteins. A list of published S100A10 protein interactors was generated by summarizing public databases: NCBI (https://www.ncbi.nlm.nih.gov/), BioGRID (https://thebiogrid.org/) and ELIXIR (https://www.ebi.ac.uk/intact/home). Ontology analysis of the protein interactors was performed using DAVID 7.0 [39] (http://david.ncifcrf.gov/). Significant ontology findings with a p value < 0.05 and FDR < 0.1 are summarized in Table 1 and illustrated in Figure 4. Interactions that have been demonstrated to have physiological relevance are presented in Table 2. A complete list of reported interacting proteins is summarized in Table 3.
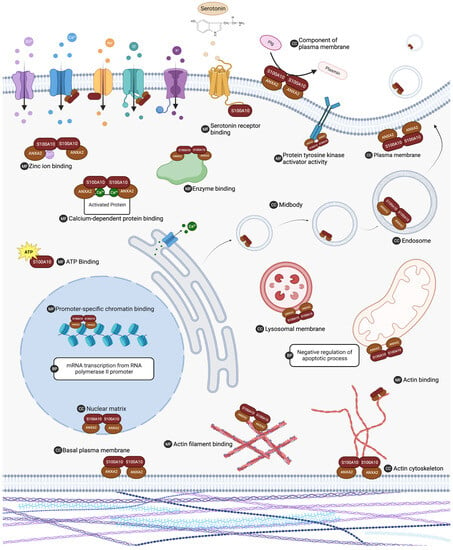
Figure 4. The list of S100A10 interacting proteins was subjected to gene ontology analysis in DAVID 7.0 to characterize putative biological activity. Gene ontology analysis indicated that S100A10 interacting proteins are ubiquitous in the cell and are involved in many vital processes. Biological processes (BP), molecular functions (MF), and cellular component (CC) ontology results with a p value < 0.05 and FDR < 0.1 are summarized. ANXA2 (annexin A2). Created with “Biorender.com (accessed on 6 August 2023)”.
Table 1. Binding partners participating in different S100A10-related functions.
| GO Classification | Ontology Terms | Proteins |
|---|---|---|
| Biological process | Negative regulation of apoptotic process (GO:0043066) | EGFR, ERBB3, ERBB4, FLNA, GDNF, KIF14, MYC, NPM1, PLA2R, PPIF, RAF1, SIN3A, TNF, TP53 |
| Biological Process | mRNA transcription from RNA polymerase II promoter (GO:0042789) | ANXA2, ATF2, HLTF, S100A10, SUPT6H |
| Cellular component | Plasma membrane region (GO:0098590) | EGFR, ERBB3, ERBB4, PLD1 |
| Cellular component | Midbody (GO:0030496) | ANLN, ANXA2, CHMP4B, CHMP4C, CIT, ECT2, KIF14, KIF20A, PRC1 |
| Cellular component | Plasma membrane (GO:0005886) | 5-HTR1B, 5-HTR1D, 5-HTR4, AHNAK, ANTXR1, ANXA2, ASIC-1 channels, CCR10, CD55, CD81, CDK16, CFTR, CHMP4B, CHMP4C, EGFR, ERBB3, ERBB4, ESR1, FLNA, FLOT1, FSHR, GOLGA4, HDLBP, HSPA8, KCNK3, KIF14, LIMA1, MAEA, MAPT, mGluR5, MLKL, MMGT1, MYH9, Myo1c, NaV1.8 sodium channel, OR14I1, OR4N2, PDYN, PIK3R6, PLA2G4C, PLA2R, PLD1, PLG, PRC1, RAF1, S100A10, S100A3, S100A8, SLC8A1, TMEM65, TNF, TNFRSF10A, Transglutaminase, TRPM4, TRPV5, TRPV6, ZGPAT |
| Cellular component | Basal plasma membrane (GO:0009925) | CD81, EGFR, ERBB3, ERBB4, Myo1c |
| Cellular component | Nuclear matrix (GO:0016363) | ANXA2, HLTF, MAEA, PHF5A, S100A10, TP53 |
| Cellular component | Actin cytoskeleton (GO:0015629) | AHNAK, ANG, ANLN, FLNA, LIMA1, MYH9, Myo1c, NEB |
| Cellular component | An integral component of the plasma membrane (GO:0005887) | 5-HTR1B, 5-HTR1D, 5-HTR4, ASIC-1 channels, CCR10, CD81, CFTR, EGFR, ERBB3, ERBB4, FSHR, KCNK3, MAEA, mGluR5, PLA2R, SLC8A1, TNF, TRPM4, TRPV5, TRPV6 |
| Cellular component | Lysosomal membrane (GO:0005765) | AHNAK, ANXA2, ATP6V1E1, CFTR, CHMP4B, CHMP4C, FLOT1, HSPA8, PLD1 |
| Cellular component | Endosome (GO:0005768) | 5-HTR4, ANXA2, ATP6V1E1, CHMP4B, EGFR, FLOT1, FSHR, PLD1 |
| Molecular function | Calcium-dependent protein binding (GO:0048306) | ANXA2, S100A10, S100A3, S100A7, S100A8, S100Z |
