Pattern recognition receptors (PRRs) play a central role in the inflammation that ensues following microbial infection by their recognition of molecular patterns present in invading microorganisms but also following tissue damage by recognising molecules released during disease states. Such receptors are expressed in a variety of cells and in various compartments of these cells. PRR binding of molecular patterns results in an intracellular signalling cascade and the eventual activation of transcription factors and the release of cytokines, chemokines, and vasoactive molecules. PRRs and their accessory molecules are subject to tight regulation in these cells so as to not overreact or react in unnecessary circumstances. They are also key to reacting to infection and in stimulating the immune system when needed. Therefore, targeting PRRs offers a potential therapeutic approach for chronic inflammatory disease, infections and as vaccine adjuvants. In this review, the current knowledge on anti-viral PRRs and their signalling pathways is reviewed. Finally, compounds that target PRRs and that have been tested in clinical trials for chronic infections and as adjuvants in vaccine trials are discussed.
1. Introduction
Infection as well as tissue injury/stress can activate inflammatory responses. Inflammation is widely recognised by the cardinal symptoms of fever, redness, oedema, pain, and loss of function. Inflammation is necessary to help with the removal of the source of infection, to help protect the infected tissue(s), and to restore homeostasis. Both immune and non-immune cells produce cytokines, chemokines, and vasoactive peptides. These molecules attract immune cells such as neutrophils that are normally restricted to the vasculature, allowing them to enter into the inflamed or infected tissue [
1,
2,
3]. Viruses cannot survive or replicate themselves and so need a host. They possess several distinct features that have allowed our immune system to develop a number of strategies to detect and remove viruses [
4]. However, viruses have in turn developed strategies of their own to hide and evade our immune system. They can be detected at several stages in their life cycle and our immune system usually initiates an appropriate response [
5]. These responses are usually initiated by a key family of receptors known as pattern recognition receptors (PRRs) that recognise pattern-associated molecular patterns (PAMPs). The binding of PAMPs to PRRs results in the initiation of an appropriate and regulated inflammatory response to the infection [
6]. PRRs can be grouped into RIG-like receptors (RLRs), NOD-like receptors (NLRs), C-type lectin receptors, and Toll-like receptors (TLRs). Stimulating these receptors may help in the fight against infection. This review will discuss the current knowledge specifically on anti-viral PRRs and molecules that target them that have been tested to date in clinical trials.
2. Anti-Viral Pattern Recognition Receptors
Viruses can be categorised as RNA viruses, DNA viruses or retroviruses (which can have RNA or DNA in some cases). They can also be categorised by whether their nucleic acid is positive or negative sense. In addition, replication of these viruses also generates intermediate molecules such as ssRNA and/or dsRNA [
7,
8]. These nucleic acids are distinct from the host and are often contained in a specific compartment following infection separate from host nucleic acid [
9,
10]. Anti-viral pattern recognition receptors employ a variety of strategies to detect nucleic acids from viruses and some bacteria. Detection can occur in the endosomes (usually following phagocytosis by immune cells) or in the cytosol of immune and non-immune cells where they sense DNA or RNA from viruses that have shed their protein coat prior to replication or genome integration [
11].
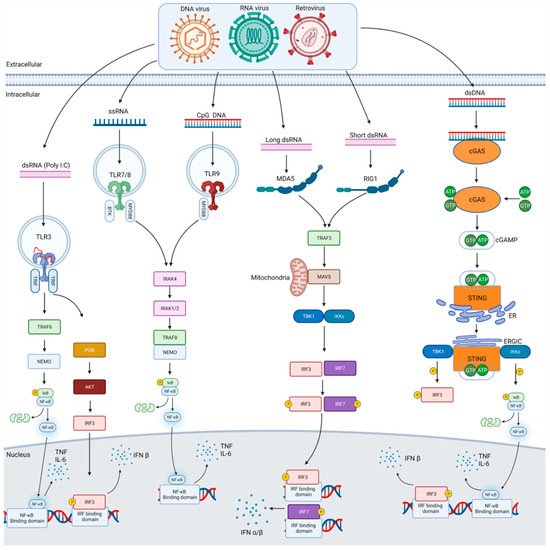
The first PRRs to be discovered were Toll-like receptors. They are most highly expressed in immune cells, but some are also expressed in non-immune cells such as epithelial cells. Basal expression levels vary across the body [
12,
13]. TLRs 1, 2, 4, 5, 6 and 10 are found on the cell surface and generally detect bacterial or fungal PAMPs and some viral proteins, whereas TLRs 3, 7, 8, 9 are located intracellularly and mostly detect viral and bacterial nucleic acids. The cellular localisation of PRRs is important to their function. Intracellular TLRs act within the endosomal compartment, which generally excludes host nucleic acid and allows them to better distinguish foreign RNA/DNA from that of the host, the in [
14].
Other receptor families such as RLRs, NLRs and DNA sensors such as absent in melanoma 2 (AIM2) and cyclic GMP-AMP synthase (cGAS) can recognise a range of microbial nucleic acids in the cytosol [
4]. This strategy allows the innate immune system to detect nucleic acids from viruses at different stages of their life cycle and cellular localisation. PRR activation culminates in the release of type I interferons and cytokines such as interleukin 1β (IL1β), IL6 and tumour necrosis factor α (TNFα) [
11,
15]. Type I interferon release acts in paracrine and autocrine pathways via interferon alpha receptor 1 and 2 (IFANR1/2). This leads to activation of signalling pathways that culminates in the induction and release of interferon stimulated genes (ISGs) such as 2′, 5′-oligoadenylate synthetase (OAS) and protein kinase R (PKR) that help suppress virus replication and assembly in the host cell [
16]. Cytokines alter the vasculature to help activate and recruit immune cells to the site of infection [
17].
