Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Immunology
Ionizing radiation therapy is an important component of cancer treatment. This review provides a summary of the latest advancements, clinical use, and limitations of radiation therapy. Moreover, this review explores how radiation affects the body’s natural defense system, which plays a crucial role in fighting cancer. The immune responses triggered by radiation therapy help the body eliminate tumors naturally.
- radiation therapy
- DNA damage
- dendric cells
- NK cells
- macrophages
1. Introduction
Cancer remains the leading cause of death globally. According to the American Cancer Society (ACS), in 2023, an estimated 1,958,310 new cancer cases and 609,820 deaths from all cancers are anticipated to occur in the United States [1]. Cancer incidences increased for prostate cancer (PCa) by 3% annually from the year 2014 through 2019, following two decades of decline, translating to an additional 99,000 new cancer cases [1]. RT is one of the most effective forms of oncology care and continues to play a major role in the treatment of various types of malignancies. The emergence of advanced technologies has propelled a revolution in the field of RT, empowering clinicians with tools and techniques to deliver accurate and safe delivery of radiation doses to tumor cells. The advancement in intensity-modulated radiation therapy (IMRT), stereotactic body radiation therapy (SBRT) and proton therapy have revolutionized the field. These technological advancements have resulted in increasingly conformal radiation treatments. In this review, we present the technological advancement in various types of RT methods and highlight their clinical utility and associated limitations.
Ionizing radiation (IR) promotes primary effects on DNA structure by directly inducing DNA strand breaks, particularly double-strand breaks (DSBs), and indirectly induces secondary effects by ionizing water molecules to produce reactive oxygen species (ROS) [2,3]. ROS oxidize lipids and proteins and also induce various forms of DNA damage, including the generation of oxidized bases and single-strand breaks (SSBs) in the radiated cells/tissue, ultimately leading to cellular apoptosis [4]. Furthermore, IR also promotes clustered DNA damage and induces covalent inter- and intra-strand cross-linking [5]. Additionally, there is a growing recognition of the immune response generated by RT. This review will focus on the implications of the innate myeloid and lymphoid lineages in anti-tumorigenic processes induced by RT and the use of various types of RT methods in cancer treatment [6,7]. Additionally, we also explore key strategies for enhancing the efficacy of RT while maintaining innate immunity during cancer therapy.
RT is an essential component of personalized medicine. It promotes immune suppression by inducing toxicity in bone marrow cells [8] and peripheral blood lymphocytes [9]. Additionally, RT activates innate immune systems and leads to bystander effects [10]. In the peripheral blood, dendritic cells, macrophages, and NK cells play a crucial role in regulating innate immunity and determining the efficacy of radiation therapy.
2. Innate Immunity
The immune system is classified into two categories: innate and adaptive immunity. Innate immunity is largely composed of myeloid/macrophages, natural killer (NK) cells, and dendritic cells. The innate immune system constitutes the first line of defense against invading microbial pathogens and recognizes the pathogens through pattern recognition receptors (PPRs) [70,71,72,73,74]. PPRs can detect conserved structures on pathogens termed pathogen-associated molecular patterns (PAMPs) [71]. However, recent findings suggest that the induction of immune effectors also commonly occurs in the absence of pathogen infection, which is termed sterile inflammation. Sterile inflammation is commonly found in RT-induced innate immune responses. PPRs also detect Damage Associated Molecular Patterns (DAMPs) [75,76] that originate within the damaged cell itself. The innate immune system initiates an immune response following the detection of DAMPs, which signals the status of tissue or cell damage or danger events. Innate immunity is activated by antigens and different immune cells, including dendritic, mast, natural killer (NK) cells, macrophages, monocytes, and granulocytes, to maintain the immune system. Adaptive immunity is mediated by lymphocytes such as T and B cells and is characterized by immunological memory cells that allow a long-lasting response. The effect of RT on adaptive immunity has been extensively discussed in the literature.
