Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Food Science & Technology
蘑菇生物活性肽(MBP)是直接或间接从食用蘑菇中提取的生物活性肽。众所周知,MBP具有抗氧化,抗衰老,抗菌,抗炎和抗高血压的特性,并促进记忆和认知改善,抗肿瘤和抗糖尿病活动以及降低胆固醇。
- edible mushrooms
- preparation
- bioactivities
1. 简介
蛋白酶水解蛋白质产生具有特殊生理功能的短序列氨基酸,称为生物活性肽[1]。生物活性肽大多是从各种动物和植物中分离出来的。生物活性肽通常从动物来源获得,例如牛奶和乳制品[2],鸡蛋[3],肉类[4]和鱼类衍生产品[5]。来自植物的生物活性肽主要来自豆类[6],谷物[7],坚果[8],水果和蔬菜[9]。到目前为止,关于从动植物中分离的生物活性肽的大量研究已被大量报道,而对食用菌生物活性肽(MBPs)的研究相对较少。目前,至少有12,000种蘑菇,其中2000种据报道是可食用的,约200种作为食品或药物成分收集[10]。因此,食用菌在相对未开发的资源中丰富而广阔。从不同种类的蘑菇中分离出的不同生物活性化合物可用于开发不同的功能性食品。食用蘑菇含有大量的生物活性代谢物,包括蛋白质、多糖、酶(例如超氧化物歧化酶)、膳食纤维和许多其他生物分子,因此是极好的功能性食品和营养补充剂[11]。例如,杂色曲霉通常以片剂、胶囊和粉末的形式作为膳食补充剂销售[12]。最近,杂色Trametes 中的天然风味化合物已成为生产无酒精或低酒精啤酒的宝贵成分[13,14]。根据美国农业部(https://fdc.nal.usda.gov/(8年2020月15日访问))的统计数据,可以看出食用菌的蛋白质含量高于大多数蔬菜,许多蛋白质具有生物活性,而更多甚至新的MBP仍有待发现[16,1]。MBPs可以被肠道完全吸收,在不增加胃肠道功能负担的情况下,在消化道中产生直接的部分益处[17]。MBPs可以通过以完整状态进入循环系统来发挥有益的生理作用。MBP可以提供高组织亲和力[18],低毒性和高稳定性[19],具有一系列营养,功能和生物学特性。内源性MBP可以通过调节抗氧化途径和激活免疫细胞来发挥抗氧化和抗炎活性。MBP也可作为镇痛药或阿片类药物,改善神经细胞的记忆力和认知能力恶化[20]。外源性MBP可用作功能性食品和药物,以改善炎症、高血糖、高血压、高胆固醇和其他退行性疾病[21]。最近的研究发现,双孢蘑菇生物活性肽作为合生元制剂中的新一代益生元和益生菌微生物具有良好的健康作用[22]。对于不喜欢食用动物源性产品的消费者来说,MBP可以成为替代动物源性食品的理想替代补充剂的来源[23]。用健康的天然食品代替一些肉类或鱼类是未来市场的趋势。食用菌的味道与肉制品相似,营养丰富。出于这个原因,许多企业家在肌肉食物(例如肉类和鱼类)中加入了食用蘑菇,以减少肉类的比例[<>]。
2. MBP的制备
食用菌是天然生物活性蛋白质和肽的重要来源,为MBPs的发现提供了良好的物质基础。食用菌副产物的蛋白质含量约为25%,从食用菌副产品中提取的生物活性肽可以回收用于生产功能性食品,可有效减少资源浪费[24]。由于食用菌的生长环境和种类不同,提取方法多种多样。因此,MBP的结构和生物活性也各不相同[9]。基本上,有两种流行的方法来制备 MBP;一种是直接从蘑菇中提取内源性MBP。许多内源性MBPs直接从新鲜子实体、干粉或发酵粉中提取,例如分别具有抗菌和ACE抑制特性的肽[25]。另一种方法是使用外源酶的蛋白水解反应,从食用蘑菇或其菌丝体中分离的蘑菇蛋白中间接释放肽片段(例如,菠萝蛋白酶水解),以从完整蛋白质中释放强效肽[9]。事实上,具有多种生物活性的MBPs,如抗氧化、抗菌、抗炎、抗衰老、抗肿瘤和抗糖尿病活性,都是通过酶水解获得的,例如,香菇与碱性酶的水解具有ACE抑制作用的肽[26]。MBP含有麦角硫因、漆酶、核糖核酸酶和其他酶[27]。发现在超高压处理辅助下获得的低分子量MBPs在体外激活乙醇脱氢酶和醛脱氢酶。它们能有效催化酒精逐渐转化为毒性较低的乙醛和乙酸,预防酒精性肝损伤[15]。
