DNA replication is fundamental to the maintenance and diversification of life. In eukaryotic cells, DNA replication initiates from multiple origins deployed across the whole genome, implying the need for a tight orchestration of their firing. Complex multi-step regulatory mechanisms coordinate such efforts and ensure that the genome is fully duplicated. Notably, a large excess of DNA replication origins are present throughout the human genome, with only 5–10% of them firing throughout S phase. Based on their usage, DNA replication origins are classified into the following three categories: (1) constitutive origins that invariably fire in all cells of a population, (2) flexible origins (the majority) that only fire in some cells of a population, and (3) dormant origins that are kept silent during normal conditions but can become activated upon DNA damage when a replication fork stalls in the vicinity.
1. Introduction
Eukaryotic DNA replication initiation follows a conserved process that leads to the assembly of two replisomes, which will move away from the activated origin in opposite directions [
4]. In budding yeast, at the core of the replisome is a hexameric DNA helicase motor, composed of the minichromosome maintenance subunits 2 to 7 (MCM2-7), in the shape of a ring that unwinds parental double-stranded DNA (dsDNA) [
5]. To avoid genome re-replication and thus strictly limit replication to once per S phase, the activation of DNA replication origins entails a two-step process consisting of (1) DNA helicase loading onto DNA and (2) DNA helicase activation, which occurs at temporally distinct times. The first step takes place in the G1 phase via the “licensing” of all replication origins, rendering them competent for firing. The second step occurs in the early S phase via the activation of the DNA replicative helicase and the formation of two active replication forks through the regulated sequential recruitment of firing factors (
Figure 1). The temporality of this two-step process is governed by the activity of global cell cycle regulators, mainly the Ser/Thr protein kinases CDKs (cyclin-dependent kinases) and DDK (Dbf4-dependent kinase, also known as CDC7-DBF4), ensuring that the DNA replication program is coupled with cell growth conditions and environmental cues [
6]. A recent study also demonstrated that CDK1 plays a redundant role with CDC7 in the G1/S transition [
7].
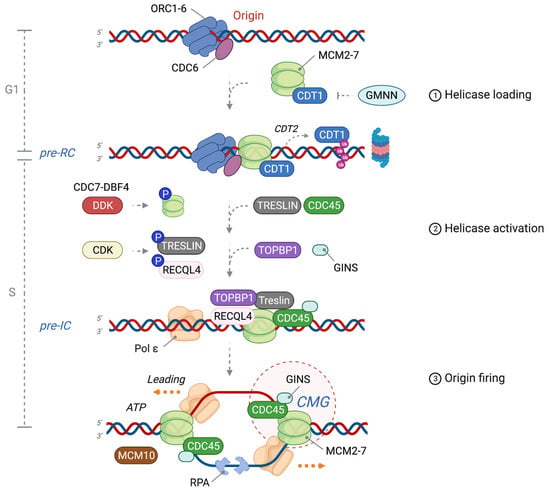
Figure 1. Licensing and activation of a replication origin. The replication origin of eukaryotes is first marked by the origin recognition complex 1-6 (ORC1-6), which recruits cell division cycle 6 (CDC6) and subsequently loads minichromosome maintenance 2-7 (MCM2-7)/Cdc10-dependent transcript 1 (CDT1) to form the pre-replication complex (pre-RC); one of two double hexamers is shown. An excess of MCM2-7 hexamers is loaded into DNA during the G1 phase, constituting dormant replication origins. Licensing occurs once per cell cycle and is restricted to G1 by the inhibitory interaction of CDT1 with GEMININ (GMNN) during S and G2 phases (in vertebrates), as well as proteasomal degradation of CDT1. The pre-initiation complex (pre-IC) is formed by DBF4-dependent kinase (DDK)-mediated phosphorylation of MCM2-7, which promotes binding of TRESLIN and CDC45 to replication origins. CDKs subsequently phosphorylate TRESLIN and RECQL4, thereby promoting the recruitment of TOPBP1 and Go-Ichi-Ni-San (GINS1-4) complex to assemble the CMG (CDC45-MCM2-7-GINS) helicase complex together with polymerase ε (Pol ε). Origin activation: engagement of pre-IC with MCM10 initiates melting of dsDNA and unwinding by the CMG. RPA binds and protects resulting ssDNA, and two separate replisomes are established to synthesize nascent DNA in a 5′-3′ direction. ℗: phosphorylation.
