Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Immunology
Immunotherapies including adaptive immune checkpoint inhibitors (ICIs) and chimeric antigen receptor (CAR) T cells, have developed the treatment of cancer in clinic, and most of them focus on activating T cell immunity. Although these strategies have obtained unprecedented clinical responses, only limited subsets of cancer patients could receive long-term benefits, highlighting the demand for identifying novel targets for the new era of tumor immunotherapy. Innate immunity has been demonstrated to play a determinative role in the tumor microenvironment (TME) and influence the clinical outcomes of tumor patients.
- innate immunity
- cancer therapy
- immune checkpoint inhibitors
1. Introduction
Cancer is one of the main leading causes of disease-associated death and it often results from genotoxic and non-genotoxic carcinogens [1]. Surgery, chemotherapy, radiotherapy and tumor immunotherapy are the principle treatment for tumors. Surgery is the treatment of choice for most solid tumors, but cannot be used for hematologic tumors and metastases and subclinical metastases of solid tumors. Compared to surgical treatment, chemotherapy is a systemic treatment for primary, metastatic and subclinical metastases of solid tumors as well as hematologic tumors. However, the poor selectivity of chemotherapeutic agents limits its clinical application. Radiation therapy protects tissue that is not affected by the tumor, produces less damage and does not require hospitalization, but the application of radiotherapy depends on the type, size and location of tumors. Compared with these traditional therapies, tumor immunotherapy can kill tumor cells by activating the host’s immune system [2,3]. The discipline of oncology has been undergoing an unprecedented revolution due to the massive advancement of immunotherapy in the treatment of cancer [4]. Infiltrating into the TME, immune cells assist in tuning the development of tumors [5,6]. In adaptive immunity, T cells participate in immune responses that are cell-mediated immune responses [7,8,9]. The treatment of solid tumors has been revolutionized by ICIs targeting PD-1/PD-L1. Blocking the PD-1/PD-L1 axis could enhance T cell infiltration and revitalize exhausted cytotoxic T cells, presenting impressive therapeutic efficacy in various cancers [10]. Interestingly, PD-L1 expression and sensitivity to immunotherapy of tumor cells are subject to metabolic reprogramming. For example, mitochondrial oxidative phosphorylation (OXPHOS) can regulate PD-L1 levels. Tumor-selective OXPHOS suppression nanoparticles have been reported to reactivate immunotherapy and may open a new therapeutic window for patients [11]. Furthermore, CAR T cells have shown impressive efficacy in the treatment of hematologic malignancies [12,13,14]. As the mechanisms of tumor development and treatment are further explored, many new strategies have also emerged. For instance, oxidative stress and reactive oxygen species (ROS) contribute to the development of cancer growth [15]. Myrrh, a traditional remedy, could be a possible therapeutic agent for cancer because of the powerful antioxidant activity of its extracts [15]. Pyroptosis is a form of programmed cell death, which is associated with tumor genesis, and therapy response. Inflammasomes are multimolecular complexes containing pattern recognition receptors that can recruit the adaptor protein containing apoptosis-associated speck-like protein containing a caspase recruitment domain and activate caspase-1. To date, six members of the gasdermin (GSDM) family have been identified, namely GSDMA, GSDMB, GSDMC, GSDMD, GSDME (DFNA5) and DFNB59. Inflammasomes can activate caspase-1 that can cleave GSDMD to induce pyroptosis [16]. It is reported that the virus-like particle can trigger the formation of an AIM2 inflammasome that can induce GSDMD-mediated pyroptosis, thus enhancing antitumor immunity [17]. The primary goal of the currently approved immunotherapies is to active T cell immunity, but innate immunity also has significant anti-tumor potential. The desire to identify novel targets in innate immunity has emerged in the new era of immunotherapy.