化学水解也是制备MBP的常用方法之一。 化学水解涉及使用酸或碱溶液破坏蛋白质的肽链,产生肽和游离氨基酸。食品工业中的化学水解具有多种局限性,例如难以控制过程,营养物质损失和环境污染[28]。微生物发酵也是近年来常用的获取MBP的方法之一。微生物具有蛋白质水解系统,可以产生蛋白酶或诱导发酵以获得MBP。MBP的类型取决于发酵时间、菌株和食用蘑菇蛋白的类型[29]。微生物发酵可以避免酶消化产生的肽中的杂质,缩短制备时间,降低生产成本。
超滤、亲水作用色谱(HILIC)、快速蛋白液相色谱(FPLC)、离子交换色谱(IEC)、体积排阻色谱(SEC)、凝胶过滤色谱(GFC)或高效液相色谱(HPLC)[25]是提取和初步生物活性筛选后纯化常用的方法。例如,使用SEC、反相高效液相色谱(RP-HPLC)和液相色谱-质谱(LC-MS/MS)从双孢蘑菇中分离并表征抗高血压肽[25]。
MBP可缓解与靶向损伤或退行性疾病相关的症状。此外,已知MBPs通过灭活活性氧和清除自由基来发挥抗氧化作用,以改善退行性疾病,如炎症,高血压,高血糖,心血管疾病,高胆固醇,记忆和认知疾病,具有抗氧化,抗菌,抗衰老和抗肿瘤的作用(图1)。
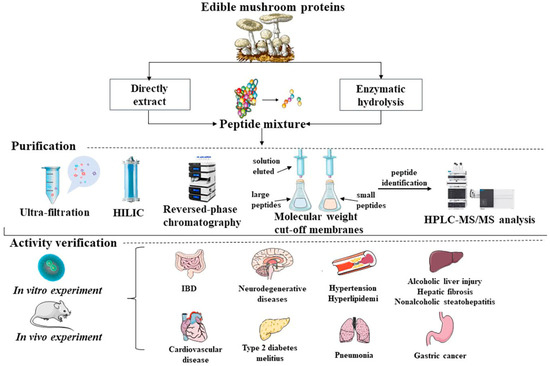
图1.常见的提取途径,纯化和初步生物活性筛选,体内和体外验证MBPs的生物活性。
3. MBP的生物活性
3.1. 抗氧化活性
食用菌富含多种具有抗氧化活性的肽[30]。由于原料和制备工艺的不同,MBP可能具有不同的靶向功能。MBPs的抗氧化机制主要分为ROS产生调控、抗氧化酶活性调控和抗氧化途径调控。MBPs可以通过提供质子、电子和螯合金属离子来清除自由基,以调节ROS的产生[31]。从双孢蘑菇(ABP)和杏鲍菇菌丝体(PEMP)中分离的低分子量肽富含带负电荷的氨基酸,可以中和自由基并调节ROS的产生[32]。G. 灵芝肽 (GLP) [33] 通过以剂量依赖性方式阻断大豆脂氧合酶活性,在大豆油系统中发挥抗氧化作用,使用 IC50值为 27.1 微克/毫升。与丁基化羟基甲苯相比,GLP通过清除生物系统中的羟基自由基和淬灭超氧自由基阴离子具有更好的抗氧化活性。
生物体可以通过内源性酶和非酶防御系统及时调节ROS的产生[34]。口服GLP通过抗氧化活性对肝损伤小鼠发挥了显著的保肝作用[35]。GLP增加了谷胱甘肽(GSH)和超氧化物歧化酶(SOD)的水平,降低了肝脏中丙二醛(MDA)的水平。此外,它降低了血清中丙氨酸转氨酶(ALT)和天冬氨酸转氨酶(AST)抵抗肝纤维化和酒精性肝损伤的活性。研究表明,GLP通过调节抗氧化酶的活性,能有效抑制线粒体中产生的过氧化物的危害,表现出优异的抗氧化活性。
MBP通过调节抗氧化途径来实现抗氧化活性,例如Kelch样ECH相关蛋白1-核因子红细胞-2相关因子2-抗氧化响应元件(Keap1-Nrf2-ARE)、活化B细胞的核因子-κ轻链增强子(NF-κB)、丝裂原活化蛋白激酶(MAPK)和磷酸肌醇3激酶/蛋白激酶B(PI3K/AKT)途径[36,37,38].MBPs可以通过下调Keapl基因和上调Nrf2基因表达来调节相关抗氧化蛋白的表达(图2)[31]。GLP [39]促进Nrf2并激活Nrf2-ARE信号通路,对过氧化氢诱导的细胞表现出抗氧化作用(H2O2).Nrf2在刺激PI1K / AKT和MAPK途径时与Keap3解离,移动到细胞核,并与抗氧化成分ARE相关,其调节抗氧化酶如HO-1,过氧化氢酶(CAT)等的表达。[40]。
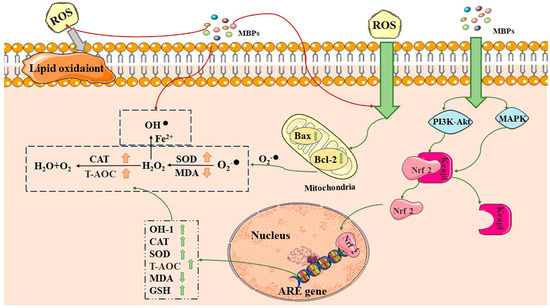
