Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Others
There is no doubt that significant progress has been made in tumor therapy in the past decades. The discovery of new molecules with potential antitumor properties still remains one of the most significant challenges in the field of anticancer therapy. Nature, especially plants, is a rich source of phytochemicals with pleiotropic biological activities.
- natural chalcones
- antiproliferative
- anticancer
1. Introduction
Cancer is one of the leading causes of death worldwide with 10 million deaths in 2020 [1]. Moreover, according to GLOBOCAN 2020, a 47% increase in the incidence of cancer disease is expected over the next 20 years [2]. Despite significant progress in the diagnosis and treatment of cancer diseases, cancer still remains a disease that is often difficult to cure [3]. In addition, “classical” chemotherapy is often associated with severe toxicity, mostly resulting from low selectivity against cancer cells. Therefore, the discovery of new, effective compounds with low toxicity still remains a great challenge.
Plants are an amazing source of active compounds with a significant impact on human health. The effects of plant products on health are primarily determined by the presence of secondary metabolites produced by the plant for protection against various external biotic and abiotic factors. Among them, polyphenols have attracted the attention of researchers due to their diversity of biological effects [4]. Based on their chemical structure, polyphenols are categorized into two classes: flavonoids and non-flavonoids. Of these, flavonoids are the most prevalent polyphenols, with several thousand members [5]. They can be divided into different groups, such as flavones, flavonols, flavanones, isoflavones, and anthocyanidins. In addition to these basic flavonoids, there are other minor groups, including chalcones and dihydrochalcones [6].
Chalcones, 1,3-diphenyl-2-propene-1-ones, are precursors for flavonoids and isoflavonoid synthesis in plants. They have relatively simple chemical structures but exhibit a great diversity of actions. The chemical structures of some chalcones cited in the text are shown in Figure 1.
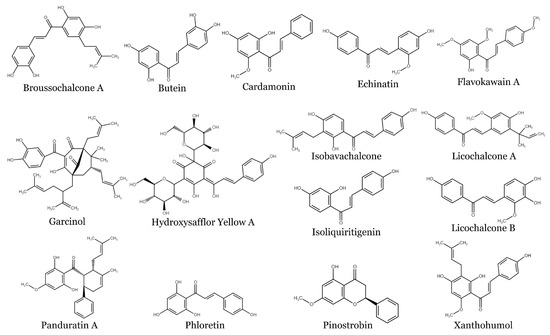
Figure 1. Structures of selected naturally occurring chalcones.
Except for plant-related functions, chalcones have been found to possess a broad spectrum of biological activities also in humans including antimicrobial, antidiabetic, cardioprotective, anti-inflammatory, neuroprotective, and antioxidant [7,8,9] effects (Figure 2). In addition to these actions, chalcones have also attracted attention due to their ability to modulate the process of carcinogenesis at different levels [10,11]. Regarding anticancer effect, we and others have reported the growth-inhibitory effect of numerous chalcones in vitro [12,13,14,15] or the anticancer effect in vivo [16,17]. Additionally, multimodal pharmacological effects of chalcones on cellular and molecular levels have been recently reviewed [18]. In the present paper, we discuss the antiproliferative and anticancer effects of natural chalcones in solid cancers, including their cellular and molecular mechanisms of action.
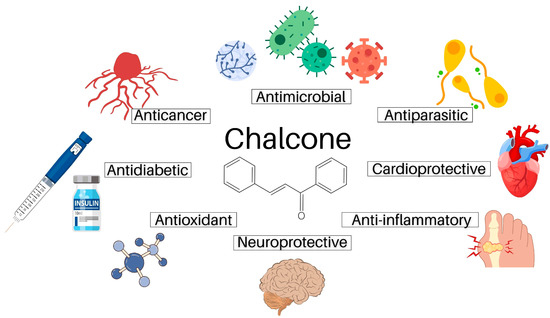
Figure 2. The pharmacological activities of natural chalcones.