In the innate immune system, myeloid-derived suppressor cells (MDSCs), macrophages, neutrophils, natural killer (NK) cells, dendritic cells (DCs), mast cells (MCs) and helper innate lymphoid cells (ILCs) have important regulatory effects on tumor progression [18]. Some innate immune cells have the capacity for detecting and eliminating tumor cells through various mechanisms, such as intrinsic cytotoxicity of NK cells and macrophage phagocytosis. The interaction of the innate and adaptive immunity is exemplified by the uptake of tumor antigens by antigen-presenting cells (APCs), which results in cross-presentation and the priming of CD8+ T lymphocytes [19]. Innate immune cells are also involved in effector responses through antibody-dependent cellular phagocytosis (ADCP) and antibody-dependent cell cytotoxicity (ADCC) [20].
2. Innate Immunity in TME
Composed of MDSCs, macrophages, neutrophils, NK cells, DCs, MCs and helper ILCs, the innate immunity plays a crucial role in defense against tumors (Figure 1). For example, macrophages can activate the tumor-killing activity of T cells through antigen cross-presentation. Antigen cross-presentation by macrophages can be achieved through the cytosolic and vacuolar pathways. In the cytosolic pathway, proteins are transported to the cell membrane, where they are degraded by the proteasome. Subsequently, the derived peptides are transferred via the transporter associated with antigen presentation to the endoplasmic reticulum (ER), where they are processed by aminopeptidases of the ER, or brought back to the antigen-containing endosome to be processed by insulin-regulated aminopeptidases. In this process, the peptide is loaded on the MHC-I. The other major cross-presentation pathway is the vacuolar pathway, in which proteins are processed by endosomal/lysosomal proteases and loaded on MHC-I within the endosomal/lysosomal compartments. The resulting peptide MHC-I molecular complex is recognized by CD8+ T cells expressing specific T cell receptors (TCRs). The signal from TCR recognition of antigen is transduced into CD8+ T cells, which triggers cytotoxic effects [21]. Neutrophils can promote tumor proliferation and invasion by secreting proteases, and can also inhibit tumor progression by secreting H2O2, based on the type, stage and location of the tumors [22]. NK cells achieve anti-tumor effects by mediating ADCC, and MCs can promote tumors or fight tumors by releasing cytokines, chemokines, eicosanoids, proteoglycans and biogenic amines [23,24]. A comprehensive understanding of innate immune cell function in the TME is vital for the design of effective cancer immunotherapy.
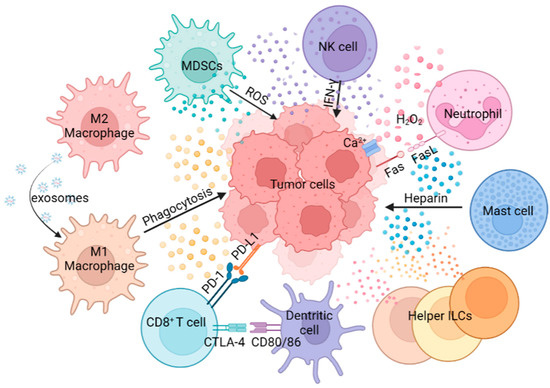
Figure 1. The role of innate immune cells in tumor progression. Innate immune cells show multiple roles in tumor progression through antigen presentation, phagocytosis, secretion of cytokines, direct killing effect, etc.
2.1. MDSCs
As a heterogeneous population of immature myeloid cells that possess potent immunosuppressive properties, MDSCs are typically classified into two major subsets based on their phenotypic, morphological and functional properties: mononuclear phagocyte-like MDSCs (M-MDSCs) and granulocyte-like MDSCs (G-MDSCs) [25,26,27]. G-MDSCs are predominantly abundant within the TME, whereas M-MDSCs primarily accumulate in the peripheral blood [28]. MDSCs play a crucial role in modulating immune responses and their dysregulation is associated with numerous diseases, including cancer, chronic infections, autoimmune disorders and inflammatory conditions [27]. In cancer, MDSCs suppress antitumor immune responses, promote tumor progression and contribute to immune evasion through a number of mechanisms, including boosting the production of Arg1, iNOS, ROS and nitrogen compounds like peroxynitrite (PNT) [29]. When Arg-1 is increased, L-arginine, an essential amino acid for T cell proliferation, could be metabolized into urea and L-ornithine [30]. When L-arginine is restricted, the up-regulation of iNOS contributes to NO production, which reacts with superoxide to produce PNT [31]. Subsequently, PNT causes the TCR nitration and nitrosylation, inhibiting the function of CD8+ T cells and promoting T cell tolerance [32,33]. Therefore, one crucial way of targeting MDSCs to reverse the immunosuppressive TME is to understand the basic mechanisms of MDSC recruitment into the TME [28].
