Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
MT4-MMP (or MMP-17) belongs to the membrane-type matrix metalloproteinases (MT-MMPs), a distinct subset of the MMP family that is anchored to the cell surface, in this case by a glycosylphosphatidylinositol (GPI) motif. Its expression in a variety of cancers is well documented.
- MT4-MMP
- MMP-17
- development
- vasculature
- tumor
1. Introduction
Matrix metalloproteinases (MMPs) are a family of zinc-dependent endopeptidases that are able to degrade and remodel the extracellular matrix (ECM) by the cleavage of distinct matrix components and also to lead to the proteolytic activation/inactivation of receptors, growth factors, adhesion molecules, cytokines, and other pericellular proteins [1][2]. Included within the MMP family are the membrane-type matrix metalloproteinases (MT-MMPs) that are anchored to the cell membrane by a type I transmembrane domain (MT1-MMP/MMP-14, MT2-MMP/MMP-15, MT3-MMP/MMP-16, and MT5-MMP/MMP-24) or by a glycosylphosphatidylinositol (GPI) anchor (MT4-MMP/MMP-17 and MT6-MMP/MMP-25) [1][3][4].
Membrane-type 4 MMP (MT4-MMP, also known as MMP-17) is anchored by a GPI motif to the plasma membrane, which confers exclusive mechanisms of biosynthesis and regulation. Apart from this, MT4-MMP conserves the three structural domains characterized in all MMPs: the prodomain, the catalytic, and the hemopexin domains. The prodomain, with a length of 80 amino acids and a consensus sequence with unpaired cysteines, keeps the enzyme in a latent state, named zymogen. The catalytic domain is located at the C-terminal of the prodomain. It has a conserved sequence (“HEXXHXXGXXH”) that includes a Zn2+ ion binding motif which is essential for the proteolytic activity of the proteinase. This is linked by the hinge region to the hemopexin domain involved in substrate recognition and degradation. Next, MT4-MMP displays a small region called the stem region which has two cysteines. These residues appear to be involved in the dimerization and oligomerization of MT4-MMP [5] in a similar manner to that described for the other GPI-anchored metalloproteinase (MT6-MMP) [4]. Once MT4-MMP is exposed at the cell surface, its enzymatic activity is regulated by endogenous inhibitors, tissue inhibitors of metalloproteinases (TIMPs). There are four mammalian TIMPs (TIMP-1, 2, 3, and 4) that inhibit MT-MMPs by binding their N-terminal domain with the catalytic zinc ion of the enzyme. MT4-MMP is inhibited by all TIMPs, among which TIMP-1 is the most effective inhibitor [2][4][6].
Interestingly, the hemopexin domain of MT4-MMP only displays a 40% similarity with the same domain of the other family members [7]. This feature may explain the specificity and exclusivity of the substrates to be cleaved by MT4-MMP and potentially its internalization mode [1][5][8]. In fact, only a limited number of substrates have been identified for this endopeptidase, including matrix and matricellular proteins such as fibrin/fibrinogen, gelatin [8], osteopontin [9], periostin [10], proteases such as ADAMTS-4 [11][12], and membrane proteins such as αM-integrin [13], Pro-TNFα [8], and LRP [14]. In addition, the type of anchoring to the cell membrane presented by MT4-MMP confers a localization in specific lipid domains that will condition its biological activity [2].
Due to their role in ECM degradation and remodeling of the pericellular microenvironment, the MMP family of proteases contributes to several physiological processes such as embryogenesis, organogenesis, tissue regeneration, angiogenesis, and wound healing. In particular, MT4-MMP expression has been reported during angiogenesis, limb development, and in distinct brain regions from early embryonic to postnatal stages [15] as well as in adult tissues such as the brain, ovaries, testis, and colon [7][16][17][18][19]. However, its physiological role remains unclear.
2. MT4-MMP Expression and Regulation in Tumors
2.1. Pattern of Expression of MT4-MMP in Normal and Tumor Tissues
Most MT-MMPs play an important role in physiological conditions. Regarding MT4-MMP, although its function has been described mainly in relation to tumor cell growth [18][20][21], its expression is spatiotemporally controlled during embryonic development. In mice, MT4-MMP is expressed in a dynamic pattern of expression from early to postnatal stages of development with a high expression of this enzyme during vascular and limb development and brain formation [15]. These data suggest that this metalloproteinase may be associated with novel functions in angiogenesis, endocardial formation, and vascularization during organogenesis as well as in central nervous system (CNS) development in correlation with its expression pattern [15]. MT4-MMP is essential for the proper organization of the aortic vessel wall. Thus, MT4-MMP expression was confirmed in vascular smooth muscle cells (VSMCs) during aortic development where, subsequently, the proteinase contributes to cell differentiation and maturation through its catalytic activity [9][15] (Figure 1A).
