Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Dermatology
Bullous pemphigoid (BP) is an autoimmune blistering skin disease, of which the incidence has increased in recent years. BP is characterized by circulating IgG and IgE autoantibodies against the hemidesmosomal proteins BP180 and BP230.
- bullous pemphigoid
- basophils
- eosinophils
1. Introduction
Bullous pemphigoid (BP), which is the most common subepidermal autoimmune blistering disease [1], mainly affects the elderly and is associated with significant mortality and morbidity risks [2]. While the early phase of BP is clinically characterized by flares and sometimes wheals that are associated with a strong sensation of pruritus, flares in conjunction with tense, fluid-filled blisters are specific for the late stage of the disease [3]. Although BP is historically characterized by the production of IgG autoantibodies against the hemidesmosomal proteins BP180 and BP230 [4,5,6], studies in recent years have further elucidated the involvement of eosinophils, basophils, and neutrophils in the pathogenesis of BP. Now, infiltration of these immune cells into skin lesions is considered to be a predominant feature of BP.
2. Role of Eosinophils in the Pathogenesis of Bullous Pemphigoid
Eosinophilic granulocytes are bone marrow-derived cells of the myeloid lineage and were first described by Paul Ehrlich in 1879 [7]. The name was given due to the bright color when stained with eosin. Eosinophil granulocytes are primarily tissue-resident cells that act as key effector cells in inflammatory skin diseases and account for only 1–5% of circulating leukocytes in peripheral blood [8]. One of their functions is host defense, particularly in response to helminths and extracellular bacteria. Eosinophils bind and kill bacteria by generating extracellular DNA traps (EETs) that contain toxic granule proteins, such as the major basic protein (MBP), eosinophil-derived neurotoxin (EDN), and eosinophil cationic protein (ECP) [9,10].
The formation of a subepidermal cleft, accompanied by a significant eosinophilic infiltration, is a major histologic feature of BP [11,12]. Eosinophils have been shown to represent the prime early immune cells invading the skin in BP and are typically localized in the upper dermis of skin lesions in addition to lining the zone of the dermal–epidermal junction, but are also present in blisters and the peripheral blood of patients with BP [13]. Several studies demonstrate that eosinophils and their granule products (chemokines and cytokines) play a key role in the pathogenesis of BP [14,15,16] (Figure 1). Levels of various cytokines and chemokines, including interleukin (IL)-1β, IL-1α, IL-2, IL-4, IL-5, IL-6, IL-8, IL-15, IL-31, eotaxin, eosinophil colony-stimulating factor, tumor necrosis factor alpha (TNFα), and interferon-gamma (IFN-γ), are elevated in the sera and blister fluids of patients with BP [11,16,17,18,19,20,21]. Significant correlations have been determined for disease activity and blister fluid levels of IL-1β, IL-6, IL-8, TNF-α, and IL-5 [18,19].
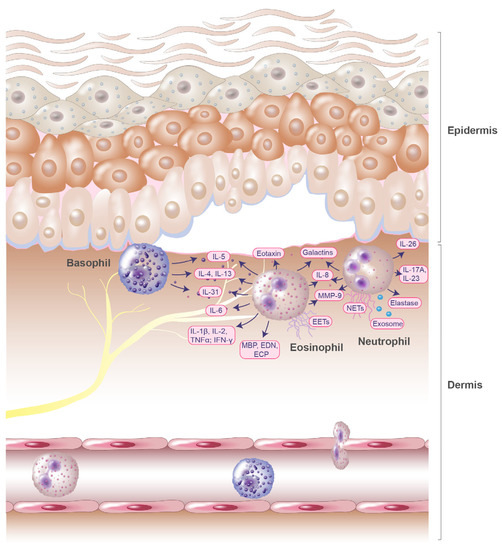
Figure 1. Potential role of eosinophils, basophils, and neutrophils in blister formation of bullous pemphigoid. Chemokine and cytokine gradients induce eosinophil, basophils, and neutrophil infiltration into the upper dermis. The inflammatory reaction is intensified by inflammatory mediators released from basophils, neutrophils, and eosinophils. Eosinophils release various cytokines, including the pruritic interleukin IL-31. Together with basophils, they secrete IL-4, IL-13, and IL-5. Additionally, eosinophils generate extracellular DNA traps (EETs) that contain toxic granule proteins, such as the major basic protein (MBP), eosinophil-derived neurotoxin (EDN), and eosinophil cationic protein (ECP). Eosinophils and neutrophils release IL-8, MMP-9, and galectins. Further, neutrophils release neutrophil extracellular traps (NETs) and exosomes.
