“Quantum biology” (QB) is a promising theoretical approach addressing questions about how living systems are able to unfold dynamics that cannot be solved on a chemical basis or seem to violate some fundamental laws (e.g., thermodynamic yield, morphogenesis, adaptation, autopoiesis, memory, teleology, biosemiotics). Current “quantum” approaches in biology are still very basic and “corpuscular”, as these rely on a semi-classical and approximated view. We review important considerations of theory and experiments of the recent past in the field of condensed matter, water, physics of living systems, and biochemistry to join them by creating a consistent picture applicable for life sciences. Within quantum field theory (QFT), the field (also in the matter field) has the primacy whereby the particle, or “quantum”, is a derivative of it.
1. Introduction
One of the main reasons why living matter challenges the descriptive capacity of the sciences so much is due to the fact that dealing with living systems essentially means dealing with open processes, flows, and relationships. Living systems are never isolated, are self-contained, and thus elude us as observable objects. A living system is not a finite “body” but is always in flux, existing as a process, as “ordered responses” along a thermodynamic history of couplings and adaptations. This fact is difficult to “handle” with the prevailing scientific jargon, that is, when trying to reduce a living entity to the physical realm of how and why living dynamics emerge. We must come to terms with the fact that the language of the physical sciences and the commonly accepted condensed matter view [
1] is only appropriate for describing objects and observables that exist in themselves
2. Shifting from QM to QFT
2.1. Order, Symmetry Breaking, Dissipation, and Coherence
Today, “quantum” approaches are becoming more common in biology [
6], simply because some properties and phenomena cannot be described classically. However, most of these approaches still rely on the semi-classical theoretical framework of quantum mechanics (QM), which is not only an incomplete theory [
7,
8] but is also unable to describe the phase transitions that constitute living matter.
Biological matter has the special property of being composed of long-range correlated components. One might be tempted to think that the components (types of matter quanta such as molecules, ions, electric charges, atoms, and electromagnetic fields) in soft matter (alive) are identical to the inanimate state (not alive) and that “something mystical” causes them to function in a “special way” to yield the alive state. As will be shown, this vitalist-tinged perspective suffers from a postulate that holds only if one remains faithful to the classical or semi-classical view of matter (or what is considered “physical events”), where the ground level (minimum energy) of a system (even of a single atom) is uniquely determinable (for classical physics, CP) or uncertain (within the Planck limit) but nonetheless unambiguous (for quantum mechanics, QM) [
9,
10]. In the conceptual framework of QM and in accordance with the fundamental theorem of von Neumann [
11], a unique ground state (vacuum) of the ensemble of interacting molecules interacting by forces does indeed exist. Therefore, a unique state (or phase) is admitted whose phase transitions are not describable [
10].
Since a biological system is constantly connected to the external environment (open), its isolation means the elimination of its functionality, its destruction that ultimately will be lethal. As addressed by Vitiello [
3,
17], when studying an open system (say, “system
α”), scholars proceed to “close” it, also considering the environment in which it is immersed. It can refer to the latter as “system
β”. Doing so assures that the flow equilibria of matter, energy, and phase correlations between the system
α and the environment is still describable through thermodynamics of closed systems. With respect to energy exchange, the energy output of
α, E(
α), must be equal to the energy input of
β, E(
β), and vice versa. In either case, it must be E(
α) − E(
β) = 0. Therefore, the set (
α,
β) of systems
α and
β behaves like a “closed” system for which there is no energy flow, neither input nor output.
