1. Introduction
The hydrolysis of proteins by proteases produces short-sequence amino acids with special physiological functions called bioactive peptides [
1]. Bioactive peptides have mostly been isolated from a variety of animals and plants. Bioactive peptides are often obtained from animal sources such as milk and dairy products [
2], eggs [
3] meat [
4] and fish-derived products [
5]. Bioactive peptides from plants are mainly obtained from legumes [
6], grains [
7], nuts [
8], fruits and vegetables [
9]. Until now, a significant amount of research about bioactive peptides isolated from animals and plants has been intensively reported, while relatively few studies have been conducted on edible mushrooms bioactive peptides (MBPs). Currently, there are at least 12,000 species of mushrooms, of which 2000 are reported to be edible, and about 200 are collected as food or pharmaceutical ingredients [
10]. Therefore, edible mushrooms are rich and vast in relatively untapped resources. Different bioactive compounds isolated from different species of mushrooms can be used to develop different functional foods. Edible mushrooms are excellent functional foods and nutritional supplements due to their high quantities of bioactive metabolites, including proteins, polysaccharides, enzymes (e.g., superoxide dismutase), dietary fibre and many other biomolecules [
11]. For example,
Trametes versicolor are commonly marketed as dietary supplements in the form of tablets, capsules and powders [
12]. Recently, the natural flavour compounds in
Trametes versicolor have been valuable ingredients in the production of non-alcoholic or low-alcoholic beers[
13,
14].Based on US Department of Agriculture(
https://fdc.nal.usda.gov/ (accessed on 8 July 2020)) statistics, it can be seen that edible mushrooms contain higher protein than most vegetables, and many proteins have bioactivities, while more or even new MBPs remain to be discovered [
16,
1]. MBPs can be absorbed by the intestine completely, producing direct partial benefits in the digestive tract without increasing the functional burden on the gastrointestinal tract [1]. MBPs can exert beneficial physiological effects by entering the circulatory system in their intact state. MBPs can provide high organisational affinity [
17], low toxicity [
18], and high stability [
19], with a range of nutritional, functional and biological properties. Endogenous MBPs may exert antioxidant and anti-inflammatory activities by regulating antioxidant pathways and activating immune cells. MBPs may also act as analgesics or opioids in improving memory and cognitive deterioration in nerve cells [
20]. Exogenous MBPs may be used as functional foods and medicines to ameliorate inflammation, hyperglycemia, hypertension, high cholesterol and other degenerative diseases[
22]. For consumers who prefer not to consume animal-derived products, MBPs can be sources of ideal alternative supplements for replacing animal-derived food [
23]. Replacing some of the meat or fish with healthy natural foods is a trend for the future market. Edible mushrooms have a similar taste to meat products and are rich in many nutrients. For this reason, many entrepreneurs have incorporated edible mushrooms into muscle foods (e.g., meat and fish) to reduce the proportion of meat [
<>].
2. Preparation of MBPs
Edible mushrooms are important sources for naturally bioactive proteins and peptides, which provide an excellent material basis for the discovery of MBPs. The protein content of edible mushrooms by-products is about 25%, and bioactive peptides extracted from edible mushrooms by-products can be recycled for the production of functional foods, which can effectively reduce the waste of resources [
24]. The extraction methods are diverse because of the different growing environments and different species of edible mushrooms. As a result, the structures and bioactivities of MBPs also vary [
9]. Basically, there are two popular approaches to prepare MBPs; one is to extract endogenous MBPs directly from the mushrooms. Many endogenous MBPs have been extracted directly from fresh fruiting bodies, dried powders or fermented powders, such as peptides with antimicrobial and ACE inhibitory properties, respectively [
25]. The other approach uses proteolytic reactions of exogenous enzymes to release peptide fragments from mushroom proteins isolated from edible mushrooms or their mycelia indirectly (e.g., hydrolysis by bromelain) to liberate potent peptides from intact proteins [
9]. Indeed, MBPs with a variety of biological activities, such as antioxidative, antibacterial, anti-inflammatory, anti-aging, antitumour and anti-diabetic activities, were obtained using enzymatic hydrolysis, for example, peptides with ACE inhibition hydrolysis from
Lentinus edodes with Alcalase [
26]. MBPs contain ergothioneine, laccase, ribonuclease and other enzymes [
27]. Low molecular weight MBPs obtained with the assistance of ultra-high-pressure processing were found to activate ethanol dehydrogenase and aldehyde dehydrogenase in vitro. They could effectively catalyse the gradual conversion of alcohol into acetaldehyde and acetic acid with lower toxicity, and prevent alcoholic liver injury [
15].