BP blister fluids have been reported to contain not only products of eosinophils but also eosinophil chemotactic factors [37]. High eotaxin levels have been detected in BP blister fluid and were found to be significantly correlated with the number of dermal infiltrating eosinophils [21]. Immunohistochemical analysis conducted by Wakugava et al., demonstrated that eotaxin was strongly expressed in epidermal keratinocytes located around BP blisters [21], which have also been observed in other Th2-associated inflammatory skin diseases, such as atopic dermatitis [38]. These common expression patterns have been suggested to arise from eosinophils, which induce eotaxin-1 expression in keratinocytes through the release of ECP and EDN [39]. Moreover, blister fluids were found to contain high amounts of Th2 cytokines such as IL-5 [11,21]. IL-5 levels in serum exceeded those measured in blister fluids and were significantly increased in patients with BP. Since IL-5 and eotaxin play a major role in eosinophil recruitment and function [40], this may partly explain the massive infiltration of eosinophils observed in BP (Figure 1).
Pruritus is a major symptom in BP [41,42], suggesting a possible involvement of the pruritogenic cytokine IL-31. This was confirmed by Salz et al., by observing increased IL-31 concentrations in BP blister fluids [22]. The correlation of IL-31 levels with eosinophil numbers indicates that eosinophils might influence the regulation of IL-31 and itch in BP. More recently, our group was able to confirm these findings and additionally show that eosinophils are the major cellular source of IL-31 in BP [23]. These data further strengthen the hypothesis that eosinophils greatly contribute to itch in patients with BP via IL-31.
Serum concentrations of eosinophil secretory granule products, such as MBP, EDN, and ECP, are also significantly elevated in patients with BP [22,24,25,26]. This suggests that the eosinophils of BP patients are in an activated state, as eosinophils release the content of their secretory granules upon degranulation. Findings from a study by Tsuda et al., support this assumption, as increased numbers of activated eosinophils were observed in blood samples from patients with BP [43]. Hypodense (activated) eosinophils were also found to be located in the basement membrane zone of skin samples, and direct adherence of eosinophils to basal keratinocytes was observed at blistering lesion sites [43]. By releasing granule proteins, eosinophils may directly damage basal keratinocytes, leading to dermo-epidermal separation. Furthermore, ECP and EDN have been reported to induce the expression of cytokines and chemokines such as IL-5, eotaxin-1, RANTES, and MMP-9 in keratinocytes. Both granule proteins initiate the release of reactive oxygen species (ROS) from keratinocytes and trigger their apoptosis [39]. In line with these findings, we showed that eosinophils in peripheral blood, skin, and blister fluids exhibited strong activation patterns, which were determined through measurement of CD69 expression [17]. Apoptosis in cultivated BP eosinophils was found to be increased, and in vivo initiation of apoptosis by caspase-3-positive eosinophils was confirmed in lesional BP skin sections [17], assuming that activated eosinophils have a shorter life span due to the release of inflammatory mediators.
Additionally to releasing their secretory granules, eosinophils have also been observed to release EETs together with ECP in BP lesions [27]. Extracellular traps expand to up to 15 times the size of eosinophils and consist of network-like structures that contain DNA in conjunction with eosinophil granule proteins. Simon et al., analyzed 25 different eosinophilic skin diseases and detected extracellular DNA in skin samples from infectious and non-infectious inflammatory skin diseases. Interestingly, EETs were also found in skin biopsies of BP patients [27].