2.2. Macrophages
Macrophages, which originate from myeloid-derived progenitor cells in the bone marrow, are the main phagocytes in tumors [34]. Macrophages can eliminate cellular debris clear pathogens and regulate inflammatory responses, which are essential for homeostasis maintenance [35]. Macrophages can be polarized into M1 phenotype and M2 phenotype [36]. M1 macrophages are characterized by producing pro-inflammatory cytokines such as interleukin (IL)-6, IL-12, and interferon (IFN)-γ, which possess anti-tumor properties. In addition, M1 macrophages recruit T helper type 1 (Th1) cells by secreting the chemokines CXCL9 and CXCL10, and promote T cell responses by upregulating the gene of antigen presentation and costimulatory molecules [35]. In contrast, M2 macrophages exhibit pro-tumor activities and produce immunosuppressive cytokines such as IL-4, IL-10, and transforming growth factor-β (TGF-β). In the TME, tumor-infiltrating macrophages, also known as tumor-associated macrophages (TAM), mainly exhibit an M2-like phenotype [37,38].
2.3. Neutrophils
Neutrophils, derived from the early committed neutrophil progenitor cells, are considered to be the most massive innate immune cells in the bone marrow and peripheral blood. These cells can respond quickly to inflammation, infection and injury. They can promote cancer progression through various mechanisms, such as angiogenesis, immunosuppression and cancer metastasis. In promoting tumor invasion, neutrophils secrete proteases that degrade structural proteins in the extracellular environment [39,40,41]. For instance, studies on breast cancer have shown that neutrophils can induce MMP-12 and MMP-13 to promote cancer metastasis [42]. Independent clinical studies have also confirmed the important role of neutrophils in tumor metastasis [43,44,45,46,47]. In addition, IL-1β promotes IL-17 production from γδT cells, leading to polarization of neutrophils in mice with mammary tumors [48]. Nonetheless, neutrophils can also exert anti-tumor by, for example, secreting H2O2 to cause an influx of Ca2+ and inducing apoptosis in tumor cells via Fas ligand/Fas interaction [22].
2.4. NK Cells
NK cells are cytotoxic lymphocytes that hold the capacity to kill cancerous cells. In general, NK cells can kill tumor cells via direct cytotoxicity and pro-inflammatory cytokines production [24]. “Missing-self” is one mechanism of direct cytotoxicity. Immunoreceptor tyrosine-based inhibitory motifs (ITIMs) can engage with the major histocompatibility complex-I (MHC-I) to stop NK cells from destroying ordinary cells, which means NK cell activation can be inhibited by the binding of MHC-I. However, tumor cells always lack or only very faintly express MHC-I in order to escape CD8+ T cell-mediated cytotoxicity. Thus, NK cells can recognize and reply to this missing-self phenotype [49]. ADCC is another crucial mechanism of NK cell-mediated direct cytotoxicity [24]. Additionally, NK cells express significant quantities of IFN-γ and TNF. They can not only enhance cytotoxic CD8+ T cell response, but also can they suppress the proliferation and angiogenesis of tumor cells while they promote the apoptosis of cancerous cells [50].