2. Origin Licensing: Pre-RC Formation in G1 Phase
DNA replication origins are first recognized by the heterohexameric origin recognition complex (ORC1-6) that possesses DNA-binding domains and exhibits AAA
+ ATPase activity. In mammals, ORC1 and ORC6 weakly interact with the ORC2-5 core, thus forming a dynamic open ring that binds to DNA through its central cavity [
8]. ORC1 is loaded onto DNA as early as in mitosis and is degraded during the S phase through the ubiquitin-proteasome system, suggesting that it promotes the timely assembly of the ORC complex at its origins [
9]. In yeast, Orc1-6 binding to origins induces a small bend in dsDNA followed by the binding of another ATPase, Cdc6, thus closing the Orc ring around duplex DNA [
10]. Then, CDT1 (CDC10-dependent transcript 1) is recruited to ORC1-6/CDC6 bound origins, forming the platform necessary to recruit and sequentially load two MCM2-7 hexamers in a head-to-head orientation using ATP hydrolysis. In yeast, Cdt1 forms a complex with MCM2-7 before being loaded onto origins [
11]. At this stage, the MCM double hexamer is inactive as it encircles duplex DNA. Together, this assembly of proteins forms the pre-replication complex (pre-RC) at the licensed replication origins. Importantly, helicase loading can only occur in the G1 phase during low CDK activity because of the inhibitory effect of the direct phosphorylation of ORC, CDC6 or MCMs [
12,
13,
14]. The CRL4
CDT2 ubiquitin E3 ligase targets CDT1 for degradation in a proliferation cell nuclear antigen (PCNA)-dependent manner to prevent DNA re-replication [
15]. Additionally, CDT1 is inhibited through sequestration by the S phase-specific inhibitor GEMININ, thus preventing pre-RC assembly [
16].
3. DNA Helicase Activation: Pre-IC Formation at the G1-S Phase Transition
The activation of the replicative DNA helicase requires a series of allosteric changes that only occur during high CDK activity in the S phase, allowing for the phosphorylation and recruitment of firing factors. Importantly, DDK transiently binds to the pre-RC and phosphorylates key sites on the double hexamer, most notably on the MCM4 and MCM6 subunits, which induces structural changes that allow for the binding of CDC45 (cell division cycle 45) and GINS heterotetramer (Go-Ichi-Ni-San, meaning 5-1-2-3 in Japanese) composed of Sld5 (synthetic lethal with Dpb11), Psf1 (partner of Sld5-1), Psf2, and Psf3 [
17,
18,
19]. Simultaneously, additional phosphorylation events by CDKs and DDK of critical firing factors, such as TRESLIN, ATP-dependent DNA helicase Q4 (RECQL4; Sld3 and Sld2 in
S. pombe), DNA topoisomerase 2-binding protein 1 (TOPBP1), and DNA polymerase ε (pol ε), collaboratively contribute to the assembly of the pre-initiation (pre-IC) complex with two replicative helicases formed by the tight CMG (CDC45-MCM2-7-GINS) complex at its core [
20,
21,
22,
23,
24]. A recent systemic chromatin immunoprecipitation from budding yeast revealed distinct intermediates of the pre-IC assembly that are mutually dependent on the origin firing factors [
25].
4. Origin Firing: Formation of Two Functional DNA Replication Forks
The CMG formation results in the release of ADP from the MCM double hexamer, which allows binding of ATP, thus triggering the initial untwisting of a short stretch of DNA and the separation of two inactive CMG complexes. MCM10 and ATP hydrolysis are instrumental for helicase activity, such that the two replisomes pass each other by translocating 3′ to 5′ on the leading strand template (with their N-terminus at the front of the helicase) after evicting the lagging strand template from the MCM pore [
26,
27]. Replication protein A (RPA) quickly binds to the exposed single-stranded DNA (ssDNA) after DNA melting, while CDC6 and CDT1 are evicted and inactivated. DNA polymerases α and δ are then recruited along with replication factor C (RFC) and PCNA to convert the pre-IC into two active replication forks, which move in opposite directions from the activated origin. DNA is thus replicated in a semi-conservative fashion, while deoxyribonucleoside triphosphates (dNTPs) are incorporated at daughter strands, in which the leading strand is copied continuously by DNA Pol ε, while the lagging strand is copied discontinuously as a succession of Okazaki fragments initiated by DNA Pol α primase (RNA primer) and elongated by DNA Pol δ [
4].
This entry is adapted from the peer-reviewed paper 10.3390/ijms241310488