| Molecular function | Zinc ion binding (GO:0008270) | BMI1, ESR1, HDAC4, HDAC6, HLTF, NANOS2, NBR1, PHF5A, Q6ZTA4, Q96KR1, S100A3, S100A7, S100A8, SETDB1, TP53, TRIM37, ZCCHC9 |
| Molecular function | Promoter-specific chromatin binding (GO:1990841) | ATF2, BMI1, HDAC4, SETDB1, TP53 |
| Molecular function | ATP binding (GO:0005524) | HSPA8, KIF14, KIF20A, MLKL, MYH9, Myo1c, NARS1, NaV1.8 sodium channel, RAF1, SLFN14 |
| Molecular function | Enzyme binding (GO:0019899) | CFTR, EGFR, ESR1, HDAC6, HSPA8, MAPT, PLA2R, PLG, RAF1, TP53 |
| Molecular function | Actin binding (GO:0003779) | ANG, ANLN, DYRK1A, HDAC6, MAEA, MAPT, MYH9, Myo1c, NEB |
| Molecular function | Calmodulin binding (GO:0005516) | ESR1, MYH9, Myo1c, SLC8A1, TRPM4, TRPV5, TRPV6 |
| Molecular function | Serotonin binding (GO:0051378) | 5-HTR1B, 5-HTR1D, 5-HTR4 |
Table 2. Proteins with known functional interactions with p11.
| Interactor | Protein Name | Uniprot Accession | Cellular Location | Method | References |
|---|---|---|---|---|---|
| 5-HTR1B | 5-hydroxytryptamine receptor 1B | P28222 | Plasma membrane | Two-hybrid | [40,41] |
| 5-HTR1D | 5-hydroxytryptamine receptor 1D | P28221 | Plasma membrane | Two-hybrid | [41] |
| 5-HTR4 | 5-hydroxytryptamine receptor 4 | Q13639 | Plasma membrane | Two-hybrid | [41] |
| AHNAK | Neuroblast differentiation-associated protein AHNAK | Q09666 | Nucleus | AC-MS 1; AC-W 2; Co-crystal structure; Co-fractionation; PL-MS 3 | [42,43,44,45] |
| ANXA2 | Annexin A2 | P07355 | Nucleus; Cytoplasm; Plasma membrane; Extracellular | AC-MS 1; AC-W 2; Co-crystal structure; Co-fractionation; PL-MS 3; Reconstituted complex; Two-hybrid | [43,45,46,47,48,49,50,51,52,53,54,55,56,57] |
| ASIC-1 channels | Acid-sensing ion channel 1 | P78348 | Plasma membrane | Two hybrid | [58] |
| BAD | BCL2-associated agonist of cell death | Q92934 | Cytoplasm | Reconstituted complex; Two-hybrid | [59] |
| Cathepsin B | Cathepsin B | P07858 | Cytoplasm; Plasma membrane; Extracellular | Two hybrid: Reconstituted complex | [60] |
| Cytosolic Phospholipase A2 | Q9UP65 | Cytoplasm: Plasma membrane | Two-hybrid | [61] | |
| NaV1.8 | Sodium channel protein type 8 subunit α | Q9UQD0 | Plasma membrane | Two-hybrid | [62] |
| PCTAIRE-1 | Cyclin-dependent kinase 16 | Q00536 | Cytoplasm | Two-hybrid | [63] |
| Plg | Plasminogen | P00747 | Extracellular | AC-W 2; Surface plasmon resonance | [32,33] |
| S100A7 | Protein S100-A7 | P31151 | Cytoplasm; Extracellular | AC-MS 1 | [64] |
| S100A8 | Protein S100-A8 | P05109 | Plasma membrane; Cytoplasm; Extracellular | AC-MS 1 | [64] |
| TASK-1 | Potassium channel subfamily K member 3 | O14649 | Plasma membrane | Two-hybrid | [65,66] |
| TRPV5 | Transient receptor potential cation channel subfamily V member 5 | Q9NQA5 | Plasma membrane | Reconstituted complex; Two-hybrid | [67] |
| TRPV6 | Transient receptor potential cation channel subfamily V member 6 | Q9H1D0 | Plasma membrane | Reconstituted complex | [67] |
| Transglutaminase | Protein-glutamine-γ-glutamyltransferase K & 2. | P22735; P08587 | Plasma membrane; Cytoplasm | AC-W 2 | [68] |
1—Affinity Capture—Mass Spectrometry, 2—Affinity Capture—Western Blot, 3—Proximity Label—Mass Spectrometry.