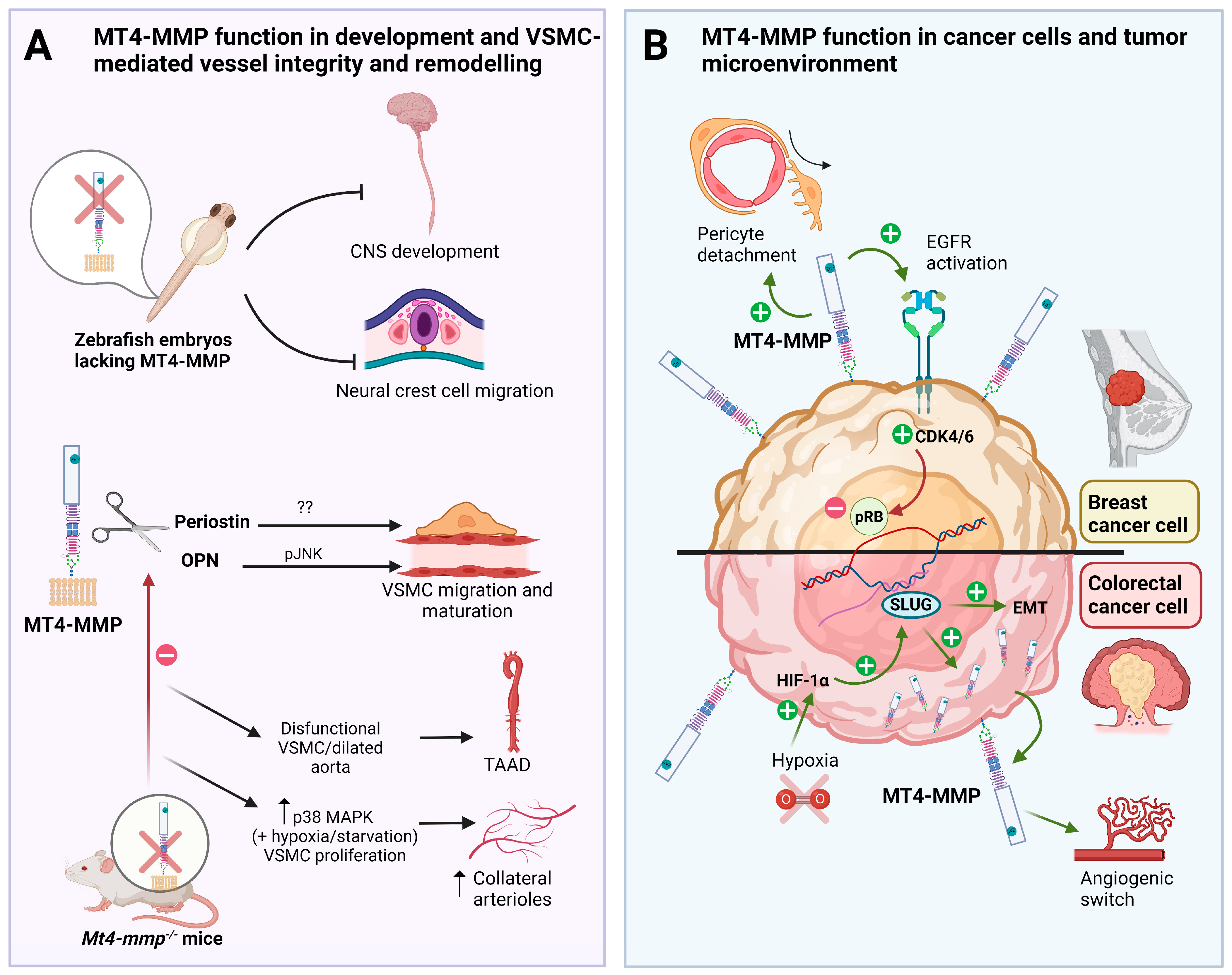
Figure 1. Role of MT4-MMP in embryonic development, VSMC-mediated vessel stability and cancer progression. (A) MT4-MMP is required for central nervous system (CNS) development and neural crest cell migration in the zebrafish embryo. MT4-MMP is expressed in VSMCs which cleaves osteopontin (OPN) to promote JNK signaling and the proper migration and maturation in the aorta. Dysfunctional VSMCs from MT4-MMP knockout mice, dilate aortas and lead to adventitial fibrosis and hypotension, playing a potential role in thoracic aortic aneurysms and dissections (TAAD). The role of periostin cleavage by MT4-MMP in VSMCs and TAAD is not known yet. Moreover, the absence of MT4-MMP promotes VSMC proliferation via p38 MAPK signaling in hypoxic/starvation context leading to an increased formation of collateral arterioles post-ligation. (B) MT4-MMP expression by breast cancer cells promotes EGFR signaling and pRB inactivation leading to tumor growth in triple-negative breast cancer (TNBC). MT4-MMP also promotes breast cancer metastasis by inducing pericyte detachment and vessel destabilization in a paracrine manner. In colorectal carcinoma, MT4-MMP expression is regulated by hypoxia and HIF1-α through SLUG and promotes cancer development since its expression is also required for early tumor angiogenic switch and tumor growth.