Given the energy balance (and the balance of flows of all other quantities exchanged between
α and
β), system
β behaves like a copy of system
α, in the sense that it behaves the same as system
α in terms of flows, provided they are reversed: what is an input for
α is an output for
β and vice versa. Thus, reversing the direction of flow is equivalent to exchanging
α with
β or vice versa. Since technically the flow direction is reversed by changing the sign of the time variable, it can say that
β behaves like the copy of
α for which the time arrow has been reversed (
β is the time-reversed copy of
α). In essence,
β is the system that describes the environment in terms of the balance of
α’s energy flow, while it is also the mirror image of
α in the mirror of time (
β is the time mirror image of
α): Vitiello effectively expressed this fact by writing that
β is the “double” of
α [
17]. Thus, in the case of open systems (such as organisms or brains), we need to consider their doubles, and this offers some interesting insights. The formation of each configuration (
α,
β) is enabled by the breaking of symmetry induced by the external stimuli, so that:
- (i)
-
The manifold of possible configurations is made possible by the existence of the numerous (infinite) possible ground states in the scenario offered by QFT;
- (ii)
-
The coexistence of these multiple configurations is given by the fact that these ground states are orthogonal to each other;
- (iii)
-
Their temporal sequence is given by the dissipative dynamics, i.e., by their thermodynamic history (by all possible pairs of values of N(α) and N(β) obeying the relations N(α) − N(β) = 0);
- (iv)
-
The succession of the states of the system is indeed a time-dependent thermodynamical (dissipative) history along which the following states depends on the previous ones in a deterministic, but a priori unpredictable, way because of the presence of its thermodynamical “double” (environment, inclusive of every possible quality and kind of stimuli).
Based on the theoretical foundations of quantum field theory (QFT), the structural and functional order of a biological system (which is neither stochastic nor predeterminable) results in a condition that arises dynamically as a consequence of symmetry breaking over a wide range of degrees of freedom of the system [
10,
18].
In spontaneous symmetry breaking (SSB), the symmetric properties of the microscopic dynamics imply the presence of fields which, distributed throughout the system, cause the formation of ordered configurations of the elementary components.
The quanta associated with such dynamics (i.e., SSB) are called Nambu-Goldstone (NG) quanta (or particles or modes) [
3,
19,
20,
21], and include the phonons in crystals, the magnons in ferromagnets, the polarons in ferroelectrics, etc. Quanta of NG and responsible for the space–time arrangements observed in such systems, which, depending on the case (geometric, magnetic, rotational, electric, etc.), carry the specific correlations that generate the observed order. Even at very different scales, as in cosmology [
9,
22], or where non-trivial forms of order occur, as in biological systems (a very special case of condensed matter), the breaking of several kinds of symmetry underlies the appearance of order parameters [
3,
10,
19,
23,
24].
The NG quanta are bosons, i.e., many of them can occupy the same physical state [
25]. Under ideal conditions (without “edge effects”) they have zero mass, and in their low-momentum state they do not contribute to the energy of the ground state (vacuum): the emergence of order is therefore a condensation effect of the NG bosons in the vacuum (ground state). The NG quanta are the mediators of correlation and are structural elements.
In a liquid or gas, the atoms (or molecules) are in a state where their positions in space can be shifted without changing the macrostate. A shift in any direction of space therefore produces a configuration equivalent with identical free energy (and entropic content). In practice, a gas or liquid are symmetric systems with respect to the degrees of freedom of spatial displacement.
Therefore, when the system is open and capable of dissipating a certain amount (i.e., entropy, latent heat of condensation), a phase transition from a disordered state to a more ordered state occurs spontaneously [
1,
10]. This new state, the vacuum level, is energetically lower than the previous one. This energy gap (in electron volts, eV) characterizes its thermodynamic stability. The translational symmetry is broken, and it is no longer possible to move along another direction of the system experiencing the same potential landscapes.
This suggests that (i) continuous symmetry breaking as a result of the guided pathways in biotic systems that seeks to maintain a viable homeostasis leads to a lack of symmetry and (ii) is the consequence of quantum dynamics in which the components of the matter field phase their oscillations and create a correlation that manifests itself as a field consisting of a discrete set of bosons (in this case phonons) [
17]. This correlation field is indeed a classical field (the order parameter), since it is insensitive to quantum fluctuations and causes the macroscopic (classical) stability of large quantum systems (such as a crystal) [
10].
However, phonons cannot be “detached” from the system, i.e., there are no freely propagating phonons outside a crystal because they exist only as long as the crystal exists. This implies that the quanta of NG are identified with the function of the specific order for which they are responsible and thus also express the functionality, the mode of being of the system. Without them, the system is “another system” with completely different physical properties (functionality and therefore structure).