Figure 2. Antioxidant and anti-aging mechanisms of MBPs. MBPs may exert antioxidant and anti-aging effects by regulating Keap1-Nrf2-ARE through the PI3K/AKT and MAPK pathways. CAT: catalase, SOD: superoxide dismutase, T-AOC: total antioxidant capacity, MDA: malondialdehyde, GSH: glutathione, Bax: BCL2-Associated X, Bcl-2: B-cell lymphoma-2. Excess ROS by generating superoxide anions via Bax and Bcl-2 in cellular mitochondria. MBPs regulate ROS content by providing protons, electrons and chelating metal ions. MBPs regulate the Keap1-Nrf2-ARE signalling pathway through the PI3K/AKT and MAPK pathways. Red arrows indicate that MBPs regulate ROS content by providing protons, electrons, and chelating metal ions. The grey arrows indicate that excess ROS are harmful by generating superoxide anions. Green arrows indicate that MBPs and ROS react via different pathways within the cell.
Anti-Aging Activity
Aging occurs in cells, organs and the whole organism, which leads to a reduction in biological function of the organism’s ability to eliminate oxidative stress. The accumulation of excess free radicals causes MDA levels to increase, the total antioxidant capacity (T-AOC) to reduce, and disrupt cellular structure [41], leading to cellular senescence and death. CMP [42] and GLP [43] had dose-dependent scavenging effects on oxygen free radicals and hydroxyl free radicals. SOD is a critical mitochondrial enzyme antioxidant associated with longevity [44]. MBPs could significantly enhance senescence-associated mitochondrial enzyme antioxidants. CMP and GLP scavenged hydroxyl radicals better than the specific hydroxyl radical scavenger mannitol [45]. Agaricus blazei peptide (ABp) reduced MDA and ROS contents, and increased CAT and T-AOC activities in a D-galactose-induced senescence model of the NIH/3T3 cell [46].