During CNS development, the enzyme localizes to certain regions such as the olfactory bulb, cerebral cortex, and hippocampus, suggesting a key role in brain development [15][17] (Figure 1A). Interestingly, MT4-MMP knockout mice seem to show no apparent defects in gestation, growth, fertility, and behavior and showed no evident abnormal developmental phenotypes [16]. Despite this, MT4-MMP is highly expressed in the kidney papilla and the anterior hypothalamus, and null mice display a decreased intake of water and daily urine output, suggesting a role for this enzyme in water homeostasis and the regulation of the thirst center [22].
It is also worth noticing that during embryonic development, MT4-MMP is essential for neural crest cell (NCC) migration in zebrafish [23]. In the mouse embryo, MT4-MMP is also expressed in premigratory and migratory NCCs at early stages of development [15]. This seems relevant for the regulation of cell migration, as shown in zebrafish, where it is known that its orthologue mmp17b interacts with the MMP inhibitor RECK which is required for the proper NCC migration [23].
In adult human tissues, the MT4-MMP protein was expressed preferentially in different brain regions such as the cerebral cortex, the hippocampal formation, and the basal ganglia. Additionally, this metalloproteinase is detected in female tissues (including the uterus, cervix, and ovary) where it participates in endometrial angiogenesis during the menstrual cycle [24] and in the gastrointestinal tract, particularly in the colon, under physiological conditions. In fact, among the cells that express this protein are excitatory and inhibitory neurons, oligodendrocyte precursor cells, smooth muscle cells, melanocytes, and monocytes [25][26].
Most of the current information regarding MT4-MMP comes from cancer studies. Indeed, MT4-MMP was cloned for the first time from human breast carcinoma cells [18]. Thereafter, various proteomic studies have reported the expression of this proteinase in different tumors such as prostate and oral carcinomas, lung cancer, cervical carcinomas [27][28], osteosarcomas, embryonal carcinomas, leukemia [29], adrenal adenocarcinomas, and thyroid cancer [28]. In addition, melanoma accumulates a high expression of MT4-MMP possibly related to its presence in the skin and connective tissue in physiological conditions [25][26][29]. Transcriptomic data on the expression of twenty-four MMPs, including MMP-17 from the Cancer Genome Atlas (TCGA) were reported by Gobin et al., 2019 in fifteen cancer types [30]. Differential gene expression of MT4-MMP was found to increase by more than two folds in head and neck cancer, renal clear cell cancer, lung adenocarcinoma, and in lung squamous cancer. In all breast cancers, the MT4-MMP transcript was found to increase by 1.34-fold with a significant p-value. While MT4-MMP (MMP-17) is not universally upregulated through all cancer types as happens in other MMPs (such as MMP-1, MMP-9, MMP-10, MMP-11, MMP-13, and MMP-14), the study of the prognostic value of 24 MMPs in predicting overall survival in the 15 cancer types has revealed MT4-MMP along with MMP-14, and MMP-23B as the most frequently MMPs exclusively associated with poor prognosis when they are overexpressed in a particular cancer [30].
