Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Cytoplasmic phosphoinositides (PI) are critical regulators of the membrane–cytosol interface that control a myriad of cellular functions despite their low abundance among phospholipids. The metabolic cycle that generates different PI species is crucial to their regulatory role, controlling membrane dynamics, vesicular trafficking, signal transduction, and other key cellular events. The synthesis of phosphatidylinositol (3,4,5)-triphosphate (PI3,4,5P3) in the cytoplamic PI3K/Akt pathway is central to the life and death of a cell.
- phosphoinositide
- scaffolding protein
- PI3K-Akt pathway
1. Introduction
Autophagy is the “self-eating” of cytosolic components and is an essential cellular homeostasis mechanism under conditions of nutrient scarcity. It is a multi-step process, forming specific autophagic organelles in each step [1]. Organelles initiate from the membrane formation of omegasomes, gradually elongate to phagophores and enclose to generate an autophagosome, and then fuse with lysosome to form autolysosomes, which degrade the cargo. Phosphoinositide metabolism plays a key role in every step of autophagy. Different phosphoinositides recruit specific compartments on membrane structures, mediating the proper generation and activation of autophagic organelles [1][2] (Figure 1).
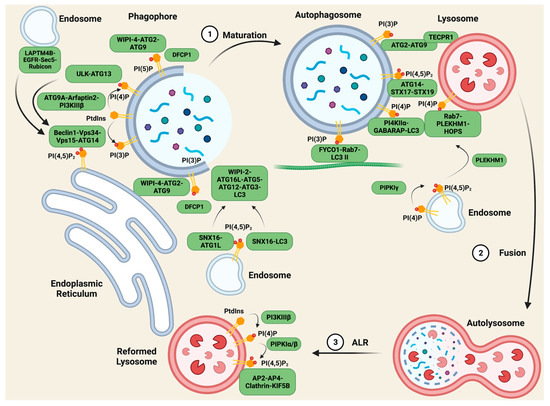
Figure 1. PIs regulate scaffolding complexes in autophagy. During the maturation of autophagosomes, the fusion between autophagosomes and lysosomes, and the reformation of lysosomes after degradation, membrane PIs play vital roles in recruiting specific scaffolding complexes to mediate lipid transfer, vesicle transport, and membrane tethering. The most important PI species in autophagy are PI(3)P and PI(4)P, as they anchor multiple crucial complexes in different steps of autophagy, while PI(5)P and PI(4,5)P2 also have unique roles. Figure created with biorender.com.
2. PI Complexes Regulate Autophagosome Maturation
The initiation of autophagic membrane structures is carried out by the protein Beclin1 [3][4][5]. Autophagy is stimulated by multiple stressors including serum starvation [6]. Upon serum starvation, inactive EGFR internalizes into LAPTM4B-positive endosomes [7]. Inactive EGFR and LAPTM4B stabilize each other at the late endosome and interact with multiple subunits of the exocyst complex, including Sec5. The EGFR-LAPTM4B-Sec5 complex recruits the autophagy inhibitor Rubicon, which in turn disassociates Rubicon from the Beclin1 complex and initiates the autophagy process [7]. The inactive EGFR-LAPTM4B complex specifically regulates autophagy, whereas the active EGF-EGFR complex prolongs EGFR signaling by blocking internalization into the lysosome where it is downregulated via the degradation of the receptor.
Beclin1 initiates the formation of the Beclin1-Vps34-Vps15 scaffolding complex, and the complex is recruited to the ER by ATG14 for autophagy specificity [8][9], while Vps34 generates PI(3)P from PtdIns. Vps34, and together with Vps15, Vps30, ATG14, and ATG38, forms PI3K complex I (PI3KCI) [8][10][11][12]. In yeast, PI3KCI constitutively binds to the vacuolar membrane protein Vac8 through ATG14 [13]. Upon autophagy initiation, ATG 1 recruits ATG9, ATG13, and ATG17-ATG31-ATG29 to assemble the ATG1 complex [14][15][16]. In an ATG1 kinase activity-dependent manner, PI3KCI associates with the ATG1 complex via the ATG38-ATG1 complex and Vps30-ATG9 interactions, anchoring PI3KC1 to the pre-autophagosomal structure for PI(3)P production [13]. A minor pool of PI(3)P is also generated from PI3KC2 in starvation-induced autophagy [17]. DFCP1, a protein containing the PI(3)P-binding FYVE domain, is recruited to these pools of PI(3)P, and together regulate the biogenesis of lipid droplets [18], acting as markers of the omegasomes [19].