2. Chalcones and Breast Cancer
Breast cancer is a malignancy preferentially affecting women with high incidence and mortality rates. It is a complex, highly heterogeneous disease with two kinds of tumor heterogeneity: inter-tumor and intra-tumor heterogeneity [19]. Therefore, classification of breast cancer is complicated and includes primary tumor, lymph node and metastasis evaluation as well as important molecular markers. These biological factors imply immunohistochemical markers, e.g., estrogen and progesterone receptor (ER, PR), human epidermal growth factor receptor 2 (HER2), and proliferation marker protein Ki-67; genomic markers, e.g., breast cancer susceptibility proteins (BRCA1, BRCA2), and phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha (PIK3CA); and immunomarkers, e.g., tumor-infiltrating lymphocytes (TILs) and programmed death-ligand 1 (PD-L1) and receptors [20]. However, among others, triple-negative breast cancer (TNBC) and inflammatory breast cancer (IBC) are considered the most aggressive types of breast cancer, with no significant treatment improvement [21]. The major obstacle to achieving effective treatment is resistance to therapies. Therefore, the therapy options and search for novel effective agents remain current. Chalcones have been shown to express different activities suppressing malignant cells, including breast cancer cell proliferation, adhesion, migration and ability to metastasize. This is why they have been classified as promising anticancer agents in breast cancer.
2.1. Xanthohumol
Xanthohumol (XH) is a prenylated chalcone known for being isolated from the hops plant (Humulus lupulus) and for its numerous biological activities, such as antimicrobial, antiviral, anti-inflammatory, immunomodulatory, and anticancer effects [22,23].
Besides other human tumors, it inhibits the proliferation of different breast cancer cells. The effect of XH has been investigated in both in vitro and in vivo studies. Initial studies in MCF7 cell lines revealed that XH decreased cell proliferation in a dose-dependent manner. The following oral administration to nude mice inoculated with MCF7 cells resulted in central necrosis within tumors, reduced inflammatory cell number and increased percentage of apoptotic cells. Anti-angiogenic effect of XH was approved by decreased microvessel density and factor VIII expression in XH-treated tumors. Decreased NF-κB activity, phosphorylated inhibitor of kappa B, and interleukin-1b also indicated that XH was able to target both breast cancer and inflammatory cells, as well as endothelial cells [24]. In addition, the actions of XH should be also mediated by the effect on cellular mitochondrial function. This was demonstrated by measuring MCF7 cell viability and reactive oxygen species (ROS) production. XH exerted a dual effect on these breast cancer cells. In low doses (0.01 mM) XH decreased ROS production along with an increase in oxidative phosphorylation system expression. In contrast, high concentrations of XH (5 mM) resulted in increased ROS production, which was accompanied by a decrease in oxidative phosphorylation system expression. The authors suggested that XH in a low dose probably improved, whereas a high dose worsened the mitochondrial function [25]. Mitochondrial effects of XH were then evaluated in another study with the MDA-MB-231 breast cancer cell line. Treatment of cells with 10 μM and 20 μM XH for 48 h significantly decreased the rate of survival with a concurrent increase in the number of sub G0/G1 cells. Observed DNA fragmentation was accompanied by increased expression of Bax and the activity of caspase-3 and -9 in the mitochondria, suggesting the inhibition of cell proliferation through mitochondria- and caspase-dependent apoptotic pathways [26].