MT4-MMP has been linked to cancer dissemination. For instance, in vitro studies and subcutaneous xenografts have confirmed the association between overexpression of MT4-MMP with cell proliferation [20][21]. In the same line, high levels of MT4-MMP expression in gastric tissues are associated with lymph node metastasis and serosal involvement, and therefore, with tumor invasion [31]. In lung metastasis, MT4-MMP alters blood vasculature and induces pericyte detachment, promoting tumor dissemination [32]. In contrast, MT4-MMP is downregulated in glioma development. In this sense, as the tumor grade advances, the expression of the proteinase continues decreasing [33]. MT4-MMP downregulation is unique to glioma because in other cancer cell lines (Jurkat or HeLa among others) MT4-MMP expression levels are higher. One possible explanation is that glioma cells are different from other cancer cells because they rarely metastasize. Moreover, brain invasion by glioma cells is very extensive locally with a large vascular development in which MT4-MMP could play an additional role. It might be interesting to analyze the specific mechanisms by which MT4-MMP downregulation favors glioma progression perhaps due to this dual role compared to other tumor cells. Furthermore, it is described that MT4-MMP is predominantly expressed by glioma cells, instead of microglia, which is the key driver for the tumor cell invasion in the CNS [34].
2.2. Regulation of MT4-MMP Expression and Activity in Normal and Tumor Tissues
Although it is known that the above-mentioned MT4-MMP functions are regulated at various levels, such as gene expression, compartmentalization, pro-enzyme cleavage, and substrate processing [35][36], little is known about the detailed molecular mechanisms relevant to tissue growth and expansion. However, its proteolytic activity has been shown to be significantly linked to its pro-metastatic activity, and the latter may explain certain aspects of its role in cancer progression (Figure 1B). For example, in contrast to most of MT-MMPs, MT4-MMP hydrolyses very few ECM components [1], and it also exhibits characteristic sensitivity to TIMPS as well as inefficient activation of pro-MMP2, which may explain its singular role in promoting tumor progression.
2.2.1. Transcription
Regarding gene expression, classical methods have been enriched by using a novel technology known as super-resolution microscopy (SRM) that allows the study of gene regulation in a much more detailed way [37]. The use of SRM approaches could be interesting for the study of MT4-MMP transcription mechanisms that are not yet fully understood.
It has been reported that MT4-MMP expression is induced by hypoxia through the hypoxia-inducible factor-1-α (HIF1-α) and the activation of SLUG, a known transcription factor involved in epithelial–mesenchymal transition (EMT) which promotes the malignant capacity of cancer cells (Figure 1B) [38]. In fact, invadopodia formation and amoeboid movements, which are both crucial mechanisms to promote metastatic dissemination, are mediated by HIF1-α-induced MT4-MMP expression in head and neck cancer tumor cells [39]. An increase in MT4-MMP has been also observed under hypoxic conditions or under constitutive expression of HIF1-α in other types of tumors such as hypopharyngeal squamous cell carcinoma (FADU) and tongue squamous cell carcinoma (SAS) [38]. Interestingly, SLUG has been identified as the key factor responsible for the hypoxia-induced MT4-MMP expression through the activation of the proteinase promoter by interacting with its E-box [38]. Notably, SLUG expression was mostly restricted to migrating neural crest cells and several mesodermal derivatives in the embryo [40], suggesting that this transcription factor may regulate MT4-MMP expression both during development and tumorigenesis. Therefore, co-expression of MT4-MMP and HIF-1α may be considered as an indicator of breast cancer prognosis. It is also worth noting that in human breast cancer, MT4-MMP transcription is also regulated by the methyltransferase hSED1A, which appears to be over-expressed in these circumstances. It is known that silencing hSED1A decreases MT4-MMP transcription, which impairs cell migration and invasion of tumor cells on lung tissue and colon cancer cells [41].
2.2.2. Post-Translational Regulation
-
Compartmentation
As a GPI-anchored protein, MT4-MMP is located on lipid domains that seem to be relevant for its activity [2]. In line with this, the HM-7 cancer cell line expresses MT4-MMP in lipid rafts, while caveolin-1 is not detected. Interestingly, restoration of caveolin-1 expression in metastatic HM-7 cells inhibits MT4-MMP localization to lipid rafts, thereby suppressing the metastatic phenotype of HM-7 colon cancer cells. These findings raise the possibility that MT4-MMP compartmentation may be directly or indirectly involved in certain intracellular signaling events that control its pericellular proteolytic capacity [42].
The functional cooperation between MT4-MMP and other GPI proteins such as the urokinase receptor uPAR may be possible since both colocalize in the same microdomains [7]. Although this role during embryonic development remains elusive, this putative co-expression could be relevant for tumor invasion in several cancers [31]. Indeed, MT4-MMP requires a permissive microenvironment to exert its tumor-promoting effect. Tumor-derived MT4-MMP cannot circumvent the absence of a host angio-promoting factor such as the plasminogen activator inhibitor-1 (PAI-1), which cooperates with the uPA/uPAR axis in different contexts [43].