The main targets of MBPs to slow down the aging process include the metabolic mitochondrial pathway, inactivating reactive oxygen species, scavenging free radicals, alleviating the oxidation of biomarkers in organisms, and reconstructing homeostatic mechanisms in vivo [47]. Nrf2 activity was tightly correlated with degenerative diseases induced by aging, and contributed to the prevention and mitigation of degenerative diseases. In the aging model established by D-galactose in mice, feeding ABp was found to lead to the down-regulation of Keap1 protein expression, thus up-regulating Nrf2. In the Keap1–Nrf2 pathway, the expressions of HO-1 and related factors such as ApoE, Hsph1 and Trim32 were up-regulated, effectively scavenging free radicals and showing excellent anti-aging activity [48]. Progressive changes were found in the epigenetic information in both dividing and non-dividing cells in modifications involving chromatin transformations, histone and DNA methylation patterns in research related to aging. The major features of aging are constituted in DNA and histones methylation, accompanied by other epigenetic alterations [49]. In previous studies, it was found that the epigenetic alterations of aging could be modulated by the consumption of peptides that directly maintain telomere length [44]. Current research about whether MBPs could regulate telomere length in different organisms has not been conducted. This would be a new area of research on the anti-aging activities of MBPs, and more relevant research is needed in the future.
In conclusion, MBPs can act on multiple targets simultaneously to exert their functional antioxidant and age-delaying activities. MBPs have a wide scope for development as one of the components of natural antioxidant and anti-aging functional foods. Currently, MBPs have become a new direction of research on functional foods. However, further research is needed on how MBPs replace synthetic antioxidants.
3.2. Antimicrobial Activity
Antibiotics can treat infections such as tuberculosis, pneumonia, leprosy and gonorrhoea. However, antibiotic resistance [54,55] has emerged from the overuse and misuse of antibiotics. Natural bioactive peptides are characterised by their high efficacy, stability and low toxicity, making them major alternatives to antibiotics and conventional drugs [56]. MBPs have been widely studied for their antimicrobial activities. A variety of antimicrobial peptides have been isolated and purified from edible mushrooms such as Polyporus alveolaris, Pleurotus eryngii [57], Lentinus edodes and Agrocybe cylindracea [58]. The possible antimicrobial mechanisms of MBPs could involve either regulating and leading to tissue-specific expression patterns, or making intracellular protein leakage and leading to bacterial death.
The hydrophobic amino acids of MBPs could regulate the NF-κB pathway and the MAPK pathway. PEMP [32] had good antibacterial activity by stimulating macrophage proliferation, increasing phagocytosis activities, TLRs expression, and releases of tumour necrosis factor-α (TNF-α), IL-6, NO and H2O2. MBPs are rich in structures with α-helices, β-folds, random coils and disulfide bonds [59]. Antimicrobial peptides are enriched with hydrophobic amino acids, α-helixes, β-folds, random coils and disulfide bonds.
Another antibacterial mechanism of MBPs is the disruption of bacterial cell membranes, causing intracellular protein leakage within the bacterial cells to achieve antibacterial effects. Antimicrobial peptides from the mycelia of Cordyceps militaris [60] may cause intracellular protein leakage of E. coli (ATCC 25922), which maintained the integrity of the intestinal mucosa and attenuated E. coli infections in mice. Antibacterial peptides isolated from the mycelia (GLM) and fruiting bodies (GLF) of G. lucidum [61] showed a dose-dependent increase at 50–125 μg/mL in protein leakages from Escherichia coli and Staphylococcus aureus. It is clear that GLF and GLM induces cell death with strong antibacterial activities against both Escherichia coli and Staphylococcus aureus.
Based on the above, MBPs show good antibacterial activities against bacteria that are resistant to drugs. Thus, MBPs could be important natural alternatives to antibiotics.