Chemical hydrolysis is also one of the commonly used methods to prepare MBPs. Chemical hydrolysis involves breaking the peptide chains of a protein using an acid or base solution, producing peptides and free amino acids. Chemical hydrolysis in the food industry has a variety of limitations, such as difficulties controlling the process, loss of nutrients and pollution of the environment [
28]. Microbial fermentation is also one of the commonly used methods to obtain MBPs in recent years. Microorganisms have protein hydrolysis systems that can produce proteases or induce fermentation to obtain MBPs. The type of MBPs depends on the fermentation time, the strain and the type of edible mushrooms protein [
29].
Microbial fermentation can avoid impurities in peptides produced by enzymatic digestion, shorten preparation time and reduce production costs.Ultrafiltration, hydrophilic interaction chromatography (HILIC), fast protein liquid chromatography (FPLC), ion exchange chromatography (IEC), size exclusion chromatography (SEC), gel filtration chromatography (GFC) or high-performance liquid chromatography (HPLC) [
25] are methods commonly used for purification after extraction and preliminary bioactivity screening. For example, antihypertensive peptides were separated and characterised from the
Agaricus bisporus using SEC, reversed-phase-high-performance liquid chromatography (RP-HPLC) and liquid chromatography–mass spectrometry (LC–MS/MS) [
25].
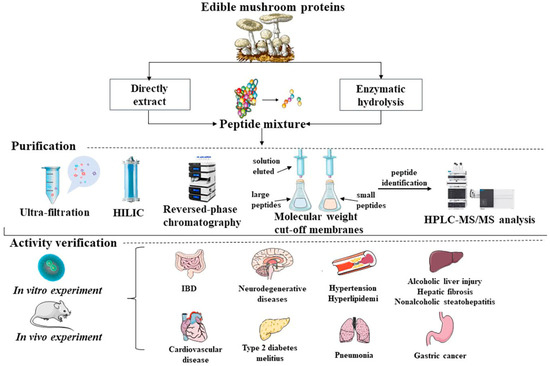
MBPs alleviate symptoms associated with targeted damage or degenerative diseases. In addition, MBPs are known to exert antioxidant effects by inactivating reactive oxygen species and scavenging free radicals to improve degenerative diseases such as inflammation, hypertension, hyperglycemia, cardiovascular disease, high cholesterol, memory and cognitive diseases with antioxidant, antibacterial, anti-aging and antitumour effects (Figure 1).
Figure 1. Common extraction routes, purification and preliminary bioactivities screening, in vivo and in vitro validation of biological activities of MBPs.