-
Internalization and recycling
Endocytosis is the mechanism that controls the amount of MT4-MMP anchored to the cell surface through the CLIC/GEEC pathway as well as recycles it back to the cell membrane [5][44]. The signaling cascade that triggers this endocytic pathway involves the Rho family GTPase and Cdc42 and the transcription factors Arf1 or GBF1 that are responsible for the regulation of Cdc42 activity. For instance, MT4-MMP internalization was shown to be primarily dependent on Cdc42 and RhoA, and to a lesser extent on Rac1 in MDA-MB-231 cells, a human breast cancer cell line overexpressing the proteinase. These data suggest that MT4-MMP proficiently uses the CLIC/GEEC pathway (rather than the caveolin-dependent or clathrin-dependent pathways) for its internalization [5]. It should be mentioned that the actin-binding protein Swiprosin-1 (Swip1) functions as a cargo-specific adaptor for CLIC/GEEC endocytic pathway mediating the endocytosis of active integrins, supporting integrin-dependent cancer cell migration and invasion [45]. As MT4-MMP can also regulate the levels and activity of integrins, particularly β2-integrins, in other cellular contexts [13], this endocytic pathway may be particularly relevant for regulating MT4-MMP activity in different cell types and physio-pathological contexts.
-
Dimerization
A relationship between MT-MMP dimerization and greater proteolytic activity in tumor cells is feasible [46][47]. MT4-MMP is found in homodimers and oligomers at the cell surface maintained via disulfide bond between the cysteine residues of the stem region [4][48]. In the context of tumorigenesis, MT4-MMP can form homodimers depending on Cys574 and the formation of disulfide bonds between the monomers both in transfected non-tumor MDCK and COS1 cells and in MD-MB-231 breast carcinoma cells [5][48]. Although the impairment of dimerization did not decrease cell invasion in vitro, it could still be relevant for other activities in the tumor context.
-
Shedding
Shedding is another alternative mechanism used by MT-MMPs to control their pericellular proteolytic activity once they are anchored to the cell membrane [3][49]. This mechanism may either involve the release of the extracellular portion of the active MT-MMP to the cellular milieu or the removal of the enzyme from the cell surface. However, the precise mechanism of MT4-MMP shedding remains to be elucidated and it is not fully understood how MT4-MMP regulates the balance between the amount of protein anchored to the membrane by its GPI moiety and the soluble enzyme since it is not affected in the presence of TIMPs [4][7]. The possibility that this metalloproteinase could be released from the cell surface through the activity of a phosphatidylinositol (PI) specific phospholipase C, similarly to other membrane dipeptidases, cannot be ruled out [7].
-
Interactions
EGFR has been reported to associate with MT4-MMP by co-immunoprecipitation. Both EGFR and MT4-MMP have been shown to cooperate in tumor cell invasion and signaling, driving cancer cell growth through the regulation of cell cycle proteins such as CDK4 activation and retinoblastoma protein inactivation (Figure 1B) [21]. MT4-MMP is thought to stimulate cell proliferation by interacting with EGFR and enhancing its activation by its ligands, the epidermal growth factor (EGF), and tumor growth factor (TGF) in cancer cells (Figure 1B). Whether this functional cooperation is also relevant during embryonic development, for example in the heart formation [50], remains to be investigated.
This entry is adapted from the peer-reviewed paper 10.3390/ijms24129944
References
- Moracho, N.; Learte, A.I.R.; Muñoz-Sáez, E.; Marchena, M.A.; Cid, M.A.; Arroyo, A.G.; Sánchez-Camacho, C. Emerging roles of MT-MMPs in embryonic development. Dev. Dyn. 2022, 251, 240–275.
- Yip, C.; Foidart, P.; Noël, A.; Sounni, N.E.; Noel, A.; Sounni, N.E. MT4-MMP: The GPI-anchored membrane-type matrix metalloprotease with multiple functions in diseases. Int. J. Mol. Sci. 2019, 20, 354.
- Itoh, Y. Membrane-type matrix metalloproteinases: Their functions and regulations. Matrix Biol. 2015, 44–46, 207–223.
- Sohail, A.; Sun, Q.; Zhao, H.; Bernardo, M.M.; Cho, J.-A.; Fridman, R. MT4-(MMP17) and MT6-MMP (MMP25), a unique set of membrane-anchored matrix metalloproteinases: Properties and expression in cancer. Cancer Metastasis Rev. 2008, 27, 289–302.