2.2. Condensed Matter from a Field Perspective: Water, the Matrix of Life
The principles of SSB described above—applied to condensed matter such as liquid water—allow a fully consistent picture to emerge accounting for phenomena occurring in both plain water and living matter.
Once a critical threshold density (below a critical temperature) is reached, SSB induces condensation of matter in coherently coupled domains [
10]. For water, such thresholds are reached at 0.32 g/cm
3 and 373.15 K (at 1 atmosphere pressure) [
31].
Experimental evidence [
32,
33,
34,
35] and descriptive approaches [
36] have demonstrated that liquid water must be a two-phase system. According to QED [
1], part of the molecules in such a system (whose abundance is in “inverse proportion” to the temperature) are organized in collective, coherent vibrational domains corresponding approximately to the size of a wavelength of the coupled electromagnetic mode. Their energy range corresponds to the spectral distance between the two energy levels at which the electron of each water molecule oscillates. Such regions are called coherence domains (CDs), and, in liquid water, where coherence is established between the sp
3 and 5d levels, their size is ideally the wavelength of the corresponding electromagnetic mode (photon), about 100 nm (neglecting thermal fluctuations, which correspond to a frequency of the field in vacuum of 3 PHz, which, during the coupling with the matter field given by the resonating molecules, will be renormalized down to the IR range, about 6 × 10
13 Hz).
The new ground state (known as a “spontaneous superradiant phase transition”) associated with coherently oscillating molecules is lower than that of incoherent, isolated molecules. The difference of this energy gap expresses the thermodynamic stability of the coherent state with respect to decohesion agents acting from outside (such as temperature, photons, fields in general, and mechanical forces). If the excitations are small enough (smaller than the energy gap), the CD receives them as a whole; if they are larger, one or more oscillators are released from coherence and “spilled back” into the normal (incoherent) part [
1,
37].
If the fluid is confined by hydrophilic surfaces, the coherent fraction is stabilized, and possibly other degrees of coherence (such as dipole rotation) can be set. This is the typical condition in living matter as described further below [
38,
39].
As water CDs are self-generated total-reflectance cavities for the EM field, the presence of an externally supplied cavity and pump (as is the case in laser physics) is not required. Indeed, the self-trapped electromagnetic field cannot be externally irradiated as it an intrinsic phenomenon related to the coupling of molecules and as such implies total internal reflection [
37,
40]. Interestingly, almost four decades ago, Persinger [
41] noted that the effects of therapeutically active substances are due to their electromagnetic oscillations rather than due to the biochemical properties; that is, a drug’s electromagnetic oscillations already induce an effect before it is ingested.
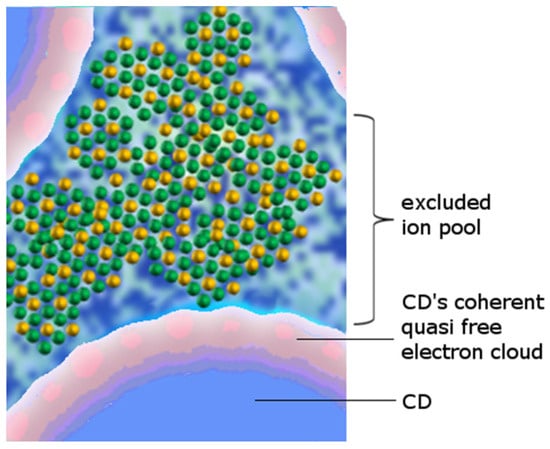
Each molecule participating in the coherent dynamics is endowed with an electron that spends about 10% of its time at the 5d energy level, 12.07 eV above the sp3 hybrid orbital (ignoring collective storable electron excitations). This level is at most 0.53 eV below the ionization threshold (Ith), which for water is 12.60 eV.