Role of RT in Priming the Innate Immune Response
RT induces apoptosis, which triggers DAMPs. Examples of DAMPs include the extracellular release of high mobility group box1, production of cytokines such as type I interferon (IFN-1), release of nuclear (nDNA) and mitochondrial DNA (mtDNA) to cytoplasm, and production of reactive oxygen species (ROS) or free radicles. These signals induce a series of chemical and immunological reactions that affect immunity. Mitochondria contain numerous potent immunostimulatory DAMPs, including mitochondrial DNA (mtDNA), ATP [77] and ROS. Mitochondrial DAMPs engage the innate immune macrophages or neutrophils [78] upon release to the cytosol or into the extracellular environment. The release of mtDNA into the cytosol activates PPRs to trigger a variety of innate immune responses. One such PPR is the DNA sensor cyclic GMP-AMP (cGAM) synthase, which binds cytosolic double-strand DNA (dsDNA) derived from mitochondria. This results in the generation of the second messenger cGAMP and activates the cGAS-STING pathway at the endoplasmic reticulum, leading to the recruitment of the tank binding kinase 1 (TBK1) and activation of the IFN signaling pathway [79]. Another DAMP is mitochondrial ATP, the key transporter of chemical energy. Recently, in several models, it has been shown that IR causes the release of ATP from tumor cells and activates DC cells [80]. ATP binds to P2X7 on DC cells, leading to the activation of NLRF3 inflammasomes [81].
3. Mechanisms of Radiation-Induced Innate Immune Cell Activation
3.1. Dendritic Cells
Myeloid cells constitute a highly diverse population that is comprised mainly of dendric cells (DCs), monocytes and macrophages [82]. Dendritic cells play a crucial role in host immunity by promoting innate inflammatory responses to environmental or damage stimuli [83]. TNF-α and IL-1β are proinflammatory signaling molecules that are upregulated in response to IR. These molecules, in turn, activate antigen-presenting innate immune cells, including dendritic cells [84,85,86,87,88]. Dendritic cells are specialized antigen-presenting cells that play a crucial role in T-cell activation following RT-induced damage in tumor cells. Dendritic cells recognize DAMPs via specific receptors and matured dendritic cells [89] stimulate cytotoxic CD8+ T cells by antigen presentation and release of activating cytokines, thereby enhancing RT treatments. The intensity of radiation doses and the number of doses determines the immunogenic action of dendritic cells in RT. For example, repeated low radiation doses in a murine mammary carcinoma model create cytosolic DNA in tumor cells, activating the cGAS-STING pathway and the release of IFN-γ and subsequent T-cell activation [72,90,91] (Figure 1A,B). RT sensitivity depends in part on DNA exonuclease called 3′ repair exonuclease 1 (Trex1). Trex1 cleaves the RT-induced cytosolic DNA, thereby abrogating IFN-β production through the cGAS-STING pathway. The Trex1 level does not increase in response to multiple smaller fractions of radiation (8 Gy, three times); rather, it induces greater IFN-β production and activation of Bat3-dependent dendritic cells, leading to enhanced T cell responses. Compared to a single fraction of high-dose radiation, the induction of Trex1 in multi-low-dose RT is efficient and suggests that a fractionated low-dose of RT likely plays a role in enhancing immunogenicity [90,92].
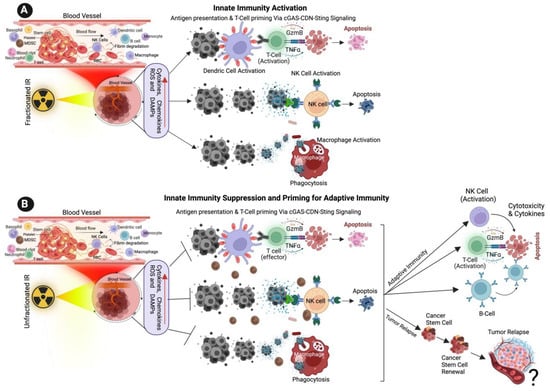
Figure 1. Effect of Ionizing Radiation on Innate Immune Activation. (A) The illustration shows the low dose of fractionated RT induces the production of ROS, cytokines, chemokines, and damage-associated molecular patterns (DAMPs) from cancer cells, DAMPs promoting the activation of innate immune dendritic cells, nature killing cells (NK cells) and macrophages. Activated dendric cells present tumor antigens and are primed for T-cell activation. Activated NK cells promote cytotoxic effects on tumor cells, and activated macrophages destroy cancer cells via active phagocytosis. (B) The cartoon illustration shows that a high dose of unfractionated radiation induces the production of ROS and DAMPs from cancer cells, inactivating innate immune dendritic cells, macrophages, and NK cells, promoting immune escape and tumor relapse. Unfractionated radiation also promotes adaptive immune cell activation via T-cells, NK cells and B-cells and promotes intra-tumoral immunity. ? indicates may or may not happen, arrow indicates the next step.