- Truong, A.; Yip, C.; Paye, A.; Blacher, S.; Munaut, C.; Deroanne, C.; Noel, A.; Sounni, N.E. Dynamics of internalization and recycling of the prometastatic membrane type 4 matrix metalloproteinase (MT4-MMP) in breast cancer cells. FEBS J. 2016, 283, 704–722.
- Wang, Y.; Johnson, A.R.; Ye, Q.Z.; Dyer, R.D. Catalytic activities and substrate specificity of the human membrane type 4 matrix metalloproteinase catalytic domain. J. Biol. Chem. 1999, 274, 33043–33049.
- Itoh, Y.; Kajita, M.; Kinoh, H.; Mori, H.; Okada, A.; Seiki, M. Membrane type 4 matrix metalloproteinase (MT4-MMP, MMP-17) is a glycosylphosphatidylinositol-anchored proteinase. J. Biol. Chem. 1999, 274, 34260–34266.
- English, W.R.; Puente, X.S.; Freije, J.M.; Knauper, V.; Amour, A.; Merryweather, A.; Lopez-Otin, C.; Murphy, G. Membrane type 4 matrix metalloproteinase (MMP17) has tumor necrosis factor-alpha convertase activity but does not activate pro-MMP2. J. Biol. Chem. 2000, 275, 14046–14055.
- Martin-Alonso, M.; Garcia-Redondo, A.B.; Guo, D.; Camafeita, E.; Martinez, F.; Alfranca, A.A.; Mendez-Barbero, N.; Pollan, A.; Sanchez-Camacho, C.; Denhardt, D.T.; et al. Deficiency of MMP17/MT4-MMP proteolytic activity predisposes to aortic aneurysm in mice. Circ. Res. 2015, 117, e13–e26.
- Martín-Alonso, M.; Iqbal, S.; Vornewald, P.M.; Lindholm, H.T.; Damen, M.J.; Martínez, F.; Hoel, S.; Díez-Sánchez, A.; Altelaar, M.; Katajisto, P.; et al. Smooth muscle-specific MMP17 (MT4-MMP) regulates the intestinal stem cell niche and regeneration after damage. Nat. Commun. 2021, 12, 6741.
- Gao, G.; Westling, J.; Thompson, V.P.; Howell, T.D.; Gottschall, P.E.; Sandy, J.D. Activation of the proteolytic activity of ADAMTS4 (aggrecanase-1) by C-terminal truncation. J. Biol. Chem. 2002, 277, 11034–11041.
- Gao, G.; Plaas, A.; Thompson, V.P.; Jin, S.; Zuo, F.; Sandy, J.D. ADAMTS4 (aggrecanase-1) activation on the cell surface involves C-terminal cleavage by glycosylphosphatidyl inositol-anchored membrane type 4-matrix metalloproteinase and binding of the activated proteinase to chondroitin sulfate and heparan sulfate on syndecan-1. J. Biol. Chem. 2004, 279, 10042–10051.
- Clemente, C.; Rius, C.; Alonso-Herranz, L.; Martin-Alonso, M.; Pollan, A.; Camafeita, E.; Martinez, F.; Mota, R.A.; Nunez, V.; Rodriguez, C.; et al. MT4-MMP deficiency increases patrolling monocyte recruitment to early lesions and accelerates atherosclerosis. Nat. Commun. 2018, 9, 910.
- Rozanov, D.V.; Hahn-Dantona, E.; Strickland, D.K.; Strongin, A.Y. The low density lipoprotein receptor-related protein LRP is regulated by membrane type-1 matrix metalloproteinase (MT1-MMP) proteolysis in malignant cells. J. Biol. Chem. 2004, 279, 4260–4268.
- Blanco, M.J.; Rodriguez-Martin, I.; Learte, A.I.R.; Clemente, C.; Montalvo, M.G.; Seiki, M.; Arroyo, A.G.; Sanchez-Camacho, C. Developmental expression of membrane type 4-matrix metalloproteinase (Mt4-mmp/Mmp17) in the mouse embryo. PLoS ONE 2017, 12, e0184767.
- Rikimaru, A.; Komori, K.; Sakamoto, T.; Ichise, H.; Yoshida, N.; Yana, I.; Seiki, M. Establishment of an MT4-MMP-deficient mouse strain representing an efficient tracking system for MT4-MMP/MMP-17 expression in vivo using beta-galactosidase. Genes Cells 2007, 12, 1091–1100.
- Funatsu, N.; Inoue, T.; Nakamura, S. Gene expression analysis of the late embryonic mouse cerebral cortex using DNA microarray: Identification of several region- and layer-specific genes. Cereb. Cortex 2004, 14, 1031–1044.