3. Bioactivities of MBPs
3.1. Antioxidant Activity
Edible mushrooms are rich in a variety of peptides with antioxidant activity [
30]. Due to the different raw ingredients and preparation processes, MBPs may have different targeting functions. The antioxidant mechanisms of MBPs are mainly categorised into those involving regulation of ROS production, regulation of antioxidant enzyme activities and regulation of antioxidant pathways. Free radicals may be scavenged by MBPs through providing protons, electrons and chelating metal ions to regulate the production of ROS
[
31]. Low molecular weight peptides isolated from
Agaricus bisporus (ABP) and Pleurotus eryngii mycelium (PEMP) were abundant in negatively charged amino acids, which could neutralise free radicals and regulate the production of ROS [
32]. G. lucidum peptide (GLP) [
33]
exerted antioxidant effects in the soybean oil system by blocking soybean lipoxygenase activity in a dose-dependent manner, with an IC50 value of 27.1 µg/mL. Compared to butylated hydroxytoluene, GLP had the better antioxidant activity through scavenging hydroxy radicals and quenching superoxide radical anions in biological systems.Organisms can regulate the production of ROS in time, through endogenous enzymes and non-enzymatic defensive systems
[
34]. The oral administration of GLP exerted significant hepatoprotective effects through antioxidant activity in mice with liver injuries [
35]. GLP increased the levels of glutathione (GSH) and superoxide dismutase (SOD), decreasing the levels of malondialdehyde (MDA) in the liver. Furthermore, it decreased the activities of alanine transaminase (ALT) and aspartate transaminase (AST) in the serum to resist hepatic fibrosis and alcoholic liver injury. Research showed that GLP could effectively inhibit the hazards of peroxide produced in mitochondria by regulating the activity of antioxidant enzymes, showing excellent antioxidant activity.
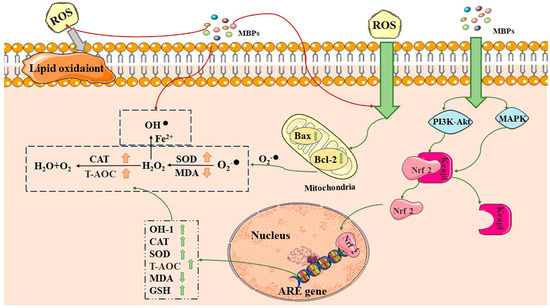
MBPs achieve antioxidant activity by modulating antioxidant pathways such as the Kelch-like ECH-associated protein 1-nuclear factor erythroid-2-related factor 2-antioxidant responsive elements (Keap1-Nrf2-ARE), nuclear factor-κ light chain enhancer of activated B cells (NF-κB), mitogen-activated protein kinase (MAPK) and the phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) pathways [
36,
37,
38]. MBPs may regulate the expression of related antioxidant proteins by down-regulating the Keapl gene and up-regulating Nrf2 gene expression
(Figure 2) [
31]. The GLP [
39] promoted Nrf2 and activated the Nrf2-ARE signalling pathway, exhibiting antioxidant effects on cells induced by hydrogen peroxide (H
2O
2). Nrf2 dissociated from Keap1 upon stimulation of the PI3K/AKT and MAPK pathways, moving to the nucleus, and associated with the antioxidant component ARE, which regulated the expression of antioxidant enzymes such as HO-1, catalase (CAT), etc.
[
40].
Figure 2. Antioxidant and anti-aging mechanisms of MBPs. MBPs may exert antioxidant and anti-aging effects by regulating Keap1-Nrf2-ARE through the PI3K/AKT and MAPK pathways. CAT: catalase, SOD: superoxide dismutase, T-AOC: total antioxidant capacity, MDA: malondialdehyde, GSH: glutathione, Bax: BCL2-Associated X, Bcl-2: B-cell lymphoma-2. Excess ROS by generating superoxide anions via Bax and Bcl-2 in cellular mitochondria. MBPs regulate ROS content by providing protons, electrons and chelating metal ions. MBPs regulate the Keap1-Nrf2-ARE signalling pathway through the PI3K/AKT and MAPK pathways. Red arrows indicate that MBPs regulate ROS content by providing protons, electrons, and chelating metal ions. The grey arrows indicate that excess ROS are harmful by generating superoxide anions. Green arrows indicate that MBPs and ROS react via different pathways within the cell.
Anti-Aging Activity
Aging occurs in cells, organs and the whole organism, which leads to a reduction in biological function of the organism’s ability to eliminate oxidative stress. The accumulation of excess free radicals causes MDA levels to increase, the total antioxidant capacity (T-AOC) to reduce, and disrupt cellular structure [
41], leading to cellular senescence and death. CMP [
42] and GLP [
43] had dose-dependent scavenging effects on oxygen free radicals and hydroxyl free radicals. SOD is a critical mitochondrial enzyme antioxidant associated with longevity [
44]. MBPs could significantly enhance senescence-associated mitochondrial enzyme antioxidants. CMP and GLP scavenged hydroxyl radicals better than the specific hydroxyl radical scavenger mannitol [
45]. Agaricus blazei peptide (ABp) reduced MDA and ROS contents, and increased CAT and T-AOC activities in a D-galactose-induced senescence model of the NIH/3T3 cell [
46].