In ERα-positive breast cancer cells, XH also works by inhibiting tumor suppressor protein prohibitin 2 binding to brefeldin A inhibited guanine nucleotide-exchange protein 3 (BIG3). This inhibition released prohibitin 2 to directly bind to both nuclear and cytoplasmic ERα. The effect was followed by complete suppression of the estrogen-signaling pathways and ERα-positive breast cancer cell growth both in vitro and in vivo. No such effects were observed in normal mammary epithelial cells [27]. Besides ERα-positive breast cancer cells, XH also effectively inhibited the growth and proliferation of TNBC cells. Evaluation of different chalcones have shown that treatment with chalcone, 2-hydroxychalcone, and XH for 24 h inhibited the growth of MDA-MB-231 cells with IC50 values of 18.1, 4.6, and 6.7 μM, respectively. Similar cytotoxicity was observed in another TNBC cell line, Hs578T. Both XH and 2-hydroxychalcone induced apoptosis by Bcl-2 downregulation. Moreover, they exerted more potent inhibitory effects on the proliferation, matrix metalloproteinase (MMP-9) expression and invasive phenotype of MDA-MB-231 than chalcone [28]. Similar results were obtained by comparing MCF-7 and MDA-MB-231 cells in vitro and in 4T1 breast tumor mouse in vivo model. XH significantly decreased cell viability, induced G0/G1 cell cycle arrest and apoptosis in vitro and inactivated the Notch signaling pathway, Notch 1 and Ki-67, in vivo [29]. In vivo studies on a BALB/c-4T1 breast cancer mouse model confirmed antiproliferative effects of XH in another TNBC cell line. In addition to decreased proliferation, XH inhibited expression of proliferation marker protein Ki-67 as well as breast cancer-specific marker cancer antigen 15-3 in this highly tumorigenic and invasive cell line. At the same time, XH enhanced the secretion of perforin, granzyme B, increased the ratio of CD8+/CD25+ and upregulated Th1 cytokines, while it downregulated Th2 cytokines, resulting in a markedly elevated ratio. Because Th1/Th2 is closely related to a variety of tumors, XH-induced enhancement of Th1 immunity response could appear as a more effective approach in mediating anticancer function [30].
Development of drug resistance, which usually goes together with resistance to ionizing radiation results in serious breast cancer treatment problems. To find a method to overcome this obstacle, several natural and synthetic compounds, including XH, were investigated. When the effect of XH on doxorubicin-resistant breast cancer cells (MCF-7/ADR) was investigated, it inhibited viability, induced apoptosis, and arrested the cell cycle of these cells in a dose-dependent manner. Moreover, it increased the inhibitory effect of doxorubicin on MCF-7/ADR cells. These processes were accompanied by decreased colony formation, migration, sphere formation, and the downregulation of stemness-related biomarkers. All these effects indicated the efficiency of XH in the regulation of stemness of doxorubicin-resistant breast cancer cells [31]. The main aim of the experiment with XH, which was performed on MCF-7 and MCF-7/ADR cells, was to determine its radio-sensitizing activity. The results revealed that pretreatment of the cells with XH significantly sensitized MCF-7/ADR cells to radiation treatment by inducing apoptosis. The effect was accompanied by decreased expression of anti-apoptotic proteins, such as MDR1, epidermal growth factor receptor (EGFR) and signal transducer and activator of transcription 3 (STAT3) and increased expression of death receptors DR4 and DR5. All these results suggest that XH pretreatment restores the sensitivity of MCF-7/ADR cells to doxorubicin and radiation therapies [32].
2.2. Butein
Butein, 2′,3,4,4′-tetrahydroxychalcone, found in several plants such as the heartwood of Dalbergia odoriferau or the stem bark of Semecarpus anacardium, has been reported to possess several beneficial properties, such as antioxidant, anti-inflammatory, and antimicrobial activities [33,34]. Moreover, numerous studies have revealed that butein exhibits antiproliferative and anticancer effects in vitro and in vivo [35].