The proteome of BP blister fluids was characterized in a recent study, and 339 different proteins were identified, of which the eosinophil-derived proteins, MBP, eosinophil peroxidase, galectin-10, and the IgEε heavy constant region were unique and consistently associated with tissue eosinophilia [28]. The presence of relatively high levels of galectin-10 is particularly notable. Galectin-10, also known as Charcot-Leyden crystal protein (CLC-P), is a common protein of human eosinophils and a hallmark of active eosinophilic inflammation and proliferation [44]. However, galectin-10 is also produced by basophils [45], and the presence of galactin-10 in blister fluid may also be an indicator of basophil involvement in the disease process.
Matrix metalloproteinases (MMPs) are increased in BP skin lesions [46] and may be involved in the mechanism of blister formation in BP too. This is of interest since eosinophils release MMP-9 upon stimulation with TNF [47] and metalloproteinase activity has been detected in BP blister fluids [48]. The extent of eosinophil involvement in the process of dermal epidermal separation through the release of MMPs was shown in a study by Stahle-Bäckdahl et al. [30]. They demonstrated that the abundant component of BP blister fluid, MMP-9 (92 kD gelatinase), is produced by eosinophils and that MMP-9 degrades the collagenous domain of a recombinant form of the 180 kD bullous pemphigoid autoantigen in vitro. These results suggest that the production and release of MMP-9 by eosinophils contribute significantly to blister formation in BP [30]. An in vitro study by Okada et al., investigated the role of MMP-9 in eosinophil basement membrane migration in Matrigel-coated chemotaxis chambers [49]. Eosinophil transmigration and degradation of the Matrigel layer were induced by platelet-activating factor (PAF), interleukin-5 (IL-5), and MMP-9, identified as key proteases. Notably, the substrate-degrading activity of MMP-9 was increased only in the presence of IL-5 and PAF [49]. These data indicate that MMP-9 release and activation mechanisms are involved in basement membrane transmigration of eosinophils.
In a study by Ständer et al., it was observed that an eosinophil-predominant inflammatory infiltrate was more frequently observed in patients that presented with intense subepidermal splitting [51]. This further underlies the role of eosinophils in BP. In recent years, the number of studies presenting data that supports the pathogenic role of IgE autoantibodies in BP, as well as their relationship with eosinophils, has increased. A mouse model that studied humanized anti-BP180-NC16A IgE and the high-affinity IgE receptor (FcεRI) showed that IgE autoantibodies induce eosinophil infiltration followed by blister formation in a dose-dependent manner. The eosinophil infiltration was further correlated with higher doses of anti-BP180-NC16A IgE [52]. Moreover, a high density of eosinophils in lesional BP skin inflammatory infiltrates was associated with increased seropositivity of anti-BP180-NC16A IgG [51]. In non-bullous pemphigoid, peripheral eosinophilia was also found to be significantly associated with elevated total IgE levels [53].
Although various observations support the pathogenic role of eosinophils in BP, few studies have examined the association between the number of tissue eosinophils, disease severity, and response to treatment. In a retrospective cohort study that included 233 patients with BP, a significant correlation was reported between peripheral blood and cutaneous eosinophil numbers and disease severity, as well as treatment outcomes [54]. The authors conclude that eosinophils may be used as a marker to predict disease severity in patients with BP [54]. The association between the amount of eosinophils in skin biopsy samples of 137 patients with BP and their demographic characteristics, comorbidities, disease severity, and treatment response was also evaluated in a very recently published retrospective study by Baum et al. [55]. Contrarily, they found no correlations between eosinophil numbers and disease severity, but their quantity may serve as a prognostic factor for treatment outcome. Recent studies, however, show that the inflammatory early phase of BP is associated with an infiltration of basophils that correlates with eosinophils, indicating a role for both granulocytes in blister development and early inflammation [13].