To date, over 170 proteins have been proposed to interact with S100A10 and observed through various biochemical approaches. Mass spectrometry was the most commonly used technique to identify S100A10 interactors. This approach involved several techniques, including affinity capture (34% of identified interactors), proximity labeling (13%), and co-fractionation (13%). Two-hybrid yeast screening was the second most popular biochemical approach to identify interactors (13%). Other methods used to identify protein interactors included, but were not limited to, co-crystal structure, affinity capture-western blot, reconstituted complex, co-immunoprecipitation, and fluorescent imaging. It will be interesting to determine the physiological relevance of the predicted binding partners of S100A10 and, more specifically, to identify the binding regions or structural determinants in S100A10 that predict and demonstrate the binding affinity to the various proteins.
2. Regulation of S100A10
The expression and function of S100A10 are tightly regulated by various cytokines, growth factors, nitric oxide (reviewed in [69]), RAS oncogene [70], PML-RAR oncogene [71], and hypoxia-inducible factor (HIF) [72]. Annexin A2 is also an important regulator of S100A10, and S100A10 also regulates the properties of annexin A2, a process called mutualistic symbiosis [1]. MicroRNA (miRNA) and long-non-coding RNA (LncRNA) are important non-protein-coding RNA machinery that make up a large percentage of the genome. They have been demonstrated to function in post-transcriptional and epigenetic regulation. LncRNA and miRNA play a crucial role in regulating development, stress-mediated processes, immune response and tumor progression, metastasis, and chemoresistance [73,74,75,76]. Most evidence of the regulation of S100A10 by miRNA and LncRNA has been obtained from studies in hepatocellular carcinoma (HCC) models. Shan et al. identified that mir-590-5p is decreased in some hepatocellular carcinoma cell lines with a concomitant upregulation of S100A10. They further showed that mir-590-5p regulated S100A10 expression by interacting with the 3′UTR of S100A10 mRNA [77]. Similarly, LINC00174 has been shown to be an oncogenic LncRNA in HCC, and it functions by sponging mir-320, which further upregulates the expression of S100A10, accelerating the proliferation and metastasis of HCC cells [78]. Another lncRNA, KCNMB2-AS1, functions by regulating S100A10 in bladder cancer through direct sponging of miR-374a-3p, which negatively regulates the expression of S100A10 [79].
3. Role of p11 in Fibrinolysis
S100A10 is present at the endothelial cell surface and bound to annexin A2, forming the AIIt complex [2]. This complex has been shown to be one of the key plasminogen-binding receptor proteins on the surface of endothelial cells. The S100A10 subunit binds plasminogen, whereas the annexin A2 subunit anchors the complex to the cell surface [3][4]. The role of AIIt in fibrinolysis was confirmed by studies of the S100A10-knockout mouse, where it was observed that S100A10-null mice displayed increased deposition of fibrin in the vasculature and reduced clearance of experimentally-induced vascular thrombi [5]. Plasminogen (Pg) is an inactive zymogen that is converted to the active serine protease plasmin (Pm) by plasminogen activators, tissue-plasminogen (tPA) activators, and urokinase plasminogen activator (uPA). This process is highly regulated; the principal inhibitor of Pm is α2-antiplasmin (AP), and the principal inhibitor of tPA and uPA is plasminogen activator inhibitor type 1 (PAI-1) [80,81,82,83]. Although tPA and uPA possess low activity, their activity dramatically increases when interacting with Pg receptors. Certain Pg receptors also protect Pg activators and Pm from inactivation. Thus, Pg receptors such as AIIt are the key regulators of Pm generation.