The main targets of MBPs to slow down the aging process include the metabolic mitochondrial pathway, inactivating reactive oxygen species, scavenging free radicals, alleviating the oxidation of biomarkers in organisms, and reconstructing homeostatic mechanisms in vivo [
47]. Nrf2 activity was tightly correlated with degenerative diseases induced by aging, and contributed to the prevention and mitigation of degenerative diseases. In the aging model established by D-galactose in mice, feeding ABp was found to lead to the down-regulation of Keap1 protein expression, thus up-regulating Nrf2. In the Keap1–Nrf2 pathway, the expressions of HO-1 and related factors such as ApoE, Hsph1 and Trim32 were up-regulated, effectively scavenging free radicals and showing excellent anti-aging activity [
48]. Progressive changes were found in the epigenetic information in both dividing and non-dividing cells in modifications involving chromatin transformations, histone and DNA methylation patterns in research related to aging. The major features of aging are constituted in DNA and histones methylation, accompanied by other epigenetic alterations [
49]. In previous studies, it was found that the epigenetic alterations of aging could be modulated by the consumption of peptides that directly maintain telomere length [
44]. Current research about whether MBPs could regulate telomere length in different organisms has not been conducted. This would be a new area of research on the anti-aging activities of MBPs, and more relevant research is needed in the future.
In conclusion, MBPs can act on multiple targets simultaneously to exert their functional antioxidant and age-delaying activities. MBPs have a wide scope for development as one of the components of natural antioxidant and anti-aging functional foods. Currently, MBPs have become a new direction of research on functional foods. However, further research is needed on how MBPs replace synthetic antioxidants.
3.2. Antimicrobial Activity
Antibiotics can treat infections such as tuberculosis, pneumonia, leprosy and gonorrhoea. However, antibiotic resistance [
54,
55] has emerged from the overuse and misuse of antibiotics. Natural bioactive peptides are characterised by their high efficacy, stability and low toxicity, making them major alternatives to antibiotics and conventional drugs [
56]. MBPs have been widely studied for their antimicrobial activities. A variety of antimicrobial peptides have been isolated and purified from edible mushrooms such as
Polyporus alveolaris,
Pleurotus eryngii [
57],
Lentinus edodes and
Agrocybe cylindracea [
58]. The possible antimicrobial mechanisms of MBPs could involve either regulating and leading to tissue-specific expression patterns, or making intracellular protein leakage and leading to bacterial death.
The hydrophobic amino acids of MBPs could regulate the NF-κB pathway and the MAPK pathway. PEMP [
32] had good antibacterial activity by stimulating macrophage proliferation, increasing phagocytosis activities, TLRs expression, and releases of tumour necrosis factor-α (TNF-α), IL-6, NO and H
2O
2. MBPs are rich in structures with α-helices, β-folds, random coils and disulfide bonds [
59]. Antimicrobial peptides are enriched with hydrophobic amino acids, α-helixes, β-folds, random coils and disulfide bonds.
Another antibacterial mechanism of MBPs is the disruption of bacterial cell membranes, causing intracellular protein leakage within the bacterial cells to achieve antibacterial effects. Antimicrobial peptides from the mycelia of
Cordyceps militaris [
60] may cause intracellular protein leakage of E. coli (ATCC 25922), which maintained the integrity of the intestinal mucosa and attenuated E. coli infections in mice. Antibacterial peptides isolated from the mycelia (GLM) and fruiting bodies (GLF) of G. lucidum [
61] showed a dose-dependent increase at 50–125 μg/mL in protein leakages from
Escherichia coli and
Staphylococcus aureus. It is clear that GLF and GLM induces cell death with strong antibacterial activities against both
Escherichia coli and
Staphylococcus aureus.