Direct actions of butein on breast cancer cell proliferation result from modulation of the effect of different cellular proteins. It has been suggested that cyclooxygenase (COX) inhibitors could be used in the chemoprevention of breast carcinogenesis. In addition, COX-2 isoform was found to be overexpressed in breast cancer tissues. Butein (at or below 10 μM) downregulated phorbol 12-myristate 13-acetate (PMA)-induced COX-2 expression in both cancerous and non-cancerous breast cells. This effect was mediated by transcriptional inhibition of this gene [36]. Similarly, CXC chemokine receptor-4 (CXCR4) is expressed in various tumors. This receptor mediates homing of tumor cells to specific organs expressing the ligand CXCL12 and plays an important role in tumor growth, invasion, metastasis, and angiogenesis. Butein downregulated the expression of CXCR4. This effect correlated with the inhibition of CXCL12-induced migration and invasion of breast cancer cells. However, the decrease in CXCR4 expression induced by butein was not cell type-specific, and also occurred in pancreatic, prostate, multiple myeloma, head and neck, and hepatocellular cancer cell lines [37]. Controversial effects of butein on ROS production in breast cancer cells have been observed. An increase in ROS production was described in MDA-MB-231 (TNBC) cells. This resulted in decreased phosphorylation of extracellular signal-regulated kinase (ERK), increased p38 activity, diminished Bcl-2 expression, induced caspase 3 cleavage and was associated with poly ADP-ribose polymerase (PARP) cleavage and inhibition of breast cancer cell proliferation [38]. Two years later, Cho et al. demonstrated that butein inhibited ROS production with subsequent breast cancer growth suppression. These experiments were performed both in vitro and in vivo with butein-sensitive or -resistant breast cancer cells. Butein reduced the viabilities of different breast cancer cells, while not affecting resistant cells HER2 positive HCC-1419 (human cell line isolated from primary ductal carcinoma), SKBR-3 (human breast cancer cell line that overexpresses the Her2 gene product) and HCC-2218 cells (epithelial fibroblast cell isolated from the mammary gland ductal carcinoma). This was attributed to butein mediated inhibition of ROS levels resulting in the inhibition of AKT phosphorylation, as ROS regulate AKT activity and vice versa. The authors concluded that butein suppression of breast cancer growth was correlated with its reduction of the levels of both ROS and phosphorylated AKT, in vivo [39]. Additional experiments revealed that butein binds to a specific pocket of estrogen receptor (ERα) and promotes proteasome-mediated degradation of this receptor. Degradation results in butein-induced ERα downregulation, cell cycle arrest and inhibition of the growth of ERα+ breast cancer cells both in vitro and in vivo [40].
Indirect actions of butein in the modulation of breast cancer cell proliferation are represented by its inhibitory effect on estrogen production by aromatase inhibition apart of malignant breast cancer cells. Butein was found to be the strongest aromatase inhibitor among 5 tested hydroxychalcones with its IC50 value of 3.75 μM [41]. Another indirect effect appeared when breast cancer cells were co-cultured with fibroblasts, believed to play an important role in promoting the growth of breast cancer cells. Butein was found again as the most potent inhibitor of clonogenic growth of UACC-812 breast cancer cells isolated from the mammary gland ductal carcinoma. However, only when they were co-cultured with butein-pretreated fibroblasts [42]. Butein also reduced cell proliferation rate and the release of proinflammatory cytokines in two TNBC cell lines. However, cytokine release was suppressed only in MDA-MB-231 (Caucasian), and not in MDA-MB-468 (African American) cells, indicating different responses to butein treatment [40]. Similar results were also obtained in cardamonin evaluation in the same breast cancer cell lines (see below). Sensitive MDA-MB-231 breast cancer cells were then used to test the combination of butein with frondoside-A (triterpenoid glycoside from the Atlantic Sea Cucumber). The tests were performed both in vitro and in vivo CAM assay (on chick embryo chorioallantoic membrane) and revealed that butein alone significantly reduced cancer cell viability and colony growth, as well as migration and invasion. The effects were due to the potent inhibition of STAT3 phosphorylation (signal transducer and activator of transcription 3), leading to PARP cleavage and consequent cell death. Combination of butein with frondoside-A led to additive effects [43].
2.3. Isoliquiritigenin
Isoliquiritigenin (2′,4′,4-trihydroxychalcone, ISL), a natural chalcone originally isolated from licorice root, has been found to have a broad spectrum of biological activities including anticancer activity [44]. Among other effects, it exhibits anti-proliferative, anti-angiogenic, and anti-invasive effects in breast cancer cells. It also acts as an estrogenic agonist of both ER isoforms and shows a dual effect in different cancer cells at different concentrations. Growth promotion is typical for low and intermediate ISL concentrations in hormone-sensitive MCF7 breast cancer cells. On the other hand, at a high level, ISL becomes cytotoxic in both ER-receptor positive as well as negative cells [45]. Antitumorigenic effects of ISL are mediated by different mechanisms and affect several cellular processes. Diminished cell viability, 5-bromo-2′-deoxyuridine (BrdU) incorporation, clonogenic ability and subsequent apoptosis were demonstrated in both MCF-7 and MDA-MB-231 cells breast cancer cells. Among others, these growth inhibitory effects and apoptosis were attributed to the downregulation of arachidonic acid metabolic network and the deactivation of the phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) signaling pathway, which is considered one of the most important pathways regulating cell proliferation, the cell cycle and apoptosis. The results were supported by the finding that ISL inhibited mRNA expression of multiple forms of arachidonic acid-metabolizing enzymes as well as downregulated the levels of several phosphorylated forms of cell cycle regulating proteins [46]. An additional mechanism of cancer cell growth inhibition could be the ISL-induced block of β-catenin transcription activity. This mechanism, which significantly limits the side population and cancer stem cell (CSC) ratios in breast cancer cells, could have a synergistic effect with chemotherapeutic drugs. It could inhibit breast cancer cell proliferation and colony formation and enhance breast CSC chemosensitivity with little toxicity in normal tissues and mammary stem cells [47].