Plasmin is crucial for dissolving fibrin clots in a process known as fibrinolysis. Fibrin blood clot formation begins when a break in a blood vessel allows plasma to contact tissue factors in the extracellular matrix. Initially, factor VII binds to tissue factor and is activated, which participates in the activation of a clotting factor cascade and results in the cleavage of prothrombin into thrombin. This thrombin activates platelets, resulting in the generation of FXa/Va complexes and the generation of sufficient thrombin from prothrombin to convert fibrinogen to fibrin, resulting in the formation of a stable hemostatic plug [84,85]. The fibrinolytic surveillance system is critical in preventing the inappropriate formation of blood clots and in regulating the size of blood clots that form at the site of injury. Fibrinolysis is initiated by the secretion of plasminogen activators (tPA and uPA). The endothelium of blood vessels secretes tPA, which circulates in the blood as a single-chain protein (sct-PA). The kidney secretes uPA, which circulates in an inactive single-chain form (scu-PA), which Pm must convert into the active two-chain form (tcu-PA) [86,87].
Pm degrades the fibrin blood clot, producing D-dimer and other fibrin degradation products. Impaired breakdown of fibrin clots is associated with many diseases, including diabetes, insulin resistance, sepsis, stroke, and the metabolic syndrome [88,89,90,91]. Impaired fibrinolysis also contributes to deep venous thrombosis and pulmonary embolism [92]. Impaired fibrinolysis is a feature of ischemic stroke and is present in both the acute and convalescent phases of the disease [93,94,95].
4. Neurological Functions
Previous reviews of S100A10 have uncovered new insight into the protein’s role in regulating mood-related behavior [97]. S100A10 has been implicated in the pathophysiology of depression and is being examined as a critical modulator of neurological functions [97]. Several G protein-coupled receptors, channels, and transporters have been identified to interact with S100A10, including serotonin 5-HT receptors [40], metabotropic glutamate receptor mGluR5 [98], Na2+ ion channel NaV1.8 [62], acid-sensing channel ASIC-1 [58], K+ channel subfamily K member 3 (TASK-1) [65,66], and transient receptor potential cation Ca2+ channels subfamily TRPV5 and TRPV6 [67]. Global proteomic screens have additionally identified C-C chemokine receptor type 10 (CCR10) [99], flotillin-1 (FLOT1) [44], follicle-stimulating hormone receptor (FSHR) [100], olfactory receptor 4N2 (OR4N2), and 4N1 (OR14l1) [43] as S100A10 interactors.
S100A10 expression is widespread in the brain and spans multiple structures and cell types. Milosevic et al. suggest that the expression is distinct to specific regions when comparing neuronal and nonneuronal cell types [101]. Some identified areas include the hippocampus [45,102], amygdala [102], cerebral cortex [103], anterior cingulate cortex [40], and nucleus accumbens [104]. The expression of S100A10 in the brain has been extensively examined for its role in modulating mood-related behaviors, including major depressive disorder, Parkinson’s disease, and other neuropsychiatric disorders [105,106].
The role of S100A10 in depression and as a regulator of the antidepressant response has been examined in various mouse and human studies. Reduced S100A10 expression in multiple brain regions in depressed individuals implicated S100A10′s role in depression pathology [40,102,107]. Epigenetic studies of S100A10 propose it as a biomarker to predict treatment response and diagnose depressive-like behavior. Expression of S100A10 can be regulated by various factors, such as antidepressants, nitric oxide, growth factors, and dexamethasone [108,109]. While S100A10 is reduced in brain tissue from depressed patients, S100A10 expression increases in the peripheral blood. One study comparing S100A10 expression levels by quantitative PCR found that compared to healthy patients, depressed and high-stressed individuals studied had higher expression of S100A10 in their blood cells [110].
The S100A10-knock-out mouse models have been instrumental in studying the function of S100A10 in depression, mood disorders, and anxiety. In one of the first hallmark studies, Svenningson et al. showed that S100A10-depleted mice demonstrate depression-like behaviors, and S100A10 expression is increased in rodent brains after anti-depressant therapy. S100A10 mediates its function in depression via interaction with the serotonin 1B receptor [5-hydroxytrptamin (5HT1B) receptor] [40]. More recently, Seo et al. observed that S100A10 is a key protein that modulates chronic stress-induced depression in rodent models, and anti-depressants reverse this phenotype by upregulating S100A10 in the prelimbic cortex (PrL) [109]. Furthermore, they showed that S100A10 in ependymal cells regulates depressive states in chronic stress by modulating the flow of cerebrospinal fluid (CSF) by maintaining the planar cell polarity in ependymal cells [109].