3.3. Anti-Inflammatory Activity
MBPs may enhance the cytotoxicities of natural killer cells and the phagocytosis of macrophages, promoting multiplication and maturation of immune cells and lymphocytes and inhibiting pro-inflammatory responses, thereby improving the host’s defence against invading pathogens (Figure 3) [69]. Yu [70] prepared a bioactive peptide, KSPLY, with a molecular weight of 608.3834 Da, from Hericium erinaceus. KSPLY promoted TNF-α, NO, IL-6 and IL-1β secreted by macrophages, which inhibited lipopolysaccharide (LPS)-induced inflammatory responses at a concentration of 100 μmol/L. The proliferation of splenic lymphocytes was significantly reduced in mice fed with Pleurotus eryngii peptide (PEP), and the serum haemolysin level of CTX-induced mice was significantly increased [71]. This confirmed that PEP could significantly improve humoral immune function of immunosuppressed mice.
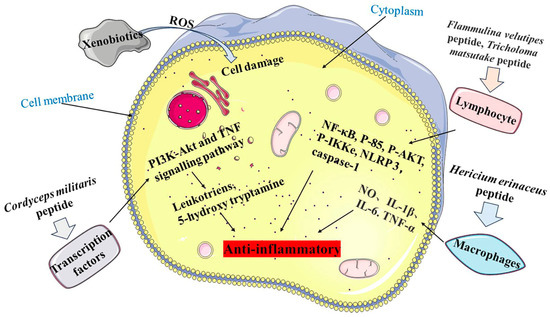
Figure 3. Anti-inflammatory mechanism of MBPs. Pathway 1 is Cordyceps militaris peptide regulating the PI3K-Akt and TNF signalling pathways to release Leukotriens and 5-hydroxy tryptamine. Pathway 2 is Flammulina velutipes peptide and Tricholoma matsutake peptide acting on lymphocytes, which in turn regulate NF-κB, P-85, P-AKT, P-IKKe, NLRP 3 and caspase-1. Pathway 3 is Hericium erinaceus peptide acting on macrophages to release NO, IL-1β, IL-6 and TNF-α.
MBPs may enhance the oxidative defensive system and barrier function by promoting the production of antibodies, cytokines and chemokines to reduce the inflammatory response. After local nasal immunotherapy (LNIT) with Flammulina velutipes peptides (FIP-fve), the production of proinflammatory cytokines and chemokines were significantly reduced [72]. It has been shown that FIP-fve induces the potent activator of peripheral lymphocytes through activation of the p38 mitogen-activated protein kinase (p38 MAPK) signalling pathway with anti-inflammatory activity. The Tricholoma matsutake-derived peptide WFNNAGP [53] attenuated the inflammatory response by inhibiting the expression of pro-inflammatory cytokines and myeloperoxidase (MPO), as it promoted expressions of tight junction proteins closing ribbon-1, claudin and occluding. WFNNAGP reduced colonic inflammation in mice by down-regulating NF-κB expression to inhibit formation and activation of NLRP 3 and caspase-1.