These quasi-free electrons can also escape from a CD through the quantum tunneling and in doing so become available to suitable reaction partners (such as the oxygen molecule, non-coherent water molecules, or other species) or are extracted by small excitations [
42]. Tunneling of electrons from the CD leaves behind ionized water molecules. These ions no longer participate in the coherent oscillation and therefore pass to the non-coherent fraction—exactly as described in the Landau model of liquid helium [
37].
The extracted electron on the other hand could be captured by an oxygen molecule dissolved in water, creating a negative ion. The resulting ion pair, after some chemical steps [
43], eventually leads to the formation of a proton and reactive oxygen species such as a hydroxyl. Thus, the CD is an electron donor and then a chemical reducer, which, together with the non-coherent moiety, forms a “redox pile” [
4,
43,
44].
Because water CDs are easily excitable, they can collect many small external excitations (high-entropy energy) and generate unique coherent vortices whose energy is the sum of all small, collected energy excitations but whose entropy is low (high-degree, low-entropy energy) [
48].
In turn, to generate coherence between the CDs, it is necessary to make them oscillate, which means that in one part of the oscillatory cycle the CDs should be able to release energy to the outside, and in the other part of the oscillatory cycle they should recover this energy from external excitations [
4,
16,
48].
Thus, if external non-aqueous molecules present in the fluid contain in their own spectrum at least one frequency close to the vibrational frequency of the water CD (resonance), then these molecules could participate in the coherent dynamics of the fluid and would be subject to dynamics that we now outline [
42].
2.3. Living Phase of Matter: Time-Dependent Order and Morphogenesis
The molecules belonging to the coherent fraction form a set in which the phase is well defined: their common wave function is defined by eigenstates of the phase (
φ), complementing the observable “number” (
N): the uncertainty relation (Δ) expressed in natural units (with
h/2π =
c =
kB = 1, where ”
h” is Planck’s constant, ”
c” is the speed of light in vacuum, and ”
kB” is Boltzmann’s constant), results in the “fundamental uncertainty relationship” expressed as Δ
φΔ
N ≥ ½ [
1].
In a perfectly coherent state, the number of oscillators is completely indeterminate, while the phase, the wave-like aspect of the field, is precisely defined. This means that in a coherent state, the individuality of the oscillators loses physical meaning, since the matter field of uncountable quanta coupled to an electromagnetic field whose massless part defines itself as a quasiparticle in the coherent phase [
53,
54,
55,
56,
57] is the only definable object [
1,
7].
This is equivalent to minimizing the uncertainty of the phase (Δφ) and thus maximizing the uncertainty of the number of quanta (ΔN). In order for this to happen, the system tends to have a large number (N) of quanta because ΔN ≤ N, with a continuous transition, i.e., an exchange between the coherent phase and the external environment. Since the coherent state is thermodynamically more stable (lower vacuum level than the disordered state), coherent systems tend to share their oscillations with other systems, with which they can resonate as to increase N. This is a fundamental feature for understanding that living systems are de facto (super)coherent systems. The open flow of matter and energy quanta is thus possible as they share a common vibrational phase with everything capable of doing so.
Since phase correlations are nonlocal correlations that do not imply an exchange of kinetic energy [
58,
59], the definition of the term “ecosystem” in the theoretical framework of QED is appropriate. In this context, the term “ecosystem” is considered a region of spacetime, in which living systems share an oscillation phase of certain electromagnetic modes that cover very large spatial domains (extending over kilometers and more) at long wavelengths [
15,
60].
Quantum-mechanically, the density of water does not significantly alter with or without a solute. When the solute concentration is high, the concentration of water coherence domains is low. As the concentration of the solute decreases, the concentration of water coherence domains increases [
67] Thus, by modulating the ionic concentration, the cell is capable of regulating the degree of water CDs.