5. Inflammation and Wound Healing
The plasminogen activation system and S100A10 play an integral role in the inflammatory process. S100A10 functions on the plasma membrane and in the extracellular space, correlated with the immune response and regulation of immune activities [111]. The Ca2+ binding activity of S100A proteins plays an important role during the inflammatory process by regulating different molecules and inflammatory signaling pathways that lead to inflammation [112,113]. S100 isoforms can contribute to the immune response as pro-inflammatory stimulators, chemo-attractants, and antimicrobial peptides [111].
During inflammation and wound healing processes, extracellular matrix (ECM) degradation is a vital step, and the S100A10 dimer component of Allt is a crucial molecule in the process [112]. Pg binds to the S100A10 dimer of Allt, and plasminogen activators tPA and uPA mediate its activation into Pm, which can initiate downstream proteolytic cascades associated with the wound repair process [32,34,60,114,115,116]. This S100A10-mediated protease activation is utilized by highly motile cells, such as metastatic cancer cells and macrophages, to facilitate its migration through the ECM [114,116,117,118,119,120,121,122,123].
Macrophages play a critical role in the pathogenic inflammatory response by initiating, maintaining, and resolving inflammation processes [119,124]. Cell-surface generation of plasmin is required for macrophage recruitment [125]. This recruitment is partly mediated through the plasmin-dependent activation of matrix metallopeptidase 9 (MMP-9) [119,125]. Activation of plasmin occurs through four plasminogen receptors found on the cell surface, including S100A10 [32,34,35,126], α-enolase [127], Plg-RKT [128], and histone H2B [129].
We have previously demonstrated a direct involvement of S100A10 in response to inflammatory stimuli to recruit macrophages [119]. P11 was up-regulated in macrophages that were activated by inflammatory mediators. Thioglycollate-stimulated peritoneal macrophages had higher S100A10 and annexin A2 protein levels when compared to resident peritoneal macrophages. In S100A10-/- mice, macrophage migration into the peritoneal cavity across the peritoneal membrane was decreased. S100A10 and the other carboxyl-terminal Pg receptors contribute to Pm generation in macrophages, which allows macrophages to play a role in directly facilitating the proteolysis of the basement membrane, hydrolyzing ECM proteins, and activating MMP-9 during inflammatory responses [119].
S100A10 can also activate human and murine macrophages directly through the toll-like receptor 4 (TLR4) pathway [130,131]. Cell surface Pm generated by Allt can trigger the phosphorylation of PKC signaling molecules, which can activate mitogen-activated protein kinase (MAPK), TLR4, and NFκB signaling pathways [130]. Allt disassembly after Allt phosphorylation can activate the CD11b-dependent integrin-linked kinase (ILK) pathway [132], which, together with Pm, can induce NFκB nuclear translocation and promote pro-inflammatory factor production [26,130,133]. The release of these pro-inflammatory cytokines, including IL-1, IL-6, and TNFα, can facilitate immune-escape mechanisms by evading immunosurveillance and mitigating T cell cytotoxicity [26,130,133]. Allt-driven cytokine production is inhibited by TLR-4 knockdown [130], which suggests that TLR-4 is important for Allt-mediated inflammation.
More recently, S100A10 has been examined for its role in the inflammatory condition of COVID-19 patients. The S100 family of proteins has been suggested to be able to direct more monocytes and neutrophils to the target site of COVID-19 patients by controlling the cytokine release syndrome [113]. A study comparing NK and NK-T cell subsets between COVID-19 patients and healthy individuals identified S100A10 as a marker for COVID-19-derived NK-T cells [134]. Disease trajectory transcriptomic models of COVID-19 severity from peripheral mononuclear cells found that genes encoding Ca2+-binding proteins play important roles in regulating inflammatory pathways; however, p11 was not indicated as one of the S100 proteins involved [135]. Another study looking at peripheral blood samples from COVID-19 cases observed an upregulation of S100A10, S100A4, and S100A9 mRNA [113].