It is well accepted that microRNAs (miRNA) regulate tumor progression by modulation of the expression of several oncogenes and tumor suppressor genes. The effect of ISL on miRNAs’ regulation of tumorigenesis in breast cancer was evaluated in subsequent papers. The results revealed that ISL downregulated miR-374a. This is one of the main miRNAs involved in the migration and invasion of cancer cells. A decrease in miR-374a resulted in increased tumor suppressor gene PTEN expression (phosphatase and tensin homolog—tumor suppressor), and an inhibition of aberrant Akt signaling [48]. On the other hand, ISL increased miR-200c in BT-549 (papillary, invasive ductal tumor) and MDA-MB-231 TNBC cells. The effect correlated with subsequent ISL–induced inhibition of metastasis and tumor growth in nude mice models and was mediated by decreased c-Jun expression through the increase in miR-200c [49]. Several other mechanisms of ISL action in TNBC were discovered. ISL induced cell apoptosis, reduced Bcl-2 protein expression, increased Bax protein level and activated caspase-3 and PARP. Moreover, it reduced the expression of total and phosphorylated mammalian target of rapamycin (mTOR). All these effects resulted in the inhibition of TNBC MDA-MB-231 breast cancer cell growth through autophagy-mediated apoptosis [50].
Inhibition of angiogenesis is another possibility as to how to decrease cancer cell growth and proliferation. Some chalcones can inhibit neovascularization, influencing the effect of hypoxia-inducible factor-1a (HIF-1α), a signaling molecule in a central axis, activating oncogenic signaling. It also acts as a metabolic switch in endothelial cell (EC)-driven tumor angiogenesis. For example, ISL suppressed sprout formation from VEGF-treated aortic rings and inhibited cancer angiogenesis via significant inhibition of VEGF-receptor expression. This inhibition was mediated via the promotion of HIF-1a proteasome degradation [51].
The potential of ISL was also investigated as a preventive and therapeutic agent for breast cancer cell-induced metastatic bone destruction. ISL exerted its effect through the proteins directly involved in the differentiation, activity, and survival of osteoclasts. At non-toxic concentrations, it significantly inhibited the RANKL/OPG (receptor activator of nuclear factor kappa-B ligand/osteoprotegerin) ratio by reducing the production of RANKL and restoring OPG production in hFOB1.19 osteoblast cell culture stimulated with conditioned medium of MDA-MB-231 cells. ISL also reduced the expression of COX-2 in similarly stimulated hFOB1.19 cells. These effects may result in inhibitory potential of ISL on metastatic breast cancer-induced bone destruction [52].