In humans, there are 10 TLR family members that are membrane-bound receptors located intracellularly and on the cell surface [
14]. Ligand binding is carried out by the extracellular domain, which consists of leucine-rich repeats (LRRs), while intracellular signalling is carried out by the intracellular Toll-interleukin-1 resistance domain (TIR) [
18]. Ligand recognition initiates a signal transduction cascade within the cytoplasm via myeloid differentiation primary response gene 88 (MyD88), an adaptor molecule, and specific kinases such as IL1 receptor-associated kinases (IRAK) 1/2/4. A series of phosphorylation reactions activates transcription factors such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), or interferon regulatory factors (IRF) 3/5/7. These then move to the nucleus inducing transcription of cytokines and interferons (
Figure 1) [
19].
Figure 1. Anti-viral pattern recognition signalling pathways. Shown in this figure are the various signalling pathways used to respond to viral infection from DNA, RNA and retroviruses. Each of the major forms of nucleic acids that act as PAMPs from such viruses are illustrated along with the PRRs they bind to including TLRs, RLRs and DNA sensors such as cGAS. Each pathway activates specific kinases via their own adaptor molecules, and this ultimately leads to the activation of transcription factors that translocate to the nucleus and help initiate the transcription of cytokines such TNFα and IL6 or type I interferons, e.g., IFNβ, which then get released from the cell.
The intracellular TLR, TLR3 was identified as recognising polyinosinic-polycytidylic acid (poly(I:C)), a synthetic analogue of double stranded RNA (dsRNA) [
20]. TLR3 is important in recognising ssRNA viruses such as respiratory syncytial virus (RSV), encephalomyocarditis virus, West Nile virus and small interfering RNAs [
19]. Its role has been illustrated using
tlr3 deficient mice, which are susceptible to lethal viral infection [
21]. TLR7 recognises ssRNA from viruses such as vesicular stomatitis virus (VSV), influenza type A (IAV) and human immunodeficiency virus (HIV) but was originally described as recognising imidazoquinolinone chemical derivatives such as imiquimod and resiquimod [
19,
22].
Expression of TLR7 is relatively high in plasmacytoid dendritic cells (pDCs). TLR8 also recognises ssRNA and is similar phylogenetically to TLR7. Interestingly, no release of interferon or cytokines occurs in the absence of TLR7 following ssRNA stimulation but immune responses are normal in TLR8 deficient mice [
19,
22]. It may have other roles in immune response. TLR9 has been shown to recognise unmethylated CpG containing ssDNA and it induces the expression and release of Type 1 interferon [
23]. Like TLR7, TLR9 has relatively high constitutive expression in pDCs and B cells. It is important in the prevention of replication by certain viruses such as herpes simplex virus (HSV) and adenovirus [
24,
25]. Further details of TLR signalling have been discussed in detail elsewhere [
26].
TLR recognition of RNA and DNA is limited to the endosomal compartment of immune cells. Other PRRs have evolved for sensing nucleic acids in different cellular locations and cell types to detect viruses at different stages of their life cycle [
27]. The second category of PRR are the cytosolic RNA sensors. These are DExD/H box helicases (DHX) and include retinoic acid inducible gene I (RIG-I), melanoma differentiation-associated antigen 5 (MDA5) and laboratory of genetics and physiology 2 (LGP2) which does not have the N-terminal caspase recruitment (CARD) domain. RIG-I and MDA5 are primarily involved in the cytosolic recognition of RNA viruses whereas LGP2 regulates MDA5 and RIG-I signalling [
28]. RLRs are expressed across many tissues and cell types. RIG-I recognises short RNA molecules (<300 bp) in the cytosol containing 5′-diphosphates as well as 5′-triphosphorylated uncapped RNA. MDA5 recognises long dsRNA (>300 bp) as well as high molecular weight branched RNAs [
7]. Both receptors use mitochondrial anti-viral signalling protein (MAVS) to signal leading to activation of the inhibitor of κB kinase (IKK)-related kinases TANK-binding kinase (TBK1) and IKKi which then activate the transcription factors IRF3 and IRF7 and induce type I interferon transcription (
Figure 1) [
15,
29]. Viruses commonly recognised by RIG-I include (−)ssRNA viruses such as influenza A/B as well as (+)ssRNA viruses, e.g., hepatitis C virus whereas MDA5 commonly recognises dsRNA viruses as well as some (+)ssRNA viruses [
4].
The third category of PRRs are cytosolic receptors that recognise microbial DNA during infection. DNA is mostly contained within the nucleus or mitochondria in mammalian cells. During infection, viruses inject DNA into the cytosol of cells. DNA sensors include absent in melanoma 2 (AIM2) and cyclic GMP-AMP (cGAMP) synthase (cGAS) among others. This then activates an endoplasmic reticulum-bound adaptor called stimulator of interferon genes (STING). STING undergoes a conformational change and inducing the production of type I interferon via the activation of TBK1 and IRF3 (
Figure 1). All of these are widely expressed and recognise DNA from DNA viruses, retroviruses, genomic DNA and cyclic dinucleotides (CDNs) from bacteria and self-DNA from dead cells [
30,
31]. Recognition of cytosolic DNA by AIM2 activates the inflammasome leading to activation of caspase 1 which subsequently cleaves gasdermin and leads to the release of interleukin 1 and 18 [
32,
33]. Upon recognition of DNA of various sizes by cGAS it can synthesise cGAMP from ATP and GTP ([
34,
35,
36].
This entry is adapted from the peer-reviewed paper 10.3390/cells10092258