3. Role of Basophils in the Pathogenesis of Bullous Pemphigoid
Basophils have also been detected by Paul Ehrlich and are relatively rare cells, that represent less than 1% of peripheral blood leukocytes in humans [56]. They are characterized by a constitutive expression of FcεRI [57,58]. Basophils are important early producers of type 2 mediators such as IL-4 and IL-13, which drive inflammation and pruritus [59,60,61,62]. The predominance of the cytokines IL-4, IL-5, and IL-13 that can be detected in biopsies of lesional skin from patients with BP [20] suggests that Th2-type immunity is involved in BP pathogenesis. Beside eosinophils, basophils are key effector cells of inflammatory skin diseases. Recent studies reveal that basophils play a more important role in inflammatory skin diseases than previously thought. Infiltration of basophils into the skin has been observed in several inflammatory skin disorders, including atopic dermatitis (AD), prurigo, and urticaria, and, interestingly, also in autoimmune skin diseases such as BP [63].
While the accumulation of eosinophils in the skin tissue is a common finding in BP [16], basophils also infiltrate BP skin lesions and are involved in the induction of Th2 immunity alongside eosinophils (Figure 1). The involvement of basophils in the development of IgE-mediated chronic allergic skin inflammation and increased serum IgE levels in patients with BP [64] further suggest a pivotal role for basophils in BP. However, there are very few studies investigating the role of basophils in the pathogenesis of BP.
Infiltration of basophils into lesional BP skin was first detected in a study by Dvorak et al., in 1982 [65], where the inflammatory response in lesions of a BP patient was evaluated at different stages of disease progression by electron and light microscopy. Infiltration of immune cells into lesions was initially described for lymphocytes and later for eosinophils and basophils. Light microscopy showed basophils mainly in clinically prominent lesions and adjacent normal skin. Intralesional eosinophil location was associated with proximity to degranulated basophils.
The extent of basophil infiltration into the skin in other inflammatory skin diseases was investigated in a study by Ito et al. [63]. Basophil infiltration was analyzed in 136 samples from 24 skin diseases through immunohistochemical staining. Contrary to previous assumptions, basophils were detectable in lesions of many diseases and conditions such as AD, prurigo, urticaria, drug eruptions, eosinophilic pustular folliculitis, insect bites, scabies, Henoch-Schönlein purpura, and dermatomyositis, as well as in BP. Interestingly, basophil infiltration was observed in nine of ten BP skin samples, a frequency that exceeds that seen in AD. Additionally, CD203c expression, a marker for blood basophil activation, was investigated by flow cytometry and found to be increased in patients with BP when compared to healthy donors [63].
The pathophysiological mechanisms of itch were investigated in BP patients in a recent study by Hashimoto et al. [66]. Lesions from 24 patients with BP were analyzed through immunofluorescence staining. The number of dermal infiltrating basophils was found to be significantly increased in BP skin sections compared to healthy controls. Interestingly, the number of dermal basophils significantly correlated with the severity of pruritus, highlighting the involvement of basophils in BP-associated itch. Beside basophils, eosinophils, SP, NK1R, IL-31, IL-31RA, OSMR, IL-13, and periostin were also correlated with the severity of pruritus, and eosinophils were identified as a major source of IL-31 [66].
The dual role of basophils in the development and resolution of BP was confirmed in a recent study by Kimura et al. [13], where erythematous and bullous lesions from a total of 25 patients with BP were compared by histopathology, immunohistochemistry, and electron microscopy. The number of basophils in the early phase of BP was found to be positively correlated with the number of eosinophils. Interestingly, basophil numbers were significantly higher in the bullous phase of BP.
This entry is adapted from the peer-reviewed paper 10.3390/biom13071019
This entry is offline, you can click here to edit this entry!