The size of coherent domains is never smaller than the maximum distance between any surface (membrane, molecular chain, etc.) and another [
42]. Such supramolecular organization of water molecules into CDs has two major important implications: (i) the topology of the quasi-free electrons circulating around CDs cannot be tridimensional [
69,
70] (this does not exclude that CDs occupy 3D volumes and can exist in bulk pure water [
32], in contrast to Sen et al. [
69]) and (ii) no supercoherence can be observed in pure liquid water [
42,
71]. In order to self-organize on further levels, CDs require the presence of other species with which to resonate or some surfaces such as nano-particles (e.g., ions or bubbles) and/or hydrophilic walls [
4,
67,
72]. Only then will the entire water matrix in living matter be able to experience multimodal coherence, termed supercoherence, which allows for general “multiplexed” phase correlation throughout the system—be it a single cell or a multicellular organism [
4,
62].
One of the most difficult aspects of molecular biology to understand has always been the fact that biochemical processes occur with incredible efficiency, with an extraordinary degree of precision and timing [
43,
64]. This capacity for biochemical activity also allows living matter to express one of its most challenging properties that cannot be simulated in vitro, namely the ability to perform chemical reaction cycles [
64] that are perfectly selected from a large number of educts that are present together in the cellular landscape—a good example is the Krebs or Calvin cycle. How can this be explained?
In trying to summarize this very illustrative aspect (for details, see [
2,
4] it should be remembered that any chemical species is first and foremost a physical species, i.e., an oscillator characterized by its own precise proper frequencies (modes).
In the coherent phase, there are regions where a background field oscillation acts as a director and coordinator of the molecular encounters. Inside the CD, however, and due to the high dielectric constant 𝜀𝑐𝑜ℎ~160, fields are strongly shielded.
Once the reactive species have been brought together and activated, the water CD catalyzes the biochemical reaction (typically an oxidation–reduction reaction) by releasing electrons (which in the coherent phase occupy states close to the ionization threshold and are therefore readily released at charges below 0.4 eV [
72] or by emitting field quanta I the form of biophotons [
61,
77,
78,
79].
These reaction paths (guided by the sequence of vibrational modes of the coherent fraction) are deterministic and by no means random. On the other hand, they are also unpredictable, since they are determined by (i) the preceding overall thermodynamic history at a given time, (ii) the coherent configuration of the CD at that time, and (iii) the current boundary conditions according to the available reagents and their concentrations, ambient fields, phase correlations, macroscopic thermodynamic variables, etc.
2.4. A Sensible Case: Ions and Ultraweak Fields
Although dissolving salt in water is a common and everyday occurrence, it holds very deep mysteries: How can solvation begin? How can a small molecule like water, which is about 3 Å in size and whose dipole amplitude is no more than 0.38 Å, be able to detach ions from an ionic crystal whose stability is determined by a considerable bond strength (bond strength for most common salts is approx. 5 eV)? Although it is known that the dielectric constant of water at room temperature is about 80, the question arises of why calculations based on the molecular properties of H2O using the Langevin equation give a value of about 13. And why does the solvent power increase with an increase in temperature while at the same time the dielectric constant decreases? These questions cannot be answered consistently from either the CP or the QM points of view and can only be explained by considering coherence in liquid water. The empirically measured high value of the dielectric constant in liquid water (ε = 80 at 300 K) applies only macroscopically to the bulk liquid. This could justify the steady state of the already dissolved ions, which provides a rationale for their separation from each other. But it does not realistically support any process involving the onset of dissolution (its dynamics).
The first transition of the electric dipole of water occurs at about 7.4 eV, i.e., in the full UV spectrum, and the oscillator strength (f) of such dipolar transitions is very low (f∼0.05) [
84]. Thus, the water molecule is an electric object that is rather rigid, which contradicts the high polarizability required by the pair potentials used in the second-order perturbation theory. This means that the only electrostatic effects water can produce are related to its modest electric dipole and are too tiny to keep oppositely charged ions apart and separated. The attractive Coulomb potential between charged ions in the crystal lattice of the salt can be overcome only if a lower-energy state is accessible; a component is needed that must be able to balance the large enthalpy of ionic bonding, which reaches values in the range of 5–7 eV on average in common salts. The dissolution of ions can take place only if the dielectric constant of water is lower than the known value of
εbulk = 80 (in fact, within the QED picture, the incoherent phase has a value of
εinc = 13, whereas the coherent phase of water with
εcoh~160 is quite high. It can be concluded that the ions oscillate in phase with each other and in a cross distribution of ions and counterions. Thus, the energetic gain of the system Δ, which allows it to overcome the strong stability given by the Coulomb ionic bonds, is associated with the creation of new dynamical structures that are ordered and coherent and embedded in the noncoherent phase of the solvent.