PI(3)P also plays a critical role in the maturation of phagophores to autophagosomes. WIPI-2 binds to PI(3)P on the omegasomes and mediates ATG16L-ATG5-ATG12 complex formation [20]. The ATG16L complex acts together with ATG3 as an E3-like ligase and E2-enzyme for LC3 lipidation, coupling LC3 with phosphatidylethanolamine (PE) to generate LC3 II, which regulates the closure, fusion, and transport of the autophagosome [21][22]. Another PI(3)P effector, WIPI-4, forms a complex with ATG2 and ATG9 on the PI(3)P pools, tethering the autophagic membrane to the ER for lipid transfer to elongate this membrane structure [23].
Phosphoinositides are also involved in the initiation of autophagosomes. ATG9A, the ubiquitously expressed variant of ATG9, locates to autophagosomes on vesicle compartments where it binds to Arfaptin2 and PI4KIIIβ, bringing the complex to autophagic sites, and promoting the generation of PI(4)P [24]. PI(4)P then recruits the ULK-ATG13 complex, which activates Vps34 [24][25], enhancing autophagy as positive feedback. PI(5)P synthesized by PIKfyve on phagophores can act as a binding partner of DFCP1 and WIPI2 independent of the PI(3)P pool, forming scaffolding complexes and mediating the biogenesis of autophagy as mentioned above [26]. PI(4,5)P2 plays a more controversial role in autophagy initiation. On the one hand, PI(4,5)P2 phosphorylated from PI(5)P by PIPKIIs negatively regulates autophagosome biogenesis by depleting the PI(5)P pool. On the other hand, PI(4,5)P2 contributes to the autophagic membrane generated from endosomes and the ER. On ATG16L1-positive endosomes, SNX16, a PX-domain-containing protein, that specifically interacts with PI(4,5)P2, ATG16L1, and LC3, mediates the delivery of the ATG16L1 complex to the autophagic site and the lipidation of LC3 [27], while another pool of PI(4,5)P2 generated by PIPKIγi5 on the ER binds to ATG14, stabilizing the autophagy-specific Beclin-Vps34-Vps15-ATG14 scaffolding complex [28].
3. PI Complexes Regulate Autolysosome Fusion
After the maturation of the autophagosome, the next step of cellular content degradation is fusion with lysosomes, where phosphoinositide scaffolding complexes also play a vital role. To fuse with lysosomes, the autophagosome must first be transported by microtubules, and the tethering of the autophagosome and microtubule depends on FYCO1-Rab7-PI(3)P-LC3 II complex formation [29][30]. This allows the autophagosome to be transported to the cell periphery, where the lysosomes reside. Upon coming into proximity with the lysosomes, lysosome membrane protein TECPR1 interacts with autophagosome membrane proteins ATG5–ATG12, enabling its binding to autophagosome PI(3)P through its PH domain. This intervesicular complex facilitates lysosome–autophagosome fusion [31].
PI(4)P presented on both the autophagosome and lysosome membranes is crucial for their fusion. Gamma-aminobutyric acid receptor-associated protein (GABARAP) binds and recruits PI4KIIα to the autophagosome for PI(4)P generation. Knocking down this complex results in the enlargement of the autophagosome and the accumulation of IC3 II, indications of impaired autophagosome–lysosome fusion [32]. On the lysosomes, PI(4)P synthesis is controlled by PI4KIIIβ, and this PI(4)P pool interacts with Rab7, anchoring the PH domain-containing protein family member 1 (PLEKHM1) and the homotypic fusion and protein-sorting (HOPS) complex, and PLEKHM1 binds to the LC3/GABARAP complex via the LC3 interaction region [33]. In this way, the PI(4)P pool on both vesicles regulate the tethering and fusion of autophagosomes and lysosomes. Other studies indicate that PI(4,5)P2 also controls this process via multiple mechanisms. The conversion of PI(4)P into PI(4,5)P2 by PIPKIγ inactivates Rab7 and releases PLEKHM1 from endosomes, allowing the complex’s formation at autolysosome fusion sites [34], while ATG14, a PI(4,5)P2 effector, binds to the soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) domain of STX17 and SNAP29, stabilizing the STX17-SNAP29 binary t-SNARE complex on autophagosomes to facilitate the fusion [35].