2.4. Cardamonin
Cardamonin (CAR) is a chalcone isolated from Alpiniae katsumadai. It has been described as an anti-inflammatory and anti-tumor agent in breast, lung, colon, and gastric human cancer cell lines, in both in vitro culture systems as well as xenograft mouse models [53]. Anticancer activities were demonstrated in different breast cancer cells including drug resistant cancer stem cells [54], as well as in TNBC [55]. Cytotoxic effects of CAR in TNBC cells were attributed to the modulation of Bcl-2, Bax, cyt-C, cleavage of caspase-3 and PARP, with subsequent induction of apoptosis and cell cycle arrest. CAR also reversed epithelial–mesenchymal transition (EMT) and downregulated invasion and migration of BT-549 cells. In in vivo experiments, CAR significantly inhibited the tumor volume at dose of 5 mg/kg-treated mice [55]. Additional findings indicated that CAR inhibited the growth of the TNBC cell line MDA-MB-231 in vitro and in vivo by suppressing HIF-1α mediated cell metabolism. Suppression of HIF-1α enhanced mitochondrial oxidative phosphorylation and induced ROS accumulation [56]. The crucial role of ROS in the inhibition of CAR-induced breast cancer cell proliferation was confirmed after quenching of ROS by addition of N-acetyl-cysteine (NAC) or overexpression of catalase which also blocked CAR-induced cell cycle arrest and apoptosis [57]. An additional mechanism of antiproliferative action of CAR was observed in HeLa and MCF-7 breast cancer cells resistant to mTOR inhibitors. CAR overcame the resistance by mechanism of mTOR inhibition which differed from the currently available mTOR inhibitors. It decreased the expression of regulatory associated protein of mTOR (Raptor), the effect which resulted in inhibition of cell proliferation and decrease in phosphorylation of mTOR and S6K1 (ribosomal protein S6 kinase B1) in the mTOR inhibitor resistant cells [58]. Concerning drug-resistance, the effects of CAR were also evaluated in MDA-MB-231 (Caucasian) and MDA-MB-468 (African American) breast cancer cells. Both TNBC cell lines upregulated the expression of PD-L1, known to inhibit immune system control of cancer cell proliferation. CAR treatment caused a dose-dependent decrease in cell viability in both cell lines and downregulated PD-L1 expression in MDA-MB-231 cells. In MDA-MB-468 cells, CAR had an opposite effect, upregulating the expression of PD-L1 [59]. Besides having a direct effect on tumor cell proliferation, miRNAs also modulate the expression of genes that regulate tumor angiogenesis. These miRNAs have aberrant expression profiles in many different cancers, including breast cancers. CAR and some other phytochemicals were found to exhibit their anti-angiogenic properties by targeting the miRNAs that regulate EC metabolism [60].
2.5. Licochalcone A
Licochalcone A (LCA) (3-dimethylallyl-4,4′-dihydroxy-6-methoxychalcone) is a phytoestrogen extracted from licorice root. It expresses antioxidant, antibacterial, antiviral, antiparasitic and antitumor activities [61,62,63]. Former testing in several cell lines demonstrated that LCA induced apoptosis in the MCF-7 cell line. It decreased the anti-apoptotic protein Bcl-2 and altered the Bcl-2/Bax ratio in favor of apoptosis [64]. Additionally, it reduced the expression of cyclin D1 and promoted the cleavage of PARP. These effects preceded the cell cycle arrest at the G1 phase and induced apoptosis mediated by the intrinsic pathway [65]. Additional evaluation of the anticancer effects of LCA in MCF-7 cells revealed that the agent significantly decreased cell viability and promoted autophagy and apoptosis. The actions were attributed to the suppression of PI3K/Akt/mTOR signaling pathway [66]. Similar effects of LCA were described in MDA-MB-231 cells. It inhibited cell proliferation and cell cycle, modulated mitochondrial membrane potential and DNA damage, and reduced oxidative stress. It also activated cleaved-caspase 3 and 9, significantly decreased Bcl-2 expression and ultimately caused the release of cytochrome c from the mitochondria into the cytoplasm [67]. Comparing MCF-7 and MDA-MB-231 cells, LCA showed dysfunction of mitochondrial membrane potential and mitochondrial ROS production in both cell lines. Subsequent anti-proliferative and apoptotic effects passed the intracellular mitochondrial apoptosis pathway through regulation of Sp1 transcription factor and apoptosis-related proteins [62].