6. Exocytosis and Trafficking
The AIIt complex has long been implicated at the plasma membrane to play a role in early and late secretory events and membrane-cytoskeleton lineage [47,136,137]. At the cell membrane, the AIIt complex was found localized to the contact site of the plasma membrane/secretory granule, suggesting it played a role in the exocytotic process of the cell [138,139,140,141]. While the crosslinking activity of the AIIt complex is necessary for efficient, regulated exocytosis, it is not an obligatory component [137]. The exocytotic machinery of the Allt complex is Ca2+ sensitive [142] and is more efficient than monomeric annexin protein in regulating Ca2+-dependent exocytosis [138,143,144]. These features and functions of the AIIt complex have played an important role in the pathogenesis of many viruses, such as HBV, HPV, and HIV-1 [145,146,147,148,149]. The AIIt complex has been shown to facilitate the exocytosis of von Willebrand factor (vWF) in vascular endothelial cells [150,151], the release of the bluetongue virus (BTV) in BHK21 cells [152,153], and the human papillomavirus type 16 in human keratinocytes [154]. Finally, recent studies by Bai et al. have shown that the AIIt complex promotes hepatitis B virus (HBV) exocytosis in the trophoblasts by recruiting VAMP2 and SNAP25 for membrane fusion events [145].
7. Autophagy and Metabolism
Cancer cells require increased metabolism to maintain sustained proliferative growth and spread. S100A10 can respond to a variety of signals to maintain and accelerate cancer cell metabolism. Members of the S100 family of proteins are involved in a number of metabolic functions, including redox, energy, and sugar metabolism [15]. S100A10 has been shown to increase the malignant growth of cancer cells by activating the mTOR signaling pathway in osteosarcoma [155], gastric cancer [156], pancreatic ductal adenocarcinoma (PDAC) [157,158], and HCC [159,160]. Li et al. reported that S100A10, through its interaction with annexin A2, accelerated tumor glycolysis and lactate production and contributed to the switch from oxidative phosphorylation to aerobic glycolysis [156]. Glucose consumption was significantly increased in S100A10-overespressing cells and reduced in S100A10-knockout cells. This overexpression of S100A10 also reduced the amount of intracellular ATP production, indicating the important role of S100A10 in facilitating glycolysis. By modulating the Src/annexin A2/AKT/mTOR signaling pathway, S100A10 could promote pro-tumor aerobic glycolysis, suppress cell apoptosis, and maintain cell proliferation.
S100A10 knockdown was also shown to affect the malignant growth of osteosarcoma cells by modulating glycolysis [155]. Knockdown of S100A10 in the osteosarcoma cell line inhibited proliferation, migration, and invasion and induced apoptosis via the AKT/mTOR pathway by modulating glycolysis. S100A10 played a critical role in HCC progression through the epithelial-mesenchymal transition by upregulating the epidermal growth factor receptor and AKT/ERK signaling pathways. Furthermore, Lin et al. showed that S100A10 could activate LAMB3 through the JNK pathway in PANC-1 cells [157]. By activating the JNK/LAMB3-LAMC2 axis, S100A10 was able to promote PDAC cell proliferation, migration, and adhesion.
This entry is adapted from the peer-reviewed paper 10.3390/biom13101450
References
- Alamelu Bharadwaj; Emma Kempster; David Morton Waisman; The Annexin A2/S100A10 Complex: The Mutualistic Symbiosis of Two Distinct Proteins. Biomol. 2021, 11, 1849, .
- Geetha Kassam; Kyu-Sil Choi; Jaspinder Ghuman; Hyoung-Min Kang; Sandra L. Fitzpatrick; Tracy Zackson; Saul Zackson; Mikayo Toba; Aya Shinomiya; David M. Waisman; et al. The Role of Annexin II Tetramer in the Activation of Plasminogen. J. Biol. Chem. 1998, 273, 4790-4799, .
- Geetha Kassam; Bich-Hang Le; Kyu-Sil Choi; Hyoung-Min Kang; Sandra L. Fitzpatrick; Peter Louie; David M. Waisman; The p11 Subunit of the Annexin II Tetramer Plays a Key Role in the Stimulation of t-PA-Dependent Plasminogen Activation. Biochem. 1998, 37, 16958-16966, .
- Travis J. MacLeod; Mijung Kwon; Nolan R. Filipenko; David M. Waisman; Phospholipid-associated Annexin A2-S100A10 Heterotetramer and Its Subunits. J. Biol. Chem. 2003, 278, 25577-25584, .
- Alexi P. Surette; Patricia A. Madureira; Kyle D. Phipps; Victoria A. Miller; Per Svenningsson; David M. Waisman; Regulation of fibrinolysis by S100A10 in vivo. Blood 2011, 118, 3172-3181, .
This entry is offline, you can click here to edit this entry!