- Puente, X.S.; Pendás, A.M.; Llano, E.; Velasco, G.; López-Otín, C. Molecular cloning of a novel membrane-type matrix metalloproteinase from a human breast carcinoma. Cancer Res. 1996, 56, 944–949.
- Mittal, R.; Patel, A.P.; Debs, L.H.; Nguyen, D.; Patel, K.; Grati, M.; Mittal, J.; Yan, D.; Chapagain, P.; Liu, X.Z. Intricate Functions of Matrix Metalloproteinases in Physiological and Pathological Conditions. J. Cell Physiol. 2016, 231, 2599–2621.
- Chabottaux, V.; Sounni, N.E.; Pennington, C.J.; English, W.R.; van den Brûle, F.; Blacher, S.; Gilles, C.; Munaut, C.; Maquoi, E.; Lopez-Otin, C.; et al. Membrane-type 4 matrix metalloproteinase promotes breast cancer growth and metastases. Cancer Res. 2006, 66, 5165–5172.
- Paye, A.; Truong, A.; Yip, C.; Cimino, J.; Blacher, S.; Munaut, C.; Cataldo, D.; Foidart, J.M.; Maquoi, E.; Collignon, J.; et al. EGFR activation and signaling in cancer cells are enhanced by the membrane-bound metalloprotease MT4-MMP. Cancer Res. 2014, 74, 6758–6770.
- Srichai, M.B.; Colleta, H.; Gewin, L.; Matrisian, L.; Abel, T.W.; Koshikawa, N.; Seiki, M.; Pozzi, A.; Harris, R.C.; Zent, R. Membrane-type 4 matrix metalloproteinase (MT4-MMP) modulates water homeostasis in mice. PLoS ONE 2011, 6, e17099.
- Leigh, N.R.; Schupp, M.-O.; Li, K.; Padmanabhan, V.; Gastonguay, A.; Wang, L.; Chun, C.Z.; Wilkinson, G.A.; Ramchandran, R. Mmp17b is essential for proper neural crest cell migration in vivo. PLoS ONE 2013, 8, e76484.
- Plaisier, M.; Koolwijk, P.; Hanemaaijer, R.; Verwey, R.A.; van der Weiden, R.M.F.; Risse, E.K.J.; Jungerius, C.; Helmerhorst, F.M.; van Hinsbergh, V.W.M. Membrane-type matrix metalloproteinases and vascularization in human endometrium during the menstrual cycle. Mol. Hum. Reprod. 2006, 12, 11–18.
- Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000198598-MMP17 (accessed on 22 January 2023).
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419.
- Riddick, A.C.P.; Shukla, C.J.; Pennington, C.J.; Bass, R.; Nuttall, R.K.; Hogan, A.; Sethia, K.K.; Ellis, V.; Collins, A.T.; Maitland, N.J.; et al. Identification of degradome components associated with prostate cancer progression by expression analysis of human prostatic tissues. Br. J. Cancer 2005, 92, 2171–2180.
- Grant, G.M.; Giambernardi, T.A.; Grant, A.M.; Klebe, R.J. Overview of expression of matrix metalloproteinases (MMP-17, MMP-18, and MMP-20) in cultured human cells. Matrix Biol. 1999, 18, 145–148.
- Hieronimus, B.; Pfohl, J.; Busch, C.; Graeve, L. Expression and characterization of membrane-type 4 matrix metalloproteinase (MT4-MMP) and its different forms in melanoma. Cell Physiol. Biochem. 2017, 42, 198–210.
- Gobin, E.; Bagwell, K.; Wagner, J.; Mysona, D.; Sandirasegarane, S.; Smith, N.; Bai, S.; Sharma, A.; Schleifer, R.; She, J.-X. A pan-cancer perspective of matrix metalloproteases (MMP) gene expression profile and their diagnostic/prognostic potential. BMC Cancer 2019, 19, 581.
- Wang, Y.; Yu, S.-J.; Li, Y.-X.; Luo, H.-S. Expression and clinical significance of matrix metalloproteinase-17 and -25 in gastric cancer. Oncol. Lett. 2015, 9, 671–676.
- Chabottaux, V.; Ricaud, S.; Host, L.; Blacher, S.; Paye, A.; Thiry, M.; Garofalakis, A.; Pestourie, C.; Gombert, K.; Bruyere, F.; et al. Membrane-type 4 matrix metalloproteinase (MT4-MMP) induces lung metastasis by alteration of primary breast tumour vascular architecture. J. Cell Mol. Med. 2009, 13, 4002–4013.
