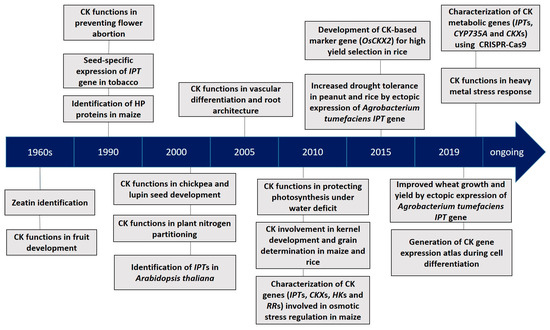
Cytokinins (CKs) are a class of the multifaceted plant hormone that plays essential roles in a wide array of processes of plant growth and development [
13]. Since the discovery of their existence in maize (
Zea mays) seeds over 50 years ago, CKs have been extensively studied for their chemical nature, metabolism and signal transduction pathway, as well as their functions in plant growth and development (). Significant regulatory functions of CKs at the tissue and organ levels include the inhibition of lateral root initiation [
14], differentiation of phloem and metaxylem in roots [
15], regulation of cell division, photomorphogenic cell differentiation in expanding leaves and shoots [
16,
17], and inhibition of leaf senescence [
18].
Endogenous CKs are adenine derivatives with isoprenoid or aromatic side chains. Depending on hydroxylation or reduction of the side chain, isoprenoid CKs, which are widespread in nature, can be distinguished as isopentenyladenine (iP)-,
trans-zeatin (
tZ)-,
cis-zeatin (
cZ)- or dihydrozeatin (DHZ)-type derivatives. By contrast, aromatic CKs, such as N6-(meta-hydroxybenzyl)adenine (BA), are found in plants at a lower abundance [
19]. The isoprenoid CKs differ in their biological functions, biochemical properties, metabolic conversions, and their transportability across the plant body [
13]. Various enzymes involved in CK metabolism, including CK biosynthesis, inter-conversion between CK types and CK degradation, are responsible for maintaining CK homeostasis [
20]. CK metabolism has been extensively explored to understand the function of a large number of genes and enzymes, as well as the composed metabolic network regulated by CKs throughout the plant kingdom (reviewed by Pavlů et al. [
13]; Zwack et al. [
18]). CK signaling pathway is also triggered by various environmental stimuli, such as changes in temperature, nutrition levels and osmotic conditions, whereby the phospho-relay cascades of the two-component system (TCS) (i.e., downstream components of CK signaling, which will be discussed in detail in
Section 2.2), are initiated and lead to the expression regulation of specific genes involved in plant adaptation [
13,
20]. More recently, CK crosstalk with ethylene (ET), jasmonates (JAs), salicylic acid (SA) and abscisic acid (ABA) have been recognized [
17,
20,
21,
22], indicating a coordinating network among these phytohormones in plant stress tolerance. Furthermore, CK biosynthesis and signaling components are known to act as constitutive signals defining plant response to drought stress and regulating plant drought acclimation [
13]. Their rapid responses, spatiotemporal expression, as well as their widely associated pathways, make CKs excellent candidates to regulate complex morphogenetic processes under water deficit. Therefore, in this review, we will emphasize and propose a model for the role and regulation of plant CKs under drought stress.