Based on the above, MBPs show good antibacterial activities against bacteria that are resistant to drugs. Thus, MBPs could be important natural alternatives to antibiotics.
3.3. Anti-Inflammatory Activity
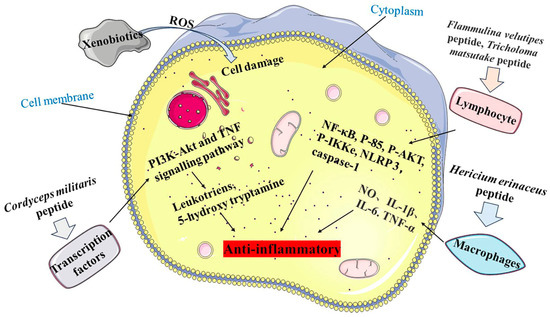
MBPs may enhance the cytotoxicities of natural killer cells and the phagocytosis of macrophages, promoting multiplication and maturation of immune cells and lymphocytes and inhibiting pro-inflammatory responses, thereby improving the host’s defence against invading pathogens (
Figure 3) [
69]. Yu [
70] prepared a bioactive peptide, KSPLY, with a molecular weight of 608.3834 Da, from
Hericium erinaceus. KSPLY promoted TNF-α, NO, IL-6 and IL-1β secreted by macrophages, which inhibited lipopolysaccharide (LPS)-induced inflammatory responses at a concentration of 100 μmol/L. The proliferation of splenic lymphocytes was significantly reduced in mice fed with
Pleurotus eryngii peptide (PEP), and the serum haemolysin level of CTX-induced mice was significantly increased [
71]. This confirmed that PEP could significantly improve humoral immune function of immunosuppressed mice.
Figure 3. Anti-inflammatory mechanism of MBPs. Pathway 1 is Cordyceps militaris peptide regulating the PI3K-Akt and TNF signalling pathways to release Leukotriens and 5-hydroxy tryptamine. Pathway 2 is Flammulina velutipes peptide and Tricholoma matsutake peptide acting on lymphocytes, which in turn regulate NF-κB, P-85, P-AKT, P-IKKe, NLRP 3 and caspase-1. Pathway 3 is Hericium erinaceus peptide acting on macrophages to release NO, IL-1β, IL-6 and TNF-α.
MBPs may enhance the oxidative defensive system and barrier function by promoting the production of antibodies, cytokines and chemokines to reduce the inflammatory response. After local nasal immunotherapy (LNIT) with
Flammulina velutipes peptides (FIP-fve), the production of proinflammatory cytokines and chemokines were significantly reduced [
72]. It has been shown that FIP-fve induces the potent activator of peripheral lymphocytes through activation of the p38 mitogen-activated protein kinase (p38 MAPK) signalling pathway with anti-inflammatory activity. The
Tricholoma matsutake-derived peptide WFNNAGP [
53] attenuated the inflammatory response by inhibiting the expression of pro-inflammatory cytokines and myeloperoxidase (MPO), as it promoted expressions of tight junction proteins closing ribbon-1, claudin and occluding. WFNNAGP reduced colonic inflammation in mice by down-regulating NF-κB expression to inhibit formation and activation of NLRP 3 and caspase-1.
MBPs have a variety of target cells and sites, such as NK cells, CD4, CD25, T lymphocytes, macrophages [
73], monocytes, B lymphocytes [
74] and mast cells [
75], among others. The results of a study showed that FIP-fve could make changes in Treg-associated immunity, with down-regulation of IL-4/CD4 T-cell expression and up-regulation of IFN-γ/CD4 T-cell expression in mice. FIP-fve effectively decreased inflammatory cell infiltration and epithelial damage [
72]. Experiments demonstrated that the anti-inflammatory effect of orally administering FIP-fve on mite-induced airway inflammations in mice. Bioactive polypeptides of
Cordyceps militaris (CMP) [
76] were implicated in the regulation of immune function in mice through transcription factors Ets1 and the Spp1, Rel, and Smad3 genes. CMP regulated TNF and the PI3K-Akt signalling pathway, playing an important role in inflammation by increasing immune organ indexes, the number of leukocytes and the content of hemolysin in the sera of mice. In summary, MBPs have good anti-inflammatory activity, and offer new ideas for the development of functional food supplements as natural ingredients.