- Nuttall, R.K.; Pennington, C.J.; Taplin, J.; Wheal, A.; Yong, V.W.; Forsyth, P.A.; Edwards, D.R. Elevated membrane-type matrix metalloproteinases in gliomas revealed by profiling proteases and inhibitors in human cancer cells. Mol. Cancer Res. 2003, 1, 333–345.
- Thome, I.; Lacle, R.; Voß, A.; Bortolussi, G.; Pantazis, G.; Schmidt, A.; Conrad, C.; Jacob, R.; Timmesfeld, N.; Bartsch, J.W.; et al. Neoplastic cells are the major source of MT-MMPs in IDH1-mutant glioma, thus enhancing tumor-cell intrinsic brain infiltration. Cancers 2020, 12, 2456.
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629.
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67.
- Hoboth, P.; Šebesta, O.; Hozák, P. How Single-Molecule Localization Microscopy Expanded Our Mechanistic Understanding of RNA Polymerase II Transcription. Int. J. Mol. Sci. 2021, 22, 6694.
- Huang, C.-H.; Yang, W.-H.; Chang, S.-Y.; Tai, S.-K.; Tzeng, C.-H.; Kao, J.-Y.; Wu, K.-J.; Yang, M.-H. Regulation of membrane-type 4 matrix metalloproteinase by SLUG contributes to hypoxia-mediated metastasis. Neoplasia 2009, 11, 1371–1382.
- Yan, X.; Cao, N.; Chen, Y.; Lan, H.Y.; Cha, J.H.; Yang, W.H.; Yang, M.H. MT4-MMP promotes invadopodia formation and cell motility in FaDu head and neck cancer cells. Biochem. Biophys. Res. Commun. 2020, 522, 1009–1014.
- Savagner, P.; Karavanova, I.; Perantoni, A.; Thiery, J.P.; Yamada, K.M. Slug mRNA is expressed by specific mesodermal derivatives during rodent organogenesis. Dev. Dyn. 1998, 213, 182–187.
- Salz, T.; Li, G.; Kaye, F.; Zhou, L.; Qiu, Y.; Huang, S. hSETD1A regulates Wnt target genes and controls tumor growth of colorectal cancer cells. Cancer Res. 2014, 74, 775–786.
- Nimri, L.; Barak, H.; Graeve, L.; Schwartz, B. Restoration of caveolin-1 expression suppresses growth, membrane-type-4 metalloproteinase expression and metastasis-associated activities in colon cancer cells. Mol. Carcinog. 2013, 52, 859–870.
- Host, L.; Paye, A.; Detry, B.; Blacher, S.; Munaut, C.; Foidart, J.M.; Seiki, M.; Sounni, N.E.; Noel, A. The proteolytic activity of MT4-MMP is required for its pro-angiogenic and pro-metastatic promoting effects. Int. J. Cancer 2012, 131, 1537–1548.
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612.
- Moreno-Layseca, P.; Jäntti, N.Z.; Godbole, R.; Sommer, C.; Jacquemet, G.; Al-Akhrass, H.; Conway, J.R.W.; Kronqvist, P.; Kallionpää, R.E.; Oliveira-Ferrer, L.; et al. Cargo-specific recruitment in clathrin- and dynamin-independent endocytosis. Nat. Cell Biol. 2021, 23, 1073–1084.
- Genís, L.; Gálvez, B.G.; Gonzalo, P.; Arroyo, A.G. MT1-MMP: Universal or particular player in angiogenesis? Cancer Metastasis Rev. 2006, 25, 77–86.
- Suárez, H.; López-Martín, S.; Toribio, V.; Zamai, M.; Hernández-Riquer, M.V.; Genís, L.; Arroyo, A.G.; Yáñez-Mó, M. Regulation of MT1-MMP activity through its association with ERMs. Cells 2020, 9, 348.
- Sohail, A.; Marco, M.; Zhao, H.; Shi, Q.; Merriman, S.; Mobashery, S.; Fridman, R. Characterization of the dimerization interface of membrane type 4 (MT4)-matrix metalloproteinase. J. Biol. Chem. 2011, 286, 33178–33189.
- Osenkowski, P.; Toth, M.; Fridman, R. Processing, shedding, and endocytosis of membrane type 1-matrix metalloproteinase (MT1-MMP). J. Cell Physiol. 2004, 200, 2–10.
- Iwamoto, R.; Mekada, E. ErbB and HB-EGF signaling in heart development and function. Cell Struct. Funct. 2006, 31, 1–14.
This entry is offline, you can click here to edit this entry!