4. PI Complexes Regulate Autophagic Lysosome Reformation
After degradation, autolysosomes are required to be recycled back into new lysosomes. If nutrients are scarce, this is achieved via a process called autophagic lysosome reformation (ALR). Studies show that a specific pool of PI(4,5)P2 mediates this process. PtdIns on the autolysosome is first phosphorylated by PI4KIIIβ to PI(4)P, to be used as a substrate for PIPKIα/β to generate PI(4,5)P2 [36][37]. The PI(4,5)P2 on the autolysosome surface recruits the adaptor proteins AP2 and AP4, and clathrin to induce membrane budding and tubule formation [37]. Then, the motor protein KIF5B interacts with PI(4,5)P2 in a clathrin-dependent manner, driving autolysosome tubulation and ALR [38].
This entry is adapted from the peer-reviewed paper 10.3390/biom13091297
References
- Palamiuc, L.; Ravi, A.; Emerling, B.M. Phosphoinositides in autophagy: Current roles and future insights. FEBS J. 2019, 287, 222–238. (In English)
- Tan, X.; Thapa, N.; Choi, S.; Anderson, R.A. Emerging roles of PtdIns(4,5)P2--beyond the plasma membrane. J. Cell Sci. 2015, 128, 4047–4056. (In English)
- Zalckvar, E.; Berissi, H.; Mizrachy, L.; Idelchuk, Y.; Koren, I.; Eisenstein, M.; Sabanay, H.; Pinkas-Kramarski, R.; Kimchi, A. DAP-kinase-mediated phosphorylation on the BH3 domain of beclin 1 promotes dissociation of beclin 1 from Bcl-XL and induction of autophagy. EMBO Rep. 2009, 10, 285–292. (In English)
- Wei, Y.; Pattingre, S.; Sinha, S.; Bassik, M.; Levine, B. JNK1-mediated phosphorylation of Bcl-2 regulates starvation-induced autophagy. Mol. Cell 2008, 30, 678–688. (In English)
- Shi, C.S.; Kehrl, J.H. TRAF6 and A20 regulate lysine 63-linked ubiquitination of Beclin-1 to control TLR4-induced autophagy. Sci. Signal. 2010, 3, ra42. (In English)
- Debnath, J.; Gammoh, N.; Ryan, K.M. Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell Biol. 2023, 24, 560–575.
- Tan, X.; Thapa, N.; Sun, Y.; Anderson, R.A. A kinase-independent role for EGF receptor in autophagy initiation. Cell 2015, 160, 145–160.
- Itakura, E.; Kishi, C.; Inoue, K.; Mizushima, N. Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG. Mol. Biol. Cell 2008, 19, 5360–5372. (In English)
- Baskaran, S.; Carlson, L.-A.; Stjepanovic, G.; Young, L.N.; Kim, D.J.; Grob, P.; Stanley, R.E.; Nogales, E.; Hurley, J.H. Architecture and dynamics of the autophagic phosphatidylinositol 3-kinase complex. eLife 2014, 3, e05115. (In English)
- Kihara, A.; Noda, T.; Ishihara, N.; Ohsumi, Y. Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae. J. Cell Biol. 2001, 152, 519–530.
- Zhong, Y.; Wang, Q.J.; Li, X.; Yan, Y.; Backer, J.M.; Chait, B.T.; Heintz, N.; Yue, Z. Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1-phosphatidylinositol-3-kinase complex. Nat. Cell Biol. 2009, 11, 468–476.
- Araki, Y.; Ku, W.-C.; Akioka, M.; May, A.I.; Hayashi, Y.; Arisaka, F.; Ishihama, Y.; Ohsumi, Y. Atg38 is required for autophagy-specific phosphatidylinositol 3-kinase complex integrity. J. Cell Biol. 2013, 203, 299–313.