++++++3.4. Memory and Cognitive Improvement Activity
Medications that are commonly used clinically to prevent and improve learning in learning and memory impairments include free radical scavengers, acetylcholinesterase (AChE) inhibitors and drugs to prevent the formation of amyloid β deposits [
77]. However, these types of drugs are associated with adverse side effects, high toxicity, poor memory improvement and no preventive effect on the onset and development of neurological disorders. CMP scavenges oxygen free radicals, decreasing AChE activity in the mouse brain, which may reduce the damage of cholinergic neurological nerves, hence improving learning and memory ability in a scopolamine-induced mice model of learning and memory impairment [
78]. The mechanism of CMP is similar to that of free radical scavengers and AChE inhibitors. CMP promotes the expressions of Il-1β and Slc18a2, as well as secretion of neurotransmitters in mice that reduce dyskinesia and depression-like behavior, and contributed to improve learning and memory. Hericium erinaceus [
79] fed to mice prevented the loss of spatial short-term and impairments of visual recognition memory induced by the formation of amyloid β deposits. However, there are relatively few studies on improvements in memory and cognitive ability by
Hericium erinaceus peptides, and more experiments are needed to verify its mechanism in the future.
Based on the above, MBPs may effectively prevent and improve learning and memory disorders, with have the advantages of fewer side effects, less toxicity, more targets of action and lower costs. MBPs may be more effective in preventing memory and cognitive disorders compared with synthetic drugs.
3.5. Anti-Hypertensive Activity
Hypertension is currently one of the most common metabolic diseases, and is a predisposing factor for many other conditions such as renal failure and heart disease. The inhibition of angiotensin-converting enzyme (ACE) regulation of the renin–angiotensin system is thought to be the main mechanism of hypotensive activity [
80]. Edible mushrooms are good sources of hypotensive peptides, and many antihypertensive peptides have been isolated. Kaprasob [
81] found that LIYAQGFSK peptide extracted from the King Boletus mushroom had the greatest ACE-binding energy of −9.2 kcal/mol through hydrogen bonds, and exhibited ACE inhibitory activity. The AHEPVK, RIGLF and PSSNK peptides of ABP showed stronger ACE inhibitory activities with lower IC
50 values after gastrointestinal digestion compared to ACE inhibitory peptides (ACEIPs) from other sources [
25]. MBPs inhibit ACE mainly through the inhibition of nucleic acid synthesis or binding, blocking protein synthesis, membrane permeabilization, inhibition of enzyme activities and triggering apoptosis. Gln-Leu-Val-Pro (QLVP) is a polypeptide with its amino acid sequence isolated from the mycelia of
Ganoderma lucidum. QLVP significantly activated the angiotensin I-mediated phosphorylation of endothelial nitric oxide synthase in human umbilical vein endothelial cells, and partially reduced mRNA and protein expressions of vasoconstrictor factor endothelin-1, which showed value in the application of GLP for treating hypertension in metabolic diseases. Overall, MBPs exhibit good anti-hypertensive activities as potential ingredients for natural anti-hypertensive functional foods and nutraceuticals.