The energy gap depends only on the temperature and the value of the dielectric constant of the incoherent part of the liquid (which in turn also depends on the temperature), and this energy gap corresponds to values of the order of a few eV. Each ion coordinates around itself a 3D plasmatic cage of counterions (with opposite charge). The constraint of coherence (i.e., all ions must vibrate in the same phase and at the same frequency as their counterparts) means that the counterion cages must be identical to avoid collisions (collision implies that these cages to be torn apart), with the concomitant loss of coherence. This comes at an energetic cost and therefore does not occur within certain concentration and temperature thresholds. Furthermore, once temperature increases, the solubility in water increases with the rise in the incoherent fraction of water, increasing the space available for ion dissolution (Figure 5).
Figure 5. Sketch showing the state of dissolved ions in liquid water according to QED theory. The coherent, non-ideal Debye–Hückel plasmas located in the non-coherent fraction of water explain how such a stable system—like the ionic crystal—can dissolve and why with the increase in temperature increases the solubility of many ionic species.
On top of that, coherence prevents ions from being subjected to thermal collisions: it is clear that ion ensembles are noiseless inside. The only noise is that coming from the collision of the collective plasmas (cages) with the incoherent molecules of the solvent. Such collisions do not produce any recoil in the ions, since they respond as a whole and all move in unison.
The fact that the ions cannot be subjected to collisions, as presented in the common “Brownian picture”, is confirmed by the fact that, if it were true, a trivial solution of NaCl in water would emit radiation (bremsstrahlung) as ions are accelerated charged bodies. If one assumes that each collision releases an energy of about 10
−17 eV, the solution should release about 10
−5 eV/s per ion, and a standard electrolytic solution at room temperature should release all of its energy (and freeze in the process) within 42 min [
85].
2.5. Hormesis: The Stimulus–Response Relationship
Hormesis (a creation from ancient Greek ὁρμέειν (
hormáein), meaning “to set in motion, drive, urge”—a term coined by Southam and Ehrlich [
94]—describes a favorable biological response to low doses of toxins and other stressors, which activate an adaptive stress response that raise the resistance of the organism against high doses of the same agent [
95]. Thus, hormesis refers to the stimulatory, non-linear effects at low concentrations of otherwise toxic substances. In biological systems, any physical, chemical, or biological agent can be considered stimulatory when administered in much lower concentrations than when administered in undiluted, harmful concentrations [
96]. While hormetic effects of ionizing radiation have been known for quite some time [
97,
98], only recently has the biomedical community started to exploit hormesis for public health, therapeutic, and commercial benefits [
99].
Whether hormesis is a “generalizable” concept valid throughout the animated world is the subject of ongoing discussions. So far, this process addresses (1) evolutionary aspects, (2) how widespread it is among biota, (3) where it is integrated in physiological response systems, and (4) how environmental physicochemical stressors trigger it [
100]. While an extensive literature study showed that 20–65% of the chemical agents investigated display biphasic response patterns [
100], the corresponding observations of a purely physical agent such as non-ionizing radiation are still regarded as non-existent. Thus, in accordance with the proposed harmonization of the numerous expressions describing the biphasic dose–response pattern (see [
101]), it would be suitable to group electromagnetically induced hormesis within the category of “radiation hormesis”, even though the radiation involved is non-ionizing. Even if this extension is considered, however, a distinct difference must be taken into account, which differs from radiation hormesis.
The word “law” in this context indicates that it has proven to be a good fit to experimental data in physiology and psychology [
107,
108] and in botany [
109]. The WBL states that the physiological or psychological response of an organism is a linear function of the logarithm of the stimulus magnitude. Mathematically, the WBL can be written as
𝑅 ~ 𝑙𝑜𝑔(𝑆)+𝑀, where
R is the response (e.g., the number of nerve impulses emitted by a sensory organ over time),
S is the stimulus magnitude (e.g., the intensity of light to which an eye is exposed), and M being a constant. The frequent occurrence of the WBL suggests that it may be a fundamental physical or chemical property of soft matter.