Moreover, it has been documented that LCA significantly suppresses breast cancer cells migration and invasivity. The effect of LCA on breast cancer cell invasion and migration was investigated in MDA-MB-231 cells. It was demonstrated that LCA effectively suppressed cell migration and invasion, and modulated E-cadherin and vimentin expression by blocking MAPK and AKT signaling [67]. Furthermore, LCA downregulated important genes associated with cancer development in MCF-7 and BT-20 breast cancer cell lines, including the AURKA protein (a member of a family of mitotic serine/threonine kinases). Moreover, they inhibited cell migration of metastatic BT-20 cells derived from a TNBC invasive ductal carcinoma. Reduction of MDR-1 protein by LCA was also observed. These findings indicate the anti-cancer, anti-metastatic and anti-resistance potential of both chalcones [68]. Recently, Gong et al. reported that protein arginine methyltransferases (PRMT) could be another molecular target of LCA. PRMT mediate arginine methylation implicated in multiple biological functions including transcriptional regulation. These enzymes were found to be upregulated in various cancers. Expression of PRMT6 is upregulated in human breast cancers and it is associated with oncogenesis. Studies of LCA effects in MCF-7 cells showed that it reversibly and selective inhibited PRMT6. This inhibition resulted in cytotoxicity by upregulating p53 expression and blocking cell cycle progression at a G2/M phase, followed by apoptosis. However, the agent was not effective towards non-cancer MCF-10A human breast epithelial cells [69].
2.6. Flavokawains
Flavokawain A (FKA) is a chalcone isolated from the root extracts of the kava-kava plant (Piper methysticum), belonging to the Piperaceae family. The extract is traditionally known as the Pacific elixir by the Pacific islanders for its role in a wide range of biological activities [70]. The anti-cancer properties of FKA in MCF-7 and MDA-MB231 cells were evaluated by measuring several parameters, indicating both apoptotic and metastatic effects. It induces apoptosis in both cell lines in a dose dependent manner through the intrinsic mitochondrial pathway. The migration and invasion process in MDA-MB231 was also inhibited. Similar effects were seen in the inhibition of the angiogenesis process performed in human umbilical vein endothelial cells (HUVECs) via tube formation assay and ex vivo rat aortic ring assay [71]. In addition, it has been shown that FKA preferentially reduces the viability of HER2-overexpressing breast cancer cell lines. Applied at cytotoxic concentrations to breast cancer cell lines, it had a minimal effect on the growth of non-malignant breast epithelial MCF10A cells. In combination with monoclonal anti-HER2 antibody trastuzumab, FKA enhanced its growth inhibitory effect and downregulated several transcription factors and cell cycle regulating proteins [72].
The immunomodulatory effects and the anti-inflammatory effects of FKA in a breast cancer murine model demonstrated an increased number of T cells (both Th1 cells and cytotoxic T lymphocytes) and elevated levels of IFN-γ and IL-2 in the serum. In the same time, FKA-treated mice had reduced levels of major pro-inflammatory mediators and decreased weight and volume of the tumor caused by apoptosis induction. All these effects indicated enhancement of antitumor immunity and prevention of inflammatory process in tumor microenvironment [73].
Similar in vivo antitumor and antimetastatic effects were also described in flavokawain B (FKB)-treated mice. The results were obtained in 4T1 (highly tumorigenic and invasive tumor cell line) tumors in mice. FKB regulated the immune system by increasing both helper and cytolytic T-cell and natural killer cell populations. It also enhanced the levels of IL-2 and IFN-γ, induced apoptosis and inhibited metastasis [73].
2.7. Garcinol
Garcinol, a polyisoprenylated chalcone containing two aromatic rings separated by a carbonyl group, is a natural agent extracted from the rind of the fruit of Garcinia indica (known as mangosteen) [74]. Several experiments have suggested its anti-cancer activity in breast cancer cells. Recently, a review article aimed to analyze the potential of Garcinia phytochemicals as a molecular therapy of breast cancer, evaluated the results of 28 article selected studies. The analysis showed that phytochemicals of Garcinia, including garcinol, have anti-cancer properties, resulting from apoptosis, inhibition of proliferation, and metastasis in breast cancer cells [75]. The mechanisms involved in its anti-cancer effects included the reversal of EMT associated with the upregulation of epithelial marker E-cadherin and downregulation of some mesenchymal markers including vimentin, ZEB1 and ZEB2. In addition, garcinol upregulated the expression of miR-200 and let-7 family miRNAs participating in EMT. Transfection of cells with NF-κB p65 subunit and anti-miR-200s attenuated the effect of garcinol on both apoptosis induction and breast cancer cell invasion. The results were confirmed in in vivo xenograft mouse model studies, where garcinol inhibited NF-κB, miRNAs, vimentin, and nuclear β-catenin [76]. Furthermore, garcinol-induced inhibition of acetyltransferase and the effect on cell proliferation, cell cycle progression and apoptosis were investigated in estrogen-stimulated MCF-7 cells. Treatment with garcinol resulted in the inhibition of proliferation, cell cycle progression arrest at the G0/G1 phase, and the increase in apoptosis. These effects were linked to hyperacetylation levels of histones and nonhistone NF-κB/p65 [77].