- Hitomi, K.; Kotani, T.; Noda, N.N.; Kimura, Y.; Nakatogawa, H. The Atg1 complex, Atg9, and Vac8 recruit PI3K complex I to the pre-autophagosomal structure. J. Cell Biol. 2023, 222, 10017.
- Kamada, Y.; Funakoshi, T.; Shintani, T.; Nagano, K.; Ohsumi, M.; Ohsumi, Y. Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J. Cell Biol. 2000, 150, 1507–1513.
- Fujioka, Y.; Suzuki, S.W.; Yamamoto, H.; Kondo-Kakuta, C.; Kimura, Y.; Hirano, H.; Akada, R.; Inagaki, F.; Ohsumi, Y.; Noda, N.N. Structural basis of starvation-induced assembly of the autophagy initiation complex. Nat. Struct. Mol. Biol. 2014, 21, 513–521.
- Fujioka, Y.; Alam, J.M.; Noshiro, D.; Mouri, K.; Ando, T.; Okada, Y.; May, A.I.; Knorr, R.L.; Suzuki, K.; Ohsumi, Y.; et al. Phase separation organizes the site of autophagosome formation. Nature 2020, 578, 301–305.
- Devereaux, K.; Dall’armi, C.; Alcazar-Roman, A.; Ogasawara, Y.; Zhou, X.; Wang, F.; Yamamoto, A.; De Camilli, P.; Di Paolo, G. Regulation of mammalian autophagy by class II and III PI 3-kinases through PI3P synthesis. PLoS ONE 2013, 8, e76405. (In English)
- Cheung, P.C.; Trinkle-Mulcahy, L.; Cohen, P.; Lucocq, J.M. Characterization of a novel phosphatidylinositol 3-phosphate-binding protein containing two FYVE fingers in tandem that is targeted to the Golgi. Biochem. J. 2001, 355 Pt 1, 113–121. (In English)
- Axe, E.L.; Walker, S.A.; Manifava, M.; Chandra, P.; Roderick, H.L.; Habermann, A.; Griffiths, G.; Ktistakis, N.T. Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol. 2008, 182, 685–701. (In English)
- Dooley, H.C.; Razi, M.; Polson, H.E.; Girardin, S.E.; Wilson, M.I.; Tooze, S.A. WIPI2 links LC3 conjugation with PI3P, autophagosome formation, and pathogen clearance by recruiting Atg12-5-16L1. Mol. Cell 2014, 55, 238–252. (In English)
- Fujita, N.; Itoh, T.; Omori, H.; Fukuda, M.; Noda, T.; Yoshimori, T. The Atg16L complex specifies the site of LC3 lipidation for membrane biogenesis in autophagy. Mol. Biol. Cell 2008, 19, 2092–2100. (In English)
- Hanada, T.; Noda, N.N.; Satomi, Y.; Ichimura, Y.; Fujioka, Y.; Takao, T.; Inagaki, F.; Ohsumi, Y. The Atg12-Atg5 conjugate has a novel E3-like activity for protein lipidation in autophagy. J. Biol. Chem. 2007, 282, 37298–37302. (In English)
- Chowdhury, S.; Otomo, C.; Leitner, A.; Ohashi, K.; Aebersold, R.; Lander, G.C.; Otomo, T. Insights into autophagosome biogenesis from structural and biochemical analyses of the ATG2A-WIPI4 complex. Proc. Natl. Acad. Sci. USA 2018, 115, E9792–E9801. (In English)
- Judith, D.; Jefferies, H.B.J.; Boeing, S.; Frith, D.; Snijders, A.P.; Tooze, S.A. ATG9A shapes the forming autophagosome through Arfaptin 2 and phosphatidylinositol 4-kinase IIIβ. J. Cell Biol. 2019, 218, 1634–1652. (In English)
- Karanasios, E.; Walker, S.A.; Okkenhaug, H.; Manifava, M.; Hummel, E.; Zimmermann, H.; Ahmed, Q.; Domart, M.-C.; Collinson, L.; Ktistakis, N.T. Autophagy initiation by ULK complex assembly on ER tubulovesicular regions marked by ATG9 vesicles. Nat. Commun. 2016, 7, 12420. (In English)