MBPs have a variety of target cells and sites, such as NK cells, CD4, CD25, T lymphocytes, macrophages [73], monocytes, B lymphocytes [74] and mast cells [75], among others. The results of a study showed that FIP-fve could make changes in Treg-associated immunity, with down-regulation of IL-4/CD4 T-cell expression and up-regulation of IFN-γ/CD4 T-cell expression in mice. FIP-fve effectively decreased inflammatory cell infiltration and epithelial damage [72]. Experiments demonstrated that the anti-inflammatory effect of orally administering FIP-fve on mite-induced airway inflammations in mice. Bioactive polypeptides of Cordyceps militaris (CMP) [76] were implicated in the regulation of immune function in mice through transcription factors Ets1 and the Spp1, Rel, and Smad3 genes. CMP regulated TNF and the PI3K-Akt signalling pathway, playing an important role in inflammation by increasing immune organ indexes, the number of leukocytes and the content of hemolysin in the sera of mice. In summary, MBPs have good anti-inflammatory activity, and offer new ideas for the development of functional food supplements as natural ingredients.++++++
3.4. Memory and Cognitive Improvement Activity
Medications that are commonly used clinically to prevent and improve learning in learning and memory impairments include free radical scavengers, acetylcholinesterase (AChE) inhibitors and drugs to prevent the formation of amyloid β deposits [77]. However, these types of drugs are associated with adverse side effects, high toxicity, poor memory improvement and no preventive effect on the onset and development of neurological disorders. CMP scavenges oxygen free radicals, decreasing AChE activity in the mouse brain, which may reduce the damage of cholinergic neurological nerves, hence improving learning and memory ability in a scopolamine-induced mice model of learning and memory impairment [78]. The mechanism of CMP is similar to that of free radical scavengers and AChE inhibitors. CMP promotes the expressions of Il-1β and Slc18a2, as well as secretion of neurotransmitters in mice that reduce dyskinesia and depression-like behavior, and contributed to improve learning and memory. Hericium erinaceus [79] fed to mice prevented the loss of spatial short-term and impairments of visual recognition memory induced by the formation of amyloid β deposits. However, there are relatively few studies on improvements in memory and cognitive ability by Hericium erinaceus peptides, and more experiments are needed to verify its mechanism in the future.
Based on the above, MBPs may effectively prevent and improve learning and memory disorders, with have the advantages of fewer side effects, less toxicity, more targets of action and lower costs. MBPs may be more effective in preventing memory and cognitive disorders compared with synthetic drugs.
3.5. Anti-Hypertensive Activity
Hypertension is currently one of the most common metabolic diseases, and is a predisposing factor for many other conditions such as renal failure and heart disease. The inhibition of angiotensin-converting enzyme (ACE) regulation of the renin–angiotensin system is thought to be the main mechanism of hypotensive activity [80]. Edible mushrooms are good sources of hypotensive peptides, and many antihypertensive peptides have been isolated. Kaprasob [81] found that LIYAQGFSK peptide extracted from the King Boletus mushroom had the greatest ACE-binding energy of −9.2 kcal/mol through hydrogen bonds, and exhibited ACE inhibitory activity. The AHEPVK, RIGLF and PSSNK peptides of ABP showed stronger ACE inhibitory activities with lower IC50 values after gastrointestinal digestion compared to ACE inhibitory peptides (ACEIPs) from other sources [25]. MBPs inhibit ACE mainly through the inhibition of nucleic acid synthesis or binding, blocking protein synthesis, membrane permeabilization, inhibition of enzyme activities and triggering apoptosis. Gln-Leu-Val-Pro (QLVP) is a polypeptide with its amino acid sequence isolated from the mycelia of Ganoderma lucidum. QLVP significantly activated the angiotensin I-mediated phosphorylation of endothelial nitric oxide synthase in human umbilical vein endothelial cells, and partially reduced mRNA and protein expressions of vasoconstrictor factor endothelin-1, which showed value in the application of GLP for treating hypertension in metabolic diseases. Overall, MBPs exhibit good anti-hypertensive activities as potential ingredients for natural anti-hypertensive functional foods and nutraceuticals.