2.8. Isobavachalcone
Isobavachalcone (IBC) is a naturally occurring prenylated chalcone derived from the seeds of Psoralea corylifolia L. It is known as a phytoestrogen with an antitumor effect. IBC concentration- and time-dependently induced apoptosis of both MCF-7 and MDA-MB-231 cells. It induced inhibition of MCF-7 cell proliferation, triggered apoptosis and autophagy. These effects were mediated by increasing Bax expression and downregulating the expressions of Bcl-2, Akt and p-Akt-473 proteins [78]. In MDA-MB-231 cells, IBC action resulted in multiple cell death processes (apoptosis, necroptosis, autophagy). These were attributed to the downregulation of Akt and p-Akt-473 and an increase in the Bax/Bcl-2 ratio. In addition, IBC induced mitochondrial dysfunction, thereby decreasing cellular ATP levels and increasing ROS accumulation [79]. IBC may also interfere with ERα and influence estradiol-induced paclitaxel resistance. It downregulated ERα. This effect resulted in decreased expression of CD44 (cell-surface glycoprotein involved in cell–cell interactions) and thus inhibited tumor growth of ER-positive breast cancer cells in paclitaxel-resistant xenograft models [80].
2.9. Other Chalcones
Panduratin A is isolated from plants like Boesenbergia pandurata. The apple polyphenols phloretin and pinostrobin are from Uvaria chamae, and some other plants possess many health benefits. Treatment of MCF-7 breast cancer cells with panduratin A resulted in a time- and dose-dependent inhibition of cell growth. The mechanism resulted from increased activity/expression of mitochondrial cytochrome C, caspases 7, 8 and 9 with a significant increase in the Bax/Bcl-2 ratio, suggesting the involvement of a mitochondrial-dependent apoptotic pathway. Several other proteins involved in cell cycle regulation were also affected [81]. Evaluation of pinostrobin resulted in selective inhibition of the migration of both MDA-MB-231 and T47D malignant cells, while the effects on MCF10A cells were blunted. Surprisingly, the inhibitory actions on cell adhesion, cell spreading, and focal adhesion formation of both malignant cell lines were not accompanied with anti-proliferation effects [82]. A distinct mechanism of action inhibiting cell proliferation of MDA-MB-231 TNBC cell line was discovered in phloretin. This apple polyphenol, which is specific antagonist of GLUT2 protein, inhibited MDA-MB-231 cell growth and arrested the cell cycle. It also decreased migration of the MDA-MB-231 cells through the inhibition of paxillin/FAK, Src, and alpha smooth muscle actin (α-sMA) through the activation of E-cadherin. In addition, the anti-tumorigenic effect of phloretin was demonstrated in vivo using BALB/c nude mice bearing MDA-MB-231 tumor xenografts. The authors concluded that the inhibition of GLUT2 phloretin could potentially suppress TNBC tumor cell growth and metastasis [83]. Similarly, a totally different mechanism of cell death in breast cancer cells was ascribed to trans-chalcone. In breast cancer cells, its anti-proliferative effects were considered to be mediated through the increase in heme oxygenase-1 expression. Blocking this enzyme diminished the effect of trans-chalcone on cell growth inhibition [84].
This entry is adapted from the peer-reviewed paper 10.3390/ijms241210354
This entry is offline, you can click here to edit this entry!