- Vicinanza, M.; Korolchuk, V.I.; Ashkenazi, A.; Puri, C.; Menzies, F.M.; Clarke, J.H.; Rubinsztein, D.C. PI(5)P regulates autophagosome biogenesis. Mol. Cell 2015, 57, 219–234. (In English)
- Knævelsrud, H.; Søreng, K.; Raiborg, C.; Håberg, K.; Rasmuson, F.; Brech, A.; Liestøl, K.; Rusten, T.E.; Stenmark, H.; Neufeld, T.P.; et al. Membrane remodeling by the PX-BAR protein SNX18 promotes autophagosome formation. J. Cell Biol. 2013, 202, 331–349. (In English)
- Tan, X.; Thapa, N.; Liao, Y.; Choi, S.; Anderson, R.A. PtdIns(4,5)P2 signaling regulates ATG14 and autophagy. Proc. Natl. Acad. Sci. USA 2016, 113, 10896–10901. (In English)
- Olsvik, H.L.; Lamark, T.; Takagi, K.; Larsen, K.B.; Evjen, G.; Øvervatn, A.; Mizushima, T.; Johansen, T. FYCO1 Contains a C-terminally Extended, LC3A/B-preferring LC3-interacting Region (LIR) Motif Required for Efficient Maturation of Autophagosomes during Basal Autophagy. J. Biol. Chem. 2015, 290, 29361–29374. (In English)
- Pankiv, S.; Alemu, E.A.; Brech, A.; Bruun, J.-A.; Lamark, T.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end-directed vesicle transport. J. Cell Biol. 2010, 188, 253–269. (In English)
- Chen, D.; Fan, W.; Lu, Y.; Ding, X.; Chen, S.; Zhong, Q. A mammalian autophagosome maturation mechanism mediated by TECPR1 and the Atg12-Atg5 conjugate. Mol. Cell 2012, 45, 629–641. (In English)
- Wang, H.; Sun, H.-Q.; Zhu, X.; Zhang, L.; Albanesi, J.; Levine, B.; Yin, H. GABARAPs regulate PI4P-dependent autophagosome:lysosome fusion. Proc. Natl. Acad. Sci. USA 2015, 112, 7015–7020. (In English)
- McEwan, D.G.; Popovic, D.; Gubas, A.; Terawaki, S.; Suzuki, H.; Stadel, D.; Coxon, F.P.; de Stegmann, D.M.; Bhogaraju, S.; Maddi, K.; et al. PLEKHM1 regulates autophagosome-lysosome fusion through HOPS complex and LC3/GABARAP proteins. Mol. Cell 2015, 57, 39–54. (In English)
- Baba, T.; Toth, D.J.; Sengupta, N.; Kim, Y.J.; Balla, T. Phosphatidylinositol 4,5-bisphosphate controls Rab7 and PLEKHM1 membrane cycling during autophagosome-lysosome fusion. EMBO J. 2019, 38, e100312. (In English)
- Diao, J.; Liu, R.; Rong, Y.; Zhao, M.; Zhang, J.; Lai, Y.; Zhou, Q.; Wilz, L.M.; Li, J.; Vivona, S.; et al. ATG14 promotes membrane tethering and fusion of autophagosomes to endolysosomes. Nature 2015, 520, 563–566. (In English)
- Sridhar, S.; Patel, B.; Aphkhazava, D.; Macian, F.; Santambrogio, L.; Shields, D.; Cuervo, A.M. The lipid kinase PI4KIIIβ preserves lysosomal identity. EMBO J. 2012, 32, 324–339. (In English)
- Rong, Y.; Liu, M.; Ma, L.; Du, W.; Zhang, H.; Tian, Y.; Cao, Z.; Li, Y.; Ren, H.; Zhang, C.; et al. Clathrin and phosphatidylinositol-4,5-bisphosphate regulate autophagic lysosome reformation. Nat. Cell Biol. 2012, 14, 924–934. (In English)
- Du, W.; Su, Q.P.; Chen, Y.; Zhu, Y.; Jiang, D.; Rong, Y.; Zhang, S.; Zhang, Y.; Ren, H.; Zhang, C.; et al. Kinesin 1 Drives Autolysosome Tubulation. Dev. Cell 2016, 37, 326–336. (In English)
This entry is offline, you can click here to edit this entry!