3.2. Natural Killer Cells
NK cells effector lymphocytes play a crucial role in regulating innate immune responses, combating microbial infections, and controlling cancer. While IR has been shown to have a significant impact on NK cells, the underlying mechanisms behind this effect remain unclear [93]. NK cells are innate immune lymphocytes that can destroy target tumor cells by producing toxic and immunoregulatory cytokines [94,95]. IR has a significant effect on modifying NK cells. Previous studies have demonstrated that IR enhances the immune response by augmenting the antigenicity and adjuvanticity of malignant cells and by interacting with the tumor microenvironment (TME) [96]. Low-dose ionizing radiation activates NK cell functions, while high-dose ionizing radiation particularly impairs NK cell function (Figure 1A,B); however, this impairment can be reversed by interleukin-2 (IL-2) pretreatment [97,98,99]. Low-dose ionizing radiation at 75–150 mGy increases the secretion of NK cell effector proteins, such as IFN-γ and TNF-α [93,100]. Similarly, low-dose total-body irradiation (0.1 or 0.2 Gy X-ray) results in the suppression of experimental tumor metastases along with the stimulation of NK cell cytolytic functions in tumor-bearing rates [101,102].
Studies have identified that low-dose ionizing RT (LDIRT) can increase the immune response in vivo [100] with IFN-γ and TNF-α in the cultured medium of NK cells in response to LDIRT, and in addition, the P38 inhibitor (SB203580) drastically suppressing the NK cell cytotoxicity, cytokine levels, FasL and perforin [100,103]. Ames et al. (2015) identified that ex vivo NK cells are activated following low dose IL-2 and IL-15 and presented an increased ability to mitigate solid tumor cells in vitro and in vivo following RT [104]. A similar study reported that the presence of the cytotoxic effect in NK cells was boosted following RT in canine models of sarcoma, and the results from a clinical are progressing with possible abscopal effects. In general, NK cells produce perforin (Prf1) and granzyme B (GzmB) and induces cancer cell apoptosis (Figure 1) [105,106]. It is also observed that dendritic cells (DC) activate NK cells and promote tumor cell apoptosis [107]. A recent study utilized a reverse translational approach and revealed that NK cells play a role in immune enhancement through a CXCL8/IL-8-dependent mechanism in response to radiation [108]. Furthermore, the study suggests that NF-κB and mTOR mediate the secretion of chemokines that facilitates the infiltration of NK cells into tumor cells. Additionally, the study highlights that higher doses of radiation promote the transfer of adoptive NK cells and improve tumor control [108].
3.3. Macrophages
Macrophages play a crucial role in various aspects of immunity, including infiltrating the TME. Macrophages belong to the myeloid family and originate from circulating bone marrow-derived monocyte precursors. Macrophages are highly plastic cells that undergo significant changes in their function depending on the environmental cues in the TME, exerting a dual function on tumorigenesis by either antagonizing the cytotoxic activity of immune cells or enhancing the antitumor responses (Figure 1A,B). Macrophages are classified into two different phenotypes, M1 and M2. M1 macrophages are called classically activated macrophages in response to pathogens and take part in the immune response. M2 macrophages are known as alternatively activated macrophages involved in wound repair and have an anti-inflammatory role. Following recruitment, the monocyte precursor cells differentiate into macrophages in the tissue. The matured macrophages polarize to functionally different phenotypes in response to microenvironmental challenges in TME in tumor cells. Tumor-associated macrophages (TAMs), a major stromal component of TME, resemble the M2-polarized macrophages [109,110,111]. M1 macrophages are also involved in antitumor immunity, while M2 macrophages exert pro-tumorigenic activities.
Macrophages are recruited to the damaged or injured sites following the radiation exposure, where they carry out their phagocytic function [112,113,114]. Macrophage responses to RT range from promoting tumor growth to enhancing the immunogenic response, depending on the tumor type, environment, IR and dose, and fractionation. Inducible nitric oxide synthase (iNOS)+ M1-like macrophages undergo differentiation in response to local low-dose ionizing radiation, allowing the recruitment of tumor-specific T-cells and tumor regression in human pancreatic carcinomas [110,115]. Irradiated cells induce cytokine secretion and hypoxia within the damaged tissue, and the activation of the transcription factor HIF1-α has been shown to contribute to the recruitment of macrophages towards the immunosuppressive phenotype. Activated macrophages can directly destroy cancer cells by enhancing the phagocytosis of tumor cells through antibody-dependent cellular cytotoxicity. Alternatively, by secreting toxic/harmful molecules such as cytokines or tumor necrosis factors TNF or nitric oxide and promote cytolysis of cancer cells. The indirect killing of tumor cells involves the recruitment of immune cells, such as cytotoxic T-cells (Figure 1) [116,117,118].
This entry is adapted from the peer-reviewed paper 10.3390/cancers15153972
This entry is offline, you can click here to edit this entry!