2.5. DCs
First discovered in 1973 by Ralph Steinman, DC is one of the most important APCs, which plays a significant role in launching and modulating innate and adaptive immune responses [51]. DCs comprise four main subsets: cDC1s, cDC2s, plasmacytoid DCs (pDCs) and monocyte-derived DCs (MoDCs). In the TME, DCs recognize, uptake, process and present tumor-associated antigens (TAAs) to T cells to shape T cell responses [52]. Distinct DCs have different T cell priming abilities. cDC1s excel at cross-presentation antigens, which could activate CD8+ T cells and promote Th1 cell polarization of CD4+ T cells. cDC2s are essential to inducing CD4+ T cell responses [53]. pDCs could effectively prime CD8+ T cells while showing poor priming of naive T cells [54,55,56]. MoDCs are mainly generated in inflammatory responses and are critical to the differentiation of CD4+ T cells. When perceiving appropriate cues, DCs express costimulatory molecules such as CD80, CD86, CD137L, OX40L, GITRL, CD70 and CD40 to regulate DC-mediated T cell priming to influence T cell-mediated immunotherapy in TME [51]. DCs also upregulated chemokine receptors such as cc-chemokine receptor 7 (CCR7) [57]. Studies show that CCR7 is critical for the migration of tumor-infiltrating DCs and can impact DCs recruitment into the TME. Chemokines such as CXC-chemokine ligand 9 (CXCL9) and CXCL10 are also produced by DCs to facilitate the recruitment of CD8+ T cells in the TME [58]. In addition, DCs could produce cytokines such as IL-12 and type 1 TNF; IL-12s are pivotal to the initiation of Th1 cells and CD8+ T cells, and type 1 TNFs have been used to treat patients with cancer [19,59,60,61].
2.6. MCs
MCs are derived from bone marrow precursors, which exert either pro- or anti-tumor effects by secreting cytokines, chemokines, eicosanoids, proteoglycans and biogenic amines after activation [23]. Studies demonstrate that IL-33-mediated MC activation promotes TAM infiltration and tumorigenesis by secreting CSF2, CCL3 and IL-6 in gastric cancer [62]. Eicosanoids, such as LTC4, LTB4 and PGD2, are also secreted upon MC activation. Transforming LTC4 to LTD4 by γ-Glutamyl transpeptidase 1 (GGT1) and GGT5, enzymes that hydrolyze γ-glutamyl of glutathione, results in lung cancer cell migration and survival via CysLT1 [63,64]. LTB4 has the potential to be a biomarker for predicting the effect of bestatin in colorectal cancer (CRC) [65]. PGD2 can suppress tumorigenesis and metastasis via PGD2/PTGDR2 signaling [66,67]. Mast cell-derived heparin is a kind of proteoglycan that could increase angiogenesis. In mouse EL-4 lymphoma and MC-38 colon cancer, histamine produced by MCs could target MDSCs and enhance the anti-tumor efficacy of checkpoint blocking [68].
2.7. Helper ILCs
ILCs are a subset of lymphocytes that lack antigen-specific receptors. There are five subsets of ILCs: NK cells, lymphoid tissue inducer cells, and groups 1, 2, and 3 (ILC1, ILC2, and ILC3) [69,70]. The Helper ILC family, which consists of ILC1, ILC2, and ILC3, can be utilized to induce and stimulate anti-tumor responses [71]. ILC1s have the capacity to express IFN-γ, NKp46, and T-bet [72]. Type 2 cytokines, including IL-5 and IL-13, are produced by ILC2s in response to TSLP, IL-18, IL-33, or IL-25 [73]. Tumor-infiltrating helper ILCs can either stimulate or restrict the proliferation of cancer cells through various mechanisms. ILC1s can produce IFN-γ and granzymes to suppress tumor growth, but express immune checkpoints on their surface may impair their function and promote tumor growth [72]. Upon stimulation with IL-33, ILC2s can secrete high levels of IL-5, GM-CSF, and CCL5, which can activate and recruit anti-tumorigenic eosinophils, DCs, and CD8+ T cells [74]. In contrast, the production of immune checkpoint molecules like PD-1 and the cytokines IL-4 and IL-13 dampens ILC2 activity and accelerates tumor growth [75]. Under the regulation of TGF-β signaling, ILC3s can also convert to ILC1s or regulatory ILCs [76]. According to the type of tumor, ILC3s produce cytokines including IL-17 and IL-22 that promote or inhibit tumor development [77,78].
This entry is adapted from the peer-reviewed paper 10.3390/ijms241411233
This entry is offline, you can click here to edit this entry!