Thus, the cyclotronic ion resonance phenomenon testifies to many important facts. First, it shows that the passage of ionic species across cell membranes (where the electric potential is typically 70–100 mV) occurs through a subtle frequency modulation of the oscillating magnetic fields in the cell medium. Second, it shows that the passage through the membrane follows a particle-wave duality, in that the wave-function determines the selectivity of the channel. Third, it tells us that the functioning of biological systems is much more sensitive to “subtle” influences (to “minimal stimuli” in fact) than to large, violent stimuli that tend to “stiffen” the sensitive response dynamics of the living state. Fourth, it can understand how biological systems, which are essentially composed of carbon, nitrogen, hydrogen, oxygen, and a number of other elements, may be able to detect and sense very low-intensity fields and use them for their autopoietic dynamics. There are numerous cases that show the sensitivity to constant magnetic fields (DC): for example, about 1 nT in bees and 10 nT in birds [
51,
52,
111].
3. Summary
The existence of complementary observables highlights an essential uncertainty and implies that at least two quantities are not simultaneously observable. When attempting to explain coherence in macroscopic systems, such as living organisms using QED, the quantum phase operator,
𝛩̂ —conjugated to the number operator,
𝑁̂
(the smallest entity in a given system at a given scale [
28])—emerges as a crucial variable. Given the thermodynamic openness of a coherent system (like a living one), intrinsic fluctuations Δ
N of its quanta are in force: [
71] Δ
N·Δ
φ ≥ ½ (in natural units).
The crucial point here is that such an uncertainty relation is scale-invariant as only pure numbers without units are involved. It follows that there is no fundamental difference between observer and observed system between the quantum and classical system. The classical way of thinking results when the total number of quanta can be determined with certainty, i.e., when Δ
N = 0. In this case, the phase φ is a random variable that varies randomly (incoherent) from one quantum to another. However, if the total number of quanta is not known with certainty, the quantum phase can take a well-defined value (Δ
φ → 0) if Δ
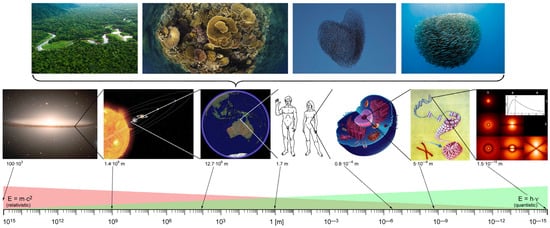
N → +∞. This situation describes a coherent quantum regime that is repeatedly observable at the macroscopic level such as in ferromagnetism, ferroelectricity, superconductivity, and superfluidity. This type of quantum coherence is typical for systems with high density and thus also occurs in water in the liquid state. The essential point here is that a “quantum” must not necessarily be an elementary particle, but can be an ion, a molecule, a protein, a cell or even a bird or fish (
Figure 9) and at a given scale can be considered as a whole entity, thus cannot be broken down into smaller parts without losing that which led to the essence (behavior). Hence, thinning out via dilution—such as the vapor phase of water, for example—(Δ
N → 0), implies that coherence will be lost ([
71]). It is this kind of quantum coherence in high-density situations that makes it particularly applicable to a living organism [
121].
Figure 9. Collective many-body coherent behavior as an emergent property seen on each of the presented hierarchical levels. The “fundamental uncertainty relationship“ (Δ
N·Δ
φ ≥ ½) becomes especially manifest in the biologic realm and is most visible in swarm of birds and fishes as well as coherent coupling in larger biomes, e.g., reefs and forests of the tropics. QFT states that the total number of quanta N and the common phase angle φ of a swarm is an emergent consequence of that relationship (adapted from [
67,
122]).
This entry is adapted from the peer-reviewed paper 10.3390/ijms241814003