3.6. Antitumour Activity
Biological therapy is an extensively alternative approach to limit the growth of cancer cells that uses biologically active compounds to treat cancer. Bioactive peptides in food can inhibit cancer at all stages of the disease, and offer advantages such as their greater affinity, target-specific effects, reducing toxicity and superior tissue penetration, in comparison to the side effects of chemotherapeutic molecules [
87]. PEP was found to inhibit the growth of cancer cells in concentrations of 0.05 to 2 mg/mL. It inhibited the growth of gastric cancer (HGC-27), breast cancer (BT-549) and cervical cancer (Hela-229) cells by 61.40%, 59.20%, and 32.80%, respectively, and demonstrated strong antitumour activity. The MBPs exerted anti-tumour activity by stimulating the host′s defensive mechanism to enhance its non-specific immune responses. CMP [
42] activated the reticuloendothelial system and macrophages to promote lymphocyte transformation and immunologically active cells like lymphocytes, lymphokines, monocyte macrophage systems and NK cells to attack target cells to have anti-tumour effects. The amino acid sequence of the Ser-Leu-Ser-Leu-Ser-Val-Ala-Arg peptide extracted from morels (
Morchella spp.) were found to reduced cell proliferation via a mitochondrial-dependent pathway. It was shown that the movement of cytochrome C from the mitochondria to the cytoplasm could be facilitated by the down-regulation of Bcl-2/Bax, thereby promoting the expression of caspase-9 and caspase-3 and suppressing tumourigenesis [
88]. MBPs facilitated intermediary metabolism or controlled DNA transcription and translation through the activation of related enzyme systems. Mice fed with CMP [
89] were found to significantly inhibit polymorphonuclear cells infiltration and the infrared ray-induced up-regulation of brain production of C3 protein levels, and the production of interleukin-1β and tumour necrosis factor-α. MBPs exhibit antitumour effects, and may be promising natural tumour preventive agents.
3.7. Other Activities
In addition to their antioxidant, antibacterial, anti-inflammatory, anti-aging, memory and cognitive improving, anti-hypertension and anti-tumour activities shown above, MBPs also have functional activities, such as reducing the levels of plasma glucose with induced diabetes, and decreasing cholesterol levels.
Diabetes is a metabolic disease that is characterised by hyperglycaemia. Prolonged uncontrolled hyperglycaemia in an organism can generate oxidative stress, promote apoptosis, activate protein kinase isozymes and transcription factors, and other pathophysiological mechanisms that impair the proper functioning of tissues and organs [
90]. Currently, glycaemia is mainly controlled by the intramuscular injection of hormones or taking synthetic anti-diabetic agents such as insulin, thiazolidinediones, dipeptidyl peptidase 4 inhibitors, biguanides, sulfonylureas and sodium–glucose cotransporter type 2 inhibitors [
91]. These hypoglycaemic agents are highly effective, but with side effects [
92]. Compared with other synthetic hypoglycaemic substances, MBPs show fewer side effects, and may regulate glycaemia by inhibiting the activities of α-amylase, α-glucosidase and dipeptidyl peptidase-IV. At 2.0 mg/mL, a morel peptide inhibited α-amylase and α-glucosidase by 34.93% and 30.56%, respectively [
93], showing excellent anti-diabetic activity.
Hypercholesterolaemia arises from increased endogenous cholesterol synthesis or an excess supply of dietary cholesterol or cholesterol precursors [
94]. Bioactive peptides may inhibit the synthesis and secretion of triglycerides and cholesterol by stimulating bile acid secretion from the gallbladder, modulating hormones and cholesterol receptors, and altering hepatic lipid metabolism [
94]. MBPs are rich in inhibitors of HMG-CoA reductase, a pivotal enzyme in endogenous cholesterol biosynthesis, as well as ergocalciferol, eritadenine and β-glucan derivatives. MBPs also have a similar mechanism of action to simvastatin or ezetimibe, which are statins [
95]. The hydrophobic amino acids of Phe, Leu, Val, Gly and Pro in bioactive peptides may promote cholesterol homeostasis in organisms by stimulating the elimination of exogenous cholesterol during gastric emptying [
96]. ABP prepared by Feng [
97] reduced the solubility of cholesterol by forming micellar structures in a manner similar to the cholesterol-lowering mechanism of statins. All functional foods with MBPs may lower serum cholesterol levels. Unlike other food derivatives, they may regulate cholesterol homeostasis in organisms through different transcriptional and translational mechanisms. These mechanisms are not completely elucidated, and may be different from those of plants that have already been identified [
95].
This entry is adapted from the peer-reviewed paper 10.3390/foods12152935