3.6. Antitumour Activity
Biological therapy is an extensively alternative approach to limit the growth of cancer cells that uses biologically active compounds to treat cancer. Bioactive peptides in food can inhibit cancer at all stages of the disease, and offer advantages such as their greater affinity, target-specific effects, reducing toxicity and superior tissue penetration, in comparison to the side effects of chemotherapeutic molecules [87]. PEP was found to inhibit the growth of cancer cells in concentrations of 0.05 to 2 mg/mL. It inhibited the growth of gastric cancer (HGC-27), breast cancer (BT-549) and cervical cancer (Hela-229) cells by 61.40%, 59.20%, and 32.80%, respectively, and demonstrated strong antitumour activity. The MBPs exerted anti-tumour activity by stimulating the host′s defensive mechanism to enhance its non-specific immune responses. CMP [42] activated the reticuloendothelial system and macrophages to promote lymphocyte transformation and immunologically active cells like lymphocytes, lymphokines, monocyte macrophage systems and NK cells to attack target cells to have anti-tumour effects. The amino acid sequence of the Ser-Leu-Ser-Leu-Ser-Val-Ala-Arg peptide extracted from morels (Morchella spp.) were found to reduced cell proliferation via a mitochondrial-dependent pathway. It was shown that the movement of cytochrome C from the mitochondria to the cytoplasm could be facilitated by the down-regulation of Bcl-2/Bax, thereby promoting the expression of caspase-9 and caspase-3 and suppressing tumourigenesis [88]. MBPs facilitated intermediary metabolism or controlled DNA transcription and translation through the activation of related enzyme systems. Mice fed with CMP [89] were found to significantly inhibit polymorphonuclear cells infiltration and the infrared ray-induced up-regulation of brain production of C3 protein levels, and the production of interleukin-1β and tumour necrosis factor-α. MBPs exhibit antitumour effects, and may be promising natural tumour preventive agents.
3.7. Other Activities
In addition to their antioxidant, antibacterial, anti-inflammatory, anti-aging, memory and cognitive improving, anti-hypertension and anti-tumour activities shown above, MBPs also have functional activities, such as reducing the levels of plasma glucose with induced diabetes, and decreasing cholesterol levels.
Diabetes is a metabolic disease that is characterised by hyperglycaemia. Prolonged uncontrolled hyperglycaemia in an organism can generate oxidative stress, promote apoptosis, activate protein kinase isozymes and transcription factors, and other pathophysiological mechanisms that impair the proper functioning of tissues and organs [90]. Currently, glycaemia is mainly controlled by the intramuscular injection of hormones or taking synthetic anti-diabetic agents such as insulin, thiazolidinediones, dipeptidyl peptidase 4 inhibitors, biguanides, sulfonylureas and sodium–glucose cotransporter type 2 inhibitors [91]. These hypoglycaemic agents are highly effective, but with side effects [92]. Compared with other synthetic hypoglycaemic substances, MBPs show fewer side effects, and may regulate glycaemia by inhibiting the activities of α-amylase, α-glucosidase and dipeptidyl peptidase-IV. At 2.0 mg/mL, a morel peptide inhibited α-amylase and α-glucosidase by 34.93% and 30.56%, respectively [93], showing excellent anti-diabetic activity.
Hypercholesterolaemia arises from increased endogenous cholesterol synthesis or an excess supply of dietary cholesterol or cholesterol precursors [94]. Bioactive peptides may inhibit the synthesis and secretion of triglycerides and cholesterol by stimulating bile acid secretion from the gallbladder, modulating hormones and cholesterol receptors, and altering hepatic lipid metabolism [94]. MBPs are rich in inhibitors of HMG-CoA reductase, a pivotal enzyme in endogenous cholesterol biosynthesis, as well as ergocalciferol, eritadenine and β-glucan derivatives. MBPs also have a similar mechanism of action to simvastatin or ezetimibe, which are statins [95]. The hydrophobic amino acids of Phe, Leu, Val, Gly and Pro in bioactive peptides may promote cholesterol homeostasis in organisms by stimulating the elimination of exogenous cholesterol during gastric emptying [96]. ABP prepared by Feng [97] reduced the solubility of cholesterol by forming micellar structures in a manner similar to the cholesterol-lowering mechanism of statins. All functional foods with MBPs may lower serum cholesterol levels. Unlike other food derivatives, they may regulate cholesterol homeostasis in organisms through different transcriptional and translational mechanisms. These mechanisms are not completely elucidated, and may be different from those of plants that have already been identified [95].
This entry is adapted from the peer-reviewed paper 10.3390/foods12152935
This entry is offline